利用野栽杂交分离群体定位水稻结实率QTLs
2017-08-11潘英华温国泉徐志健梁云涛
潘英华,温国泉,徐志健,梁云涛*
(1.广西壮族自治区农业科学院水稻研究所,广西 南宁 530007;2.广西水稻遗传育种重点实验室,广西 南宁 530007;3.广西壮族自治区农业科学院农业科技信息研究所,广西 南宁 530007)
利用野栽杂交分离群体定位水稻结实率QTLs
潘英华1,2,温国泉3,徐志健1,梁云涛1*
(1.广西壮族自治区农业科学院水稻研究所,广西 南宁 530007;2.广西水稻遗传育种重点实验室,广西 南宁 530007;3.广西壮族自治区农业科学院农业科技信息研究所,广西 南宁 530007)
【目的】利用野栽杂交分离群体定位水稻结实率,为能更好地挖掘和利用野生稻中控制穗结实率基因的QTL位点提供参考。【方法】分别以广西普通野生稻资源Y03为父本和栽培稻品种日本晴为母本,经过杂交构建包含142个单株的F2定位群体,然后利用覆盖水稻基因组的184对SSR分子标记,采用复合区间作图法(CIM),以LOD=2.5为阈值检测控制结实率的QTL。【结果】共检测到3个影响结实率的QTL。其中,2个QTL位于第1染色体,1个QTL位于4号染色体上,并分别命名为qSSR1-1,qSSR1-2和qSSR4-1。qSSR1-1位于第1染色体RM486~RM5501,表型贡献率为14.49 %;qSSR1-2位于第1染色体RM102~RM315,表型贡献率为8.63 %;qSSR4-1位于第4染色体RM252~RM119,表型贡献率为8.27 %。对结果进行分析还发现,在3个QTL位点上来源于野生稻亲本Y03的等位基因均有利于提高水稻结实率。随后,根据获得的主效QTL定位信息最终开发出与水稻结实率性状紧密连锁、可用于分子育种的分子标记RM119。【结论】发掘的新QTL和性状连锁标记可为水稻产量性状QTL的发掘和分子标记辅助选择育种提供重要的基因资源和分子选择工具。
普通野生稻;杂交;控制穗结实率;QTL
【研究意义】水稻是我国主要的粮食作物[1],其产量直接影响我国粮食安全。而水稻植株的稻谷产量主要是受粒重、穗粒数、有效穗数、穗结实率等性状所影响。其中,稻穗的结实率对产量的贡献最为重要。但由于结实率的遗传特性较为复杂,是由多基因控制,并受环境影响,作为栽培稻的野生近缘种,野生稻资源中蕴含有丰富的基因/QTL位点,野生稻成为开展重要农艺性状分子遗传研究和基因发掘的优良供体。因此,开展野生稻资源中定位控制结实率QTL位点的研究对水稻穗结实率性状的遗传机制和水稻分子育种研究均具有重要意义。【前人研究进展】目前,有关学者在水稻的不同染色体上定位到多个控制结实率的QTL位点,如Elsheikh等[2]用粳稻日本晴与籼稻N040212组配149份BC2F6家系定位到了3个与结实率相关的QTL,分别位于第1,7和9号染色体上,并用分离群体将结实率QTLqSS-1定位在1号染色体93.5 kb区间内。Zhou[3]等利用来源于Guanghui 116和 LaGrue 的307个重组自交系构建遗传连锁图谱,定位结实率,单株产量的QTL。戴陆园等[4]研究表明,可能有1~2对主效基因参与调控水稻结实率。韩龙植等[5]利用 SSR 标记对水稻结实率数量性状位点进行检测。共检测到与结实率相关的QTL14个,其中qSSR4位于4号染色体上,定位于RM335~RM307。