伞形科植物芫荽果实发育过程中的解剖结构变化
2014-04-09卞桂兰刘启新宋春凤
卞桂兰, 刘启新, 宋春凤
〔江苏省·中国科学院植物研究所(南京中山植物园), 江苏 南京 210014〕
芫荽(CoriandrumsativumLinn.)隶属于伞形科(Apiaceae)芹亚科(Apioideae Drude)芫荽属(CoriandrumLinn.),原产地中海沿岸,西汉时由张骞带入中国,现广泛栽培于全国各地[1]。芫荽俗称香菜,是人类较早食用的芳香类蔬菜[2],还作为祛风剂和抗菌素等大量应用于传统医药[3];另外,芫荽果实内含精油,具有抗氧化、杀菌和清除自由基等多种用途,因而受到人们的广泛关注[4-5]。有关芫荽的研究报道主要集中于种子萌发及栽培技术[6-9]、次生代谢成分及其药用功效[10-14]、食品添加剂及香料[2,15-17]、不同部位解剖结构和分子生物学等方面[8,18-20],而有关其果实发育过程中果实解剖结构方面的研究报道尚不多见。
双悬果是伞形科植物的重要特征之一,而且因其独特而多样的形态和结构常被作为伞形科种类的重要分类依据[21-23]。伞形科中大部分种类的果实为典型的双悬果类型,即果实成熟后2个分生果(即分果)从合生面处完全分离,而且这2个分生果常常分别悬挂在分裂的心皮柄顶端,或虽不悬挂却呈明显靠合状。但芫荽的果实在伞形科中比较特殊,其果实成熟后基本不分离,而且从外部形态上也看不到2个分生果分裂的痕迹。为什么成熟后芫荽果实不像其他种类的双悬果一样分离?它的发育过程和发育机制如何?果实内部结构有什么特殊之处?在伞形科的果实发育类型以及系统分类中的地位如何?等等,对这些问题的答案尚不清楚。由于目前研究者仅对芫荽成熟果实的解剖结构进行了初步研究[8,24],因此要弄清上述疑问,有必要对芫荽果实不同发育时期的解剖结构进行比较观察。
鉴于此,作者采用常规石蜡切片法对芫荽花后不同发育阶段的果实横切面解剖结构进行观察,以期全面了解芫荽果实发育过程中相关解剖结构的变化规律,为该种的深入研究以及为伞形科不同果实类型的发育研究提供解剖学资料。
1 材料和方法
1.1 材料
供试芫荽种植于南京中山植物园伞形科植物资源圃,于2013年4月中旬至5月下旬用随机取样方式采集供试样品。从花瓣完全脱落时开始取样,每隔7 d取样1次,至果实完全成熟为止,期间不间断连续取样。每次取样时随机选定5株不同的样株,从植株上各选取5个果实;样品采后立即用FAA固定,并带回实验室保存,以供制作石蜡切片。
1.2 方法
参照文献[25]、采用常规石蜡切片法制作果实中部横切面的组织连续切片,组织切片厚度8~10 μm,并采用番红-固绿双重染色、中性树胶封片;用Olympus BH2型光学显微镜(日本Olympus公司)对组织切片进行观察、记录、描述和拍照。
2 观察结果
2.1 果实的发育分期及其基本特点
为便于分析和讨论,根据果实外部形态特征,大体可将芫荽果实发育过程分为幼果和果熟2个发育阶段,其中花瓣完全脱落至果实接近圆球形且其表面变为深绿色之前的这一阶段为幼果发育阶段,之后则为果熟发育阶段。每个阶段又根据果实大小、形状和表面色泽等特征分别分为3个时期,各阶段及各时期的具体划分标准如下:
1)幼果发育阶段:即果实逐渐膨大期。从花瓣完全脱落至果实变为近椭圆形且表面呈黄绿色,这一时期为幼果前期(图1-1);果实逐渐膨大至椭圆形且表面呈浅绿色,这一时期为幼果中期(图1-2);果实进一步膨大至接近圆球形且表面呈深绿色,这一时期为幼果后期(图1-3)。
2)果熟发育阶段:即果后期或果实成熟期。果实表面光滑且呈微黄色的圆球形,这一时期为熟果前期(图1-4);果实表面有纵向微突起的果棱且果实表面枯黄,这一时期为熟果中期(图1-5);果实表面纵向突起的果棱更明显且数量较多,这一时期为熟果后期(图1-6)。
2.2 幼果发育阶段的解剖结构特征
2.2.1 幼果发育阶段解剖结构特征总体变化 芫荽幼果发育阶段各时期果实横切面解剖结构见图2。
观察结果显示:在整个幼果发育阶段,果实横切面形状由近椭圆形变成近圆形,其内部已形成2个分生果;2个分生果最初通过合生面紧密相连,随着合生面细胞破损程度逐渐加大,果体中央部分的大部分合生面分离。随着果体增大,果壁厚度逐渐增加,但果壁厚度占果宽的比例却逐渐减小,其中,背部果壁厚度占果宽的比例由原来的近1/3(约27%)逐渐减至近1/5(约20%)。在中果壁中具有5个因果壁油管消失转变形成的大空腔,而且空腔形状随胚乳膨大呈现出由凸透镜状逐渐变扁平的趋势。由于细胞数量和体积不断增加,胚乳不断膨胀扩张;但因胚乳腹面的部分细胞解体,至幼果后期胚乳形状由元宝形变为船形。在整个幼果期,果壁维管束的导管数量几乎没有变化,但是导管管腔有所增大,到幼果后期果壁维管束处微微向上隆起,这也标志着主棱开始发育。此外,在幼果发育阶段,在中果壁、合生面和胚乳细胞中都可观察到大量结晶。
2.2.2 幼果前期解剖结构特征 幼果前期芫荽果实横切面全形及不同部位的解剖结构见图2-1a~1e。这一时期果实横切面近椭圆形;分生果横切面长度约为1 630 μm、宽度约为860 μm,其他特征如下:
果壁较薄;其中背部果壁厚度约为240 μm,约占分生果宽度的27%,且大于合生面果壁厚度。外果壁由1层排列紧密的长方形细胞构成。中果壁由12~14层细胞(侧弯处更多)构成,可明显分为4个层次:第1层次具3~4层薄壁细胞,排列较规则,位于油管外侧,且最外层细胞较大、细胞壁较厚,与外果壁细胞相似,其下方的2~3层细胞稍小;第2层次有6~8层木化细胞,排列紧密、细胞小而密集,紧贴空腔且位于其下方并沿空腔分布呈折线状,而在空腔下方中部的细胞层数多于油管间,呈眉状弧形;第3层次为1~2层压扁的狭长形薄壁细胞;第4层次为3~4层排列较规整、较大的薄壁细胞。另外,空腔之间的中果壁细胞与紧邻的第1层细胞较为相似,但在形状上彼此有所差异。内果壁较薄,仅由1层极度压扁的薄壁细胞组成。