而栽培稻与野生稻杂交后代,由于亲本的遗传背景差异较大,结实率也呈现出分离较大的现象。袁平荣等[6]用云南元江普通野生稻、景洪普通野生稻,和东乡野生稻分别与籼稻和粳稻进行杂交,6种杂交组合的后代结实率为47.48 %~71.67 %。孙希平等[7]通过花粉管通道法将普通野生稻的DNA导入宁夏水稻品种宁粳16号和宁粳23号中,获得D4代材料,发现导入系的每穗实粒数变异丰富,出现导入系每穗实粒数比受体低或比受体高。韦贵剑等[8]利用广西普通野生稻与籼稻9311杂交杂交并自交获得F3群体,发现F3家系中秕谷数变异系数最大,结实率的变异系数为35.45 %。截止至2017年4月,Gramene已经公布了330个水稻结实率QTL(http://archive.gramene.org)。【本研究切入点】目前野生稻的重要基因尚未得到深入挖掘,还需从更多不同的遗传材料中发掘新的QTL位点才能更深入和全面地阐释其内在的遗传规律,更好地为育种实践服务。【拟解决的关键问题】以广西普通野生稻为父本,栽培稻品种日本晴作为母本,构建定位群体,使用分子标记技术进行穗结实率QTL检测和遗传效应分析,并开发出与结实率性状紧密连锁的分子标记。以期为揭示穗结实率性状的遗传机制和水稻分子育种奠定理论和材料基础。
1 材料与方法
1.1 作图群体的构建
2014年晚造,以栽培稻品种日本晴为母本,与编号为Y03的广西普通野生稻材料配组杂交获得F1。然后,2015年早造,种植F1,籽粒成熟后收获自交种子,构建F2作图群体用于目标性状QTL检测分析。
1.2 田间试验和考种
2015年晚造,F2定位群体共142个单株种植于广西农业科学院水稻研究所试验田。同时种植亲本材料日本晴以及37份水稻核心种质材料。单本栽插,株行距为20 cm×23 cm,按常规管理方法进行田间管理。籽粒成熟后,分别采收所有水稻材料的单株主穗,分别统计每个单株主穗的实粒和秕粒数,然后计算出穗结实率。
1.3 DNA提取和分子标记筛选
DNA提取方法在参考Murray[9]等方法的基础上进行优化改进。参照McCouch[10]等公布的SSR分子标记信息,从中挑选出分布于水稻12条染色体上的600个SSR标记。所有分子标记引物均由北京华大生物技术服务有限公司合成。
然后,将600个SSR标记在亲本材料Y03和日本晴间进行多态性检测,最终筛选出多态性好、条带清晰的184个分子标记用于下一步QTL检测分析。所有标记均匀分布于水稻12条染色体上。PCR扩增程序、凝胶电泳和银染均参照Panaud等[11]的方法。
1.4 数据分析和QTL检测
表型数据采用分析软件SAS8.0进行统计分析。使用软件Mapmaker Exp 3.0进行数据分析,构建了包含184个SSR标记的分子标记连锁图谱,共分为12个连锁群。然后,应用 Mapmaker QTL 软件进行QTL分析,采用复合区间作图法(Composite Interval Mapping,CIM)进行全基因组扫描检测结实率相关QTL,以LOD=2.5 为阈值判断 QTL是否存在。参照McCouch等[12]的原则命名QTL。
1.5qSSR4-1位点的表型效应分析
利用qSSR4-1基因组区段两端的连锁标记RM252和RM119检测F2分离群体。如果两个标记扩增带型均与亲本日本晴一致,可推测植株在qSSR4-1位点上携带日本晴等位基因;如两标记扩增带型与Y03相同则表明该位点为Y03等位基因。根据每一单株的标记检测结果,将携带日本晴和Y03等位基因的植株分别划分为A、B2个组。分别统计各组的平均结实率,并计算两者间的差异显著性。然后依据分析结果判断qSSR4-1对水稻结实率的作用大小。
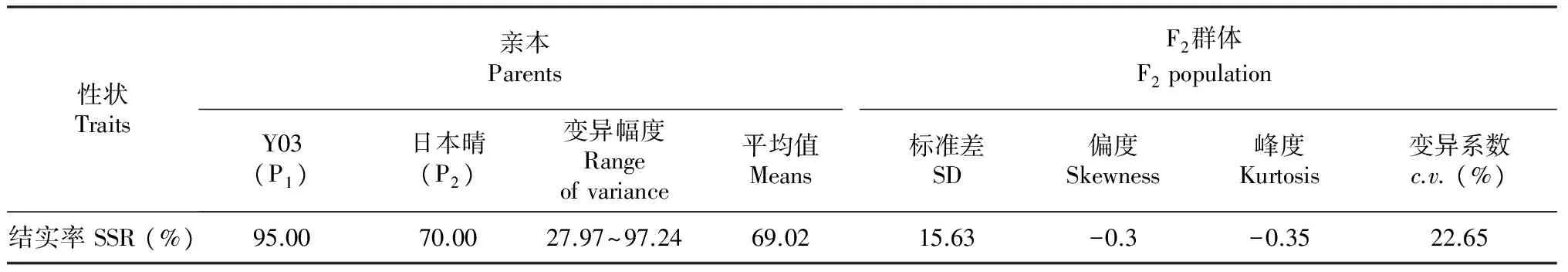
表1 亲本及其F2 群体结实率表现
1.6 连锁标记的有效性验证
通过对结实率不同的36份水稻核心种质材料进行分子鉴定,从而验证连锁标记RM119检测结实率的准确性。并将RM119标记的Y03等位基因型记为高结实率基因型G,日本晴的等位基因型记为低结实率基因型D。以结实率>95.00 %为高结实率表型,结实率<60.00 %为低结实率表型。通过综合分析基因型和表型数据,最终确定连锁标记对结实率高低检测的有效性。
2 结果与分析
2.1 亲本日本晴与F2群体的结实率表现
从表1可以看出,亲本日本晴与普通野生稻Y03的结实率分别为70.00 %和95.00 %,亲本结实率之间差异达到极显著水平(P<0.01);F2群体中单株的结实率呈现正态分布,分布范围27.97 %~97.24 %,变异系数为22.65 %;群体出现连续变异,出现明显的双向超亲分离,且存在个别超亲高结实率现象(表1,图1)。结实率性状在F2群体中表现为正态连续分布,说明该性状受多个基因控制。
2.2 结实率相关QTL检测分析
共检测到3个与结实率相关的QTL,其中2个QTL位于1号染色体上,1个QTL位于4号染色体上,分别为qSSR1-1,qSSR1-2,qSSR4-1。qSSR1-1的LOD值为3.72,对表型的贡献率为14.49 %,位于第1染色体RM486~RM5501,提高结实率的增效等位基因来源于广西普通野生稻Y03;qSSR1-2的LOD值为2.70,对表型的贡献率为8.63 %,位于第1染色体RM102~RM315,增效等位基因来源于Y03;qSSR4-1位于第4染色体RM252~RM119,LOD值为2.55,对表型的贡献率为8.27 %,增效等位基因同样也来源于Y03(表2,图2)。
2.3qSSR4-1的表型效应分析
将F2分离群体中携带qSSR4-1不同等位基因区段的单株分为A、B2个组,分析两个组的平均结实率差异,从而尝试了解QTL位点qSSR4-1对目标性状表型的影响作用。结果显示,携带qSSR4-1日本晴等位基因型的A组平均结实率为68.06 %,携带Y03等位基因型的B组平均结实率为76.59 %,B组比A组的结实率高8.53 %,但两者间差异未达到显著。
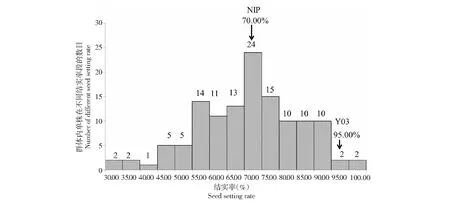
图1 结实率频度分布Fig.1 The frequency distribution of seed setting rate

性状Trains基因座Locus染色体Chr.标记区间Markerinterval阈值LOD效应值EffectE1E2贡献率(%)PVE等位基因来源Sourceofallele结实率qSSR1⁃11RM486~RM55013.720.08-0.0214.49Y03结实率qSSR1⁃21RM102~RM3152.70.07-0.018.63Y03结实率qSSR4⁃14RM252~RM1192.550.05-0.068.26Y03
注:E1:加性效应;E2:显性效应。
Note:E1:Additive gene effect; E2:Dominance gene effect.
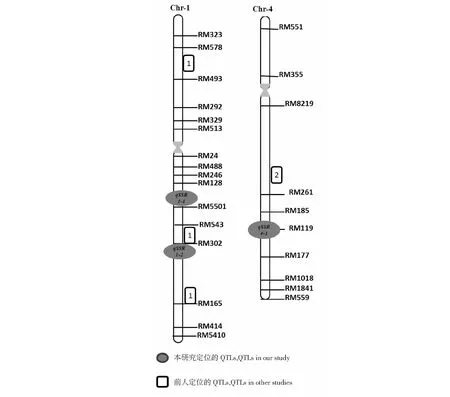
图2 结实率QTL 的染色体定位情况Fig.2 Chromosomal location of QTLs on seed setting rate
2.4 连锁标记RM119的有效性验证
利用结实率主效QTL的连锁标记RM119检测具有不同结实率的36份水稻核心种质,分析发现,16份高结实率(结实率>95.00 %)水稻核心种质中具有G基因型的材料有13份,占高结实率种质的81.25 %;20份低结实率(结实率<60.00 %)水稻核心种质中有15份具有D基因型,占全部低结实率种质的75.00 %(图3)。表明,连锁标记RM119可以针对结实率对多个不同的水稻材料进行有效鉴定筛选。
3 讨 论
3.1 栽野杂交后代结实率分离特点
普通野生稻与栽培稻遗传背景差异较大,通过两者杂交,可将不同来源的遗传因子进行重组,从而产生出许多表型差异较大的个体,整个群体上看则表现出性状明显分离。如云南元江普通野生稻、景洪普通野生稻,和东乡野生稻分别与籼稻和粳稻的杂交后代结实率个体之间差异极大[6]。
在本研究中,使用普通野生稻与栽培稻品种日本晴进行杂交产生F2群体,群体中的结实率分布为27.97 %~97.24 %,呈现正态分布,变异系数为22.65 %,出现连续变异,存在明显的双向超亲分离,且存在个别超亲高结实率现象。可以看出,本研究的F2群体结实率表现与以往研究[6-8]基本一致。群体内结实率的差异极大,出现极端个体。本研究采用的广西普通野生稻作为供体,与前人研究所用的材料不同。而且,本研究还发现F2群体在正常自然环境下的平均结实率并不高,这可能是与双亲背景差异大有着密切关系。由于亲缘较远的野生稻和栽培稻进行杂交容易产生出表型性状差异较大的个体,从而可以利用杂交后代群体中遗传物质大量重组以及性状明显分离等特性开展基因/QTL定位研究,有助于高效挖掘目标性状突出的优异基因资源。
3.2 与前人精细定位或克隆结实率相关性状的 QTL 比较
水稻结实率直接影响水稻产量潜力,为多个基因控制的数量性状,易受环境条件的影响[13-17],周勇等[18]利用广陆矮4号与日本晴的染色体单片段代换系定位水稻结实率QTLs,检测到9个与结实率相关的QTLs,分布在7条染色体上,能在多个环境下检测到。陈庆全等[19]以结实率较低的籼稻品种T219和结实率较高的籼稻品种T226为亲本构建了202个重组自交系,利用8种不同环境条件的实验结果,定位了分布在9条染色体上的17个与结实率
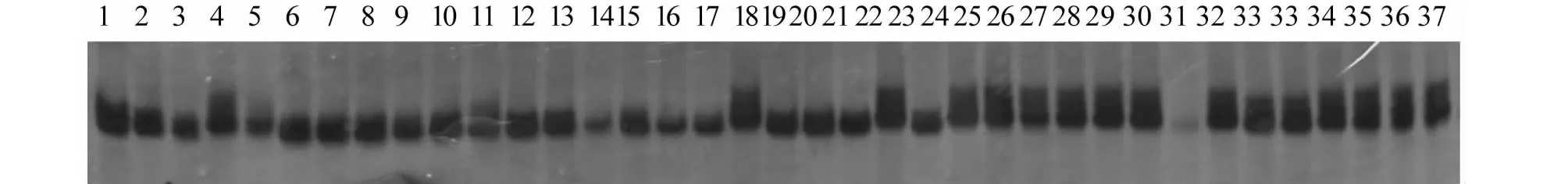
1~20结实率小于60 %,21为日本晴,22-37结实率大于95 %Lane 1-20 are rice varieties of which seed setting rate is less than 60 %;Lane 21 is Nipponbare, Lane 22-37 are rice varieties of which seed setting rate is more than 95 %图3 RM119标记对36份水稻种质的扩增结果Fig.3 Amplification result of molecular marker RM119 in 37 rice germplasm
相关的QTL,其中qSSR4-1定位在4号染色体RM3216C~RM401之间,贡献率9.9 %。到目前为止,已克隆的与水稻结实率相关的只有PTB1基因,Li等[20]利用低结实率水稻突变体ptb1和高结实率野生型材料Gui630组配了F2群体,把与结实率相关的QTL定位在5号染色体72 kb区间内,经过测序,发现基因LOC_Os05g05280 引起了232nt mRNA的缺失,导致了转录的提前终止。PTB1基因编码一个环型E3泛素连接酶,通过促进花粉管的伸长调控水稻结实率。本研究共检测到3个影响水稻结实率的QTL,其中2个QTL位于1号染色体,另1个位于4号染色体,分别命名为qSSR1-1,qSSR1-2和qSSR4-1。qSSR1-1位于第1号染色体RM486~RM5501;qSSR1-2位于第1号染色体RM102~RM315;qSSR4-1位于第4号染色体RM252~RM119。与已公布的已克隆的控制结实率基因,普通野生稻定位的与结实率相关的QTL比较,本研究检测到的QTL染色体区段与已往定位结果并不相同,因此可以推测为新的控制水稻结实率QTL。而且上述3个QTL位点上来源于广西普通野生稻Y03的等位基因均有利于提高水稻结实率,是今后开展基础理论研究和育种利用宝贵的基因材料。
3.3 结实率相关连锁标记在育种上的利用
4 结 论
本研究检测到3个来源于普通野生稻的控制结实率的QTL。其中,在第4染色体RM252~RM119标记区间检测到的qSSR4-1与之前报道的精细定位的结实率QTL均不重叠,是一个新的控制结实率QTL。另外,还开发出1个与结实率性状紧密连锁的分子标记RM119,经验证表明,可以对多个水稻育种亲本进行有效的分子标记辅助选择。本研究发掘的新QTL和性状连锁标记可为水稻产量性状QTL的发掘和分子标记辅助选择育种提供提供重要的基因资源和分子选择工具。
[1]Manickavelu A, Nadarajan N,Ganesh S K,et al. Drought tolerance in rice:morphological and molecular genetic consideration[J].Plant Growth Regulation,2006,50(2-3):121-138.
[2]Elsheikh Y M A,Zhang Y P,Yu J P et al. Mapping of three QTLs for seed setting and analysis on the candidate gene forqSS-1 in rice (OryzasativaL.) [J]. Journal of Integrative Agriculture,2016,15(4):735-743.
[3]Zhou S X,Zhu M S,Wang F,et al. Mapping of QTLs for Yield and its components in a rice recombinant inbred line population [J]. Pakistan Journal of Botany,2013,45(1):83-189.
[4]Dai L Y,Ye C R,Xu F R,et al. Genetic analysis on cold tolerance characteristics of Yunnan rice landrace (OryzasativaL.)Kunmingxiaobaigu[J]. Chinese Journal of Rice Science,1999,13(2):73-76.
[5]韩龙植,张三元,乔永利,等. 不同生长环境下水稻结实率数量性状位点的检测[J].作物学报,2006,3(7):1024-1030.
[6]袁平荣,杨从党,周 能,等. 云南元江普通野生稻分化的研究——普通野生稻与籼粳亲和性的初步研究[J]. 农业考古,1998(1):38-40.
[7]孙希平,柳哲胜,李树华,等. 普通野生稻 (Oryzarufipogon) D N A导入栽培稻后代主要性状的遗传变异[J].植物遗传资源学报, 2009,10(2):267-271.
[8]韦贵剑,黄大辉,梁仁敏,等. 野生稻与籼稻杂交后代田间表现及优良单株选育[J]. 南方农业学报, 2015,46(5):739-744.
[9]Murray M G,Thompson W F. Rapid isolation of high molecular weight plant DNA [J]. Nucleic Acids Research,1980,8(19):4321-4326.
[10]McCouch S R,Kochert G,Yu Z H,et al. Molecular mapping of rice chromosomes [J]. Theoretical and Applied Genetics,1988,76:815-829.
[11]Panaud O,Chen X,McCouch S R. Development of microsatellite markers and characterization of simple sequence length polymorphism (SSLP) in rice (OryzasativaL.) [J].Molecular and General Genetics,1996,252(5):597-607.
[12]McCouch S R. Gene nomenclatre system for rice[J]. Rice,2008,1(1):72-84.
[13]Sawada S. Studies of sterile-type cool injury in rice plants with special reference to the mechanism and inheritance of sterility[J]. Research Bulletin of Obihiro Zootechnical University,1978,1(10):837-883.
[14]Khan D R,Mackill D J,Vergara B S. Selection for tolerance to low temperature induced spikelet sterility at anthesis in rice[J]. Crop Science,1986,26(4):694-698.
[15]Kaw R N,Moon H P,Yae J D,et al. Estimates of combining ability for cold tolerance at reproductive stage in rice[J]. Korean Journal of Breeding Science,1989,21(3): 188-195.
[16]Han L Z,Yuan D L,Xuan Y S,et al. Genetic analysis of cold water response on several agronomic traits of rice [J]. Chinese Journal of Rice Science,2004,18(1): 23-38.
[17]Kaw R N,Moon H P,Yae J D,et al. Estimates of combining ability for cold tolerance at reproductive stage in rice[J]. Korean Journal of Breeding Science,1989,21(3): 188-195.
[18]周 勇,缪 军,陶亚军,等. 利用染色体单片段代换系定位水稻结实率QTLs[J]. 扬州大学学报,2013,34(4):45-50.
[19]陈庆全,余四斌,李春海,等. 不同环境条件下水稻结实率的 QTL 定位分析[J]. 农业生物技术学报,2007,15(5):834-840.
[20]Li S c,Li W B,Huang B, et al. Natural variation inPTB1 regulates rice seed setting rate by controlling pollen tube growth[J]. Nature communications,2013,doi:10.1038/ncomms3793.
[21]刘 驰,马增凤,秦 钢,等,利用分子标记辅助选择培育水稻广亲和保持系[J].南方农业学报,2015,46(3):365-369.
(责任编辑 汪羽宁)
Mapping of QTLs for Seed Setting Rate Using HybridPopulation ofOryzarufipogonGriff. × Cultivated Rice
PAN Ying-hua1,2, WEN Guo-quan3, XU Zhi-jian1, LIANG Yun-tao1*
(1.Rice Research Institute of Guangxi Academy of Agricultural Sciences, Guangxi Nanning 530007,China; 2.Guangxi Key Laboratory of Rice Genetics and Breeding, Guangxi Nanning 530007, China; 3.Agricultural Information Institute of Guangxi Academy of Agricultural Sciences, Guangxi Nanning 530007,China)
【Objective】The seed setting rate of rice was determined by wild-seeded hybrid isolates, to provide a reference for using the genes/QTLs of control seed setting rate in wild rice.【Method】The common wild rice resources in Guangxi Y03 as the male and Nipponbare as the female parent, F2locating population of 142 individuals were constructed by hybridization.184 polymorphic SSR markers were used for mapping QTLs for seed setting rate by combining MAPMAKER EXP 3.0 and composite interval mapping with LOD=2.5. 【Result】Three QTLs associated with seed setting rate were detected. These QTLs distributed on the chromosome 1,4. Two QTLs , which were namedqSSR1-1 andqSSR1-2 respectively, were mapped on the chromosome 1. One QTL,qSSR4-1, was mapped on chromosome 4.qSSR1-1 was mapped between RM486 to RM5501 on chromosome 1, explaining 14.49 % of observed phenotypic variation.qSSR1-2 located between RM102 to RM315 on chromosome 1 and explained 8.63 % of observed phenotypic variation.qSSR4-1 was mapped between RM252 to RM119 on chromosome 4, explaining 8.27 % of the observed phenotypic variation. The alleles for increasing seed setting rate were derived from Y03 atqSSR1-1,qSSR1-2 andqSSR4-1. Moreover, SSR marker RM119, was identified for associating with seed setting rate closely and being able to apply in rice molecular breeding.【Conclusion】Based on the result in the study, some important genes and molecular tool would able to be provided for rice molecular breeding in the future.
OryzarufipogonGriff.;Hybridization;Control seed setting rate;QTL
1001-4829(2017)7-1473-06
10.16213/j.cnki.scjas.2017.7.001
2017-02-22
广西壮族自治区主席科技资金项目“野生稻优异基因的挖掘和利用研究”(1517-03);广西农业科学院基本科研业务专项项目“野生稻资源保护与利用研究”(2015YT14);国家科技基础条件平台项目“国家野生稻种质资源平台(南宁)”(NICGR2016-039);广西农业科学院基本科研业务专项“广西药用野生稻基因MKK3的功能验证”(桂农科2017YZ08);广西农业科学院基本科研业务专项“广西野生稻等保护点目标物种居群多样性分析”(2015JZ12403)
潘英华(1981-),女,广西田东人,博士研究生,助理研究员,主要从事野生稻优异基因挖掘研究工作,E-mail:panyinghua2008@163.com,*为通讯作者:梁云涛,E-mail:liangyt@sina.com。
S511
A