在花期,芫荽子房中可观察到明显的果壁油管,每心皮5个,较大且彼此相接,并环绕果壁1周(另文发表)。但到幼果前期,这类果壁油管几乎消失,并在相应位置上形成大空腔,这些空腔呈凸透镜状,向外微拱起(果实外表面呈棱状),但部分空腔内陷;由于空腔外侧的果壁较薄且紧密相连,内陷空腔的外侧果壁也顺势凹陷,这也是导致果实外表面下凹的原因。合生面油管2个,呈椭圆形。
维管束小,散布于果壁油管下方的中果壁第2层次的木化细胞层中。每个维管束包含导管3~4个,导管直径大于木化细胞。
种皮与胚乳结合紧密,细胞1层,其形状与内果壁和胚乳细胞有明显区别,多为类长方形或类方形,明显小于胚乳细胞。胚乳较大,约占果实横切面面积的50%,背侧外凸、内侧微平,呈近元宝形;胚乳腹面中部的外侧贴生有1个半椭圆形突起的种脊维管束;其中央部位有1个大空腔,即胚着生位置。
2个分生果之间的合生面较厚且与果体长度相等,此时已开始出现离层,而且离层始于心皮柄维管束与中果壁第2层次之间的部位。此外,在中果壁、合生面以及靠近种脊维管束周围的胚乳细胞中分布有大量结晶。
2.2.3 幼果中期解剖结构特征 幼果中期芫荽果实横切面全形及不同部位的解剖结构见图2-2a~2e。与幼果前期相比,这一时期正常发育的分生果有4个明显变化:1)分生果横切面长度增加至2 250 μm、宽度增加至1 160 μm。2)果壁厚度约为250 μm,约占分生果宽度的22%。3)果壁中的部分空腔出现压扁现象,且外突程度明显变小。4)中果壁的木化细胞层变化较大,一是可见圆形和狭长纤维状2类细胞,且二者的走向、细胞层宽度均不同;二是在分生果中该层细胞呈线形波状分布,常成六角状(突起处即为次棱);三是该层细胞延伸至合生面并逐渐变宽,其顶端与合生面维管束相连,总体呈喇叭状。
2.2.4 幼果后期解剖结构特征 幼果后期芫荽果实横切面全形及不同部位的解剖结构见图2-3a~3e。与幼果中期相比,这一时期正常发育的分生果有7个明显变化:1)果实横切面近圆形;分生果横切面长度增加至3 780 μm、宽度增加至1 740 μm。2)果壁厚度约为360 μm,约占分生果宽度的20%。3)合生面细胞破损较多,除两端的外果壁和中果壁外侧部位外,合生面几乎完全分离。4)中果壁细胞普遍增大;第2层次的木化细胞层仍呈波状分布,但其弧度有所减小;在有维管束分布的部位木化细胞层向外微隆起,并与原有的波状突起共同形成隆起幅度较小的类似于主棱状或次棱状的结构。5)胚乳外围细胞开始破损或解体,且腹面内侧废退或解体的细胞更多,胚乳因此而内凹;胚乳开始与种皮脱离,整个胚乳呈船形或元宝形。6)由于受到果体向外扩张的挤压,分生果的5个果壁空腔和合生面油管全部被压扁,此时果实外表面的棱状隆起不明显。7)胚乳腹面细胞内分布有较多的糊粉粒结晶,而背部细胞中结晶较少,且均为染成红色的颗粒。

1. 幼果前期 The early stage of young fruit; 2. 幼果中期 The middle stage of young fruit; 3. 幼果后期 The late stage of young fruit; 4. 熟果前期 The early stage of mature fruit; 5. 熟果中期 The middle stage of mature fruit; 6. 熟果后期 The late stage of mature fruit.
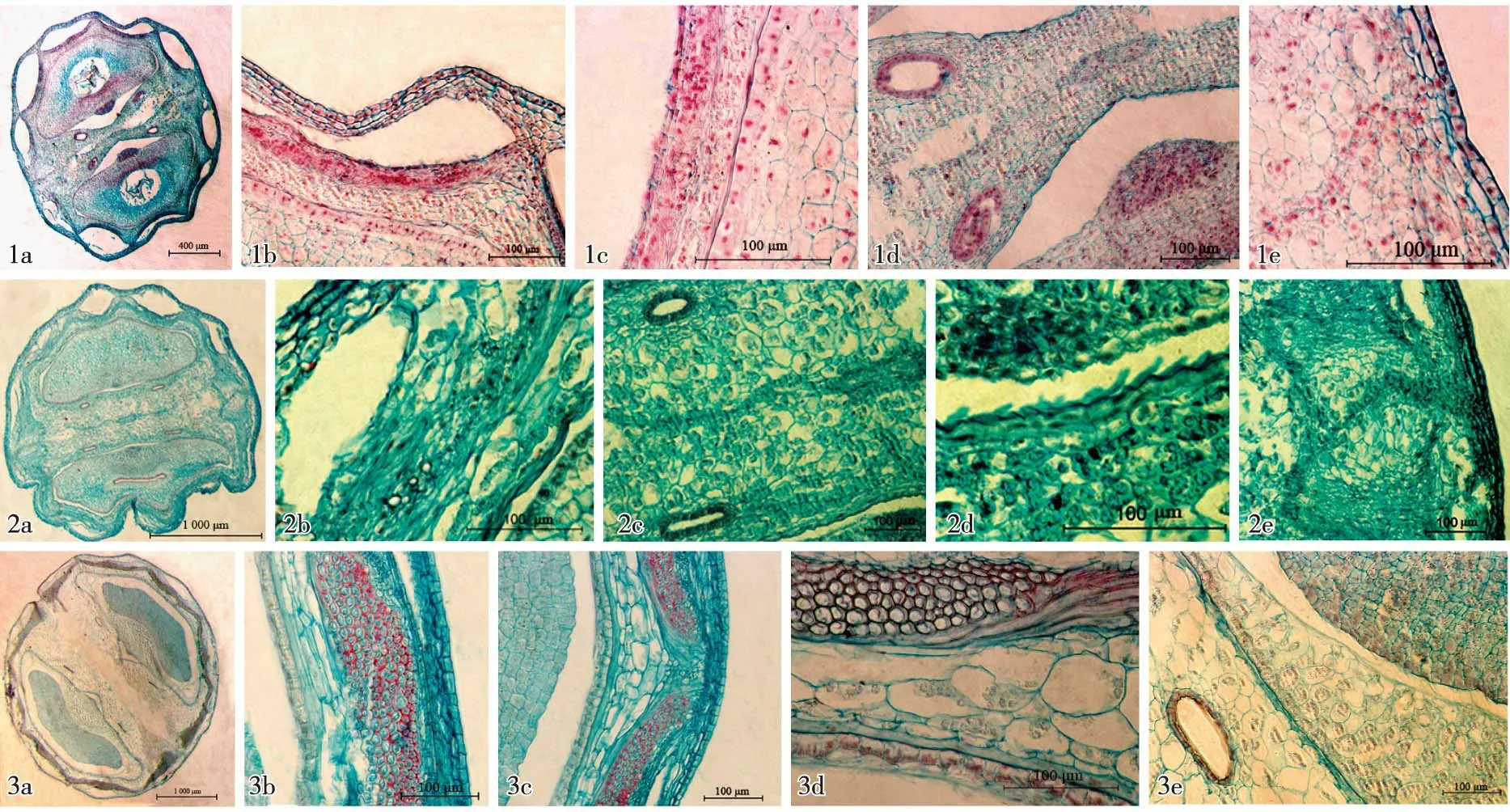
1a-1e. 幼果前期果实横切面解剖结构Anatomical structure of fruit transection at the early stage of young fruit: 1a. 示果实全形 Showing the whole fruit; 1b. 示背部 Showing the dorsal part; 1c. 示空腔下方部位 Showing the bottom part of the cavity; 1d. 示合生面中部 Showing the middle part of commissure; 1e. 示合生面端部 Showing the end part of commissure. 2a-2e. 幼果中期果实横切面解剖结构Anatomical structure of fruit transection at the middle stage of young fruit: 2a. 示果实全形 Showing the whole fruit; 2b. 示空腔部位 Showing the cavity part; 2c. 示合生面中部 Showing the middle part of commissure; 2d. 示腹面内果壁 Showing the endocarp in ventral part; 2e. 示合生面端部 Showing the end part of commissure. 3a-3e. 幼果后期果实横切面解剖结构Anatomical structure of fruit transection at the late stage of young fruit: 3a. 示果实全形Showing the whole fruit; 3b. 示主棱部位 Showing the main rib part; 3c. 示次棱部位 Showing the secondary rib part; 3d. 示分生果果壁 Showing the carpodermis of mericarp; 3e. 示合生面油管 Showing the vittae in commissure.
2.3 果熟发育阶段的解剖结构特征
2.3.1 果熟发育阶段解剖结构特征总体变化 芫荽果熟发育阶段各时期果实横切面解剖结构见图3。
与幼果发育阶段相比,整个果熟发育阶段果实横切面由近圆形变为圆形。果实内部2个分生果间的合生面在熟果后期最终形成较宽的分离断面,但心皮柄维管束以及合生面两端的外果壁和中果壁外侧数层细胞仍然相连。相对于果体宽度而言果壁更薄,内果壁木质化;主棱和次棱共同存在并微向外突出;果壁空腔逐渐减少,最终几近消失;中果壁维管束的导管数量基本维持恒定,其大小与同一时期的木化细胞相近;胚乳形状渐变成弯肾形。另外,中果壁细胞、胚乳腹面解体的细胞和合生面细胞中的结晶逐渐消失,但靠胚乳一侧的种皮内存在色素层。
2.3.2 熟果前期解剖结构特征 熟果前期的果实横切面全形及不同部位的解剖结构见图3-1a~1e。与幼果后期相比,这一时期正常发育的分生果有8个明显的变化:1)果实横切面呈椭圆形;分生果横切面长度增加至3 710 μm、宽度增加至1 540 μm。2)果壁厚度约为280 μm,占分生果宽度的18%。3)随果体的膨大,中果壁内木化细胞层的波状弧度以及主棱和次棱的差异均进一步减小,使棱状突起不明显。4)内果壁木质化,并在腹面中部形成1个“V”字形尖角,向合生面突出。5)种皮与胚乳间出现少量色素层。6)胚乳背部平滑,腹面微凹,呈肾形。7)合生面仅由心皮柄维管束和木质化中果壁至外果壁这一区域相连,其余部位的细胞均已破损。8)中果壁、胚乳和合生面细胞中的结晶基本消失。
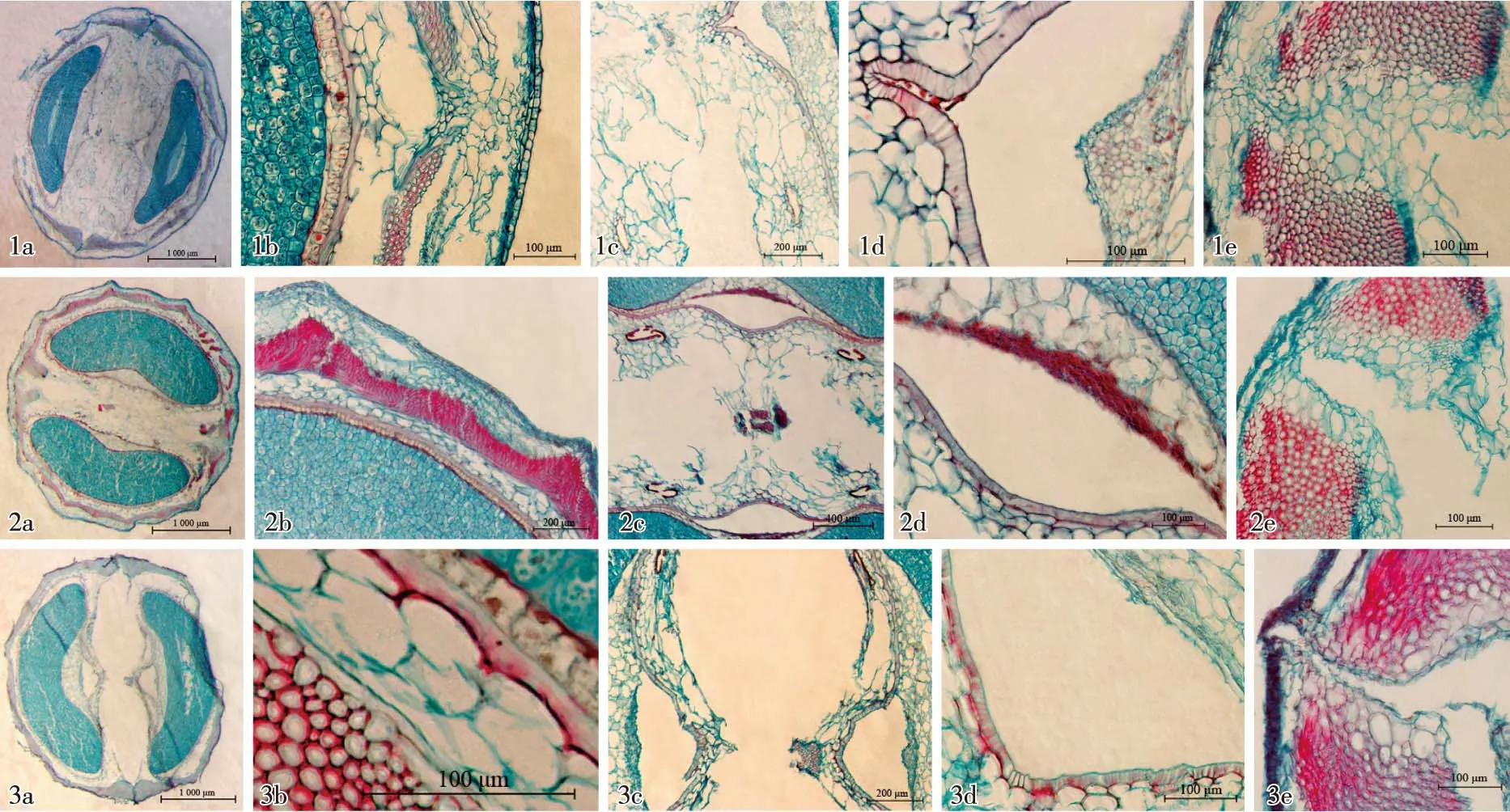
1a-1e. 熟果前期果实横切面解剖结构Anatomical structure of fruit transection at the early stage of mature fruit: 1a. 示果实全形 Showing the whole fruit; 1b. 示背部果壁Showing the carpodermis in dorsal part; 1c. 示合生面中部 Showing the middle part of commissure; 1d. 示腹面内果壁Showing the endocarp in ventral part; 1e. 示合生面端部 Showing the end part of commissure. 2a-2e. 熟果中期果实横切面解剖结构Anatomical structure of fruit transection at the middle stage of mature fruit: 2a. 示果实全形 Showing the whole fruit; 2b. 示背部Showing the dorsal part; 2c. 示合生面中部 Showing the middle part of commissure; 2d. 示腹面内果壁Showing the endocarp in ventral part; 2e. 示合生面端部 Showing the end part of commissure. 3a-3e. 熟果后期果实横切面解剖结构Anatomical structure of fruit transection at the late stage of mature fruit: 3a. 示果实全形 Showing the whole fruit; 3b. 示木化细胞至种皮部位Showing the part from lignified cells to episperm; 3c. 示合生面中部 Showing the middle part of commissure; 3d. 示腹面内果壁Showing the endocarp in ventral part; 3e. 示合生面端部 Showing the end part of commissure.
2.3.3 熟果中期解剖结构特征 熟果中期的果实横切面全形及不同部位的解剖结构见图3-2a~2e。与熟果前期相比,这一时期正常发育的分生果有7个明显变化:1)果实横切面由近圆形变为圆形;分生果横切面长度缩小至3 050 μm,宽度增加至1 600 μm。2)果壁厚度约为260 μm,占分生果宽度的16%。3)随分生果果体向外膨大,中果壁内木化细胞层的波状弧度进一步减小,但主棱和次棱部位的隆起比较明显(果实外表面可见微小果棱),而且次棱部位的隆起大于主棱部位。4)腹面中部的内果壁尖角变得平缓。5)胚乳呈肾形,其外侧与种皮间的色素层颜色变深。6)油管进一步废退和破损,几近消失。7)合生面进一步解体分离,两端的中果壁木质化细胞层也逐渐分开。
2.3.4 熟果后期解剖结构特征 熟果后期的果实横切面全形及不同部位的解剖结构见图3-3a~3e。与熟果中期相比,这一时期正常发育的分生果有5个明显变化:1)果实横切面呈近圆形或椭圆状圆形;分生果横切面长度增至3 290 μm、宽度减至1 580 μm。2)果壁变薄,厚度约为190 μm,仅占分生果宽度的12%。3)随着整个果体的膨大,果壁内的中果壁薄壁细胞层大多被破坏或消失,仅剩木化细胞层,后者在合生面处呈蹄状靠合。4)分生果腹面内凹,但在合生面中部的心皮柄维管束处向外呈山状突起,而且内果壁与胚乳的腹面间出现三角状空腔。5)在2个分生果间的合生面部位形成较宽的分离断面, 2个分生果仅由心皮柄维管束和合生面两端的外果壁及中果壁最外侧数层细胞相连。
3 讨 论
根据上述观察结果,结合伞形科果实解剖结构的相关研究报道[8,21-23,26-30],可认为芫荽果实是伞形科植物果实中比较特殊的一类,其特殊性主要表现在以下几个方面:
3.1 芫荽果实属于隐性双悬果类型
伞形科植物的果实为双悬果,可分为多种类型[26],其中大部分种类的果实在成熟后2个分生果从合生面处完全分离,并分别悬挂在分裂的心皮柄顶端;但也有因心皮柄分裂不完全或完全不分裂而导致2个分生果靠合在一起的类型,或心皮柄分裂或不分裂但合生面不分离的类型。然而芫荽果实较为特殊,其心皮柄不分裂,而合生面绝大部分已分离,但其分生果并不分离,这一类型在伞形科中很少见,是较为特殊的果实类型。虽然从外部形态上看芫荽果实是一个整体,但从横切面解剖结构上看明显是2个分生果,2个分生果没有分开仅因心皮柄维管束以及合生面两端的外果壁和中果壁最外侧2~3层细胞相连。由此可见,虽然芫荽果实表象上看不是双悬果,但从解剖结构上看属于真正意义上的双悬果,因此可称为“隐性双悬果”。
3.2 芫荽果实属于果棱退化类型
在伞形科果实中,根据果棱下部有无维管束常将果棱分为主棱和次棱2类,其中具有维管束的果棱为主棱、没有维管束的果棱为次棱,后者常位于2个维管束之间的部位。伞形科大多数种类的果实仅具5个主棱;也有的种类主棱和次棱并存,如矮泽芹属(ChamaesiumWolff)种类;少数种类仅具次棱,如胡萝卜属(DaucusLinn.)种类。据文献[8,24]记载,芫荽果实属于主棱和次棱并存的类型,共有11个棱,从果实表面看其中的5个主棱呈波状、6个次棱通直。
果棱及其维管束是伞形科分生果中的重要组织结构,而且二者关系密切。通常维管束多位于果棱发生的位置,而且是主棱发育的位置。但芫荽果实比较特殊,其维管束散布于果壁油管下方的中果壁第2层次的木化细胞层中,而且维管束很小、仅见导管。在子房期,由于主棱位置上方有大型果壁油管的阻碍,主棱不能正常发育。在果期,虽然由这些油管转而形成的大型空腔发生了塌陷或消失,但形成的主棱却很小;相反,位于空腔之间的中果壁向外突起形成相对明显的次棱(其中没有维管束)。
芫荽果实的果棱变化比较复杂,而且主棱和次棱的形成与子房期油管(另文发表)及其后来在果期演变为大型空腔的发育过程有较大的关系。由于主棱位于空腔下部,在果实发育初期果实外表面呈现出宽大果棱状隆起,这些隆起虽然位于维管束位置,但并不是真正的果棱而是由空腔造成的。随果壁油管的消失,位于其下方维管束部位的木化细胞层向外凸起并渐渐形成果棱状结构——主棱,但从果实外表面上看主棱呈波状弯曲,其致因尚有待进一步研究。
另外,从果棱的发达程度看,伞形科植物的双悬果具有无/微果棱、果棱不甚发达和果棱发达或很发达等多样形态。芫荽果实的果棱很小并且次棱比主棱明显,属子果棱退化类型的果实。
3.3 芫荽果实具有特殊的果壁油管
油管是伞形科植物果实的一个重要特征,也是该科植物分类的主要依据之一[26]。伞形科芹亚科种类的油管主要包括果壁油管和合生面油管2类,前者位于2个维管束形成的果棱之间(即棱槽内),后者位于合生面中,有时在果棱维管束的上方还有伴生小油管(即果棱油管)。但芫荽果实中的油管比较特殊:1)在果期,特别是果熟期,虽然看不到果壁油管,但是可以看到退化的小型合生面油管,而且在子房期明显可以看到果壁油管,所以前人得出的“芫荽果实没有油管[1]”的结论并不确切。2)果壁油管的发育经历了先盛后衰的过程,在子房期油管正常发育,但在子房末期油管开始解体,至幼果期油管演变为空腔并不断扩大,从幼果中后期开始大型空腔逐渐减退和消失。3)无论子房期的果壁油管、还是果期的果壁油管空腔,其位置均在维管束上方,而非棱槽(或维管束)之间,与大部分芹亚科果实的果壁油管或棱槽油管的着生位置明显不同,而与变豆菜亚科(Saniculoideae Drude)的种类十分相似[21]。4)芫荽果实含有丰富的精油[31],但从解剖结构看,果期仅合生面有油管,因此精油只可能来源于合生面油管,而且以果实未完全成熟时含量较高。5)果期油管消退,在果壁油管着生的相同位置发育出大型空腔,其发育机制有待进一步研究。
3.4 芫荽果实具有发达的中果壁
伞形科芹亚科有很多种类的果实的中果壁细胞有分层现象,并且部分种类的中果壁细胞发生木化。但芫荽果实的中果壁细胞层次和木质化程度都很高,类似特征在伞形科中比较少见。从细胞分层看,芫荽果实的中果壁由外向内可分为4个层次,而且各层次的结构特征区别明显,其中第1和第2层次具有重要的作用。第1层次的最外层细胞非常致密,细胞内外壁增厚,与外果壁相似而紧紧贴合,而且分生果分离时在合生面处始终不断裂,这也是导致芫荽分生果不分离的主要原因之一。第2层次为木化细胞层,细胞多呈狭长纤维状,其层内细胞呈不同走向排列,而且整个层带围绕分生果果体呈折线状分布,这是导致芫荽果实果壁十分坚硬的重要因素,也是形成果棱的来源和基础。
3.5 芫荽果实的维管束分散而退化
根据维管束的着生部位,芫荽果实内的维管束主要可分为3类:第1类是位于中果壁的果壁维管束;第2类是位于合生面的心皮柄维管束;第3类是位于胚乳腹面外侧处的种脊维管束。通常维管束都形成一个整体并与周围细胞有明显界限,但芫荽果壁中的维管束导管数量少且没有聚集在一起,而是单个分布或仅1~2个聚集,分别散布于中果壁的木化细胞层中,这种维管束的特征与伞形科Marlothiella属种类果实的维管束[32]相似。推测这种分散的维管束是芫荽果实主棱不发达的原因之一。
3.6 芫荽果实细胞的结晶先盛后衰
结晶类型和分布部位常作为伞形科植物系统分类的重要依据之一[21,27]。芫荽果实细胞中也含有大量结晶,但仅出现在幼果阶段,此时果壁、合生面和解体的胚乳腹面细胞中都有大量簇晶;而胚乳细胞中则有大量的糊粉粒结晶;但至熟果期,中果壁、解体的胚乳腹面和合生面细胞中的簇晶则几乎不可见。芫荽果实细胞中这种结晶出现后又消失的现象是否普遍存在于芹亚科的其他种类中,还需从果实解剖结构方面进行进一步研究和探讨。另外,刘启新等[21]的研究结果表明:伞形科变豆菜亚科变豆菜属(SaniculaLinn.)种类的中果壁薄壁细胞中散布有大小不等的放射状簇晶。因而,芫荽果实中果壁细胞内的簇晶与变豆菜亚科的种类较为接近,而其胚乳细胞中的糊粉粒结晶又与芹亚科的部分种类[30]相似。
参考文献:
[1] 中国科学院中国植物志编辑委员会. 中国植物志: 第五十五卷第一分册[M]. 北京: 科学出版社, 1979: 87-91.
[2] KAUR P, KUMAR A, ARORA S, et al. Quality of dried coriander leaves as affected by pretreatments and method of drying[J]. European Food Research and Technology, 2006, 223(2): 189-194.
[3] EMAMGHOREISHI M, KHASAKI M, AAZAM M F.Coriandrumsativum: evaluation of its anxiolytic effect in the elevated plus-maze[J]. Journal of Ethnopharmacology, 2005, 96(3): 365-370.
[4] 戴国彪, 姜子涛, 李 荣, 等. 芫荽籽精油抗氧化能力研究[J]. 食品研究与开发, 2010, 31(8): 8-11.
[5] ZOUBIRIS,BAALIOUAMERA.EssentialoilcompositionofCoriandrumsativumseed cultivated in Algeria as food grains protectant[J]. Food Chemistry, 2010, 122(4): 1226-1228.
[6] 黄雅琴, 黄群策, 赵帅鹏, 等. 反季节芫荽种子在不同条件下的萌发状态研究[J]. 北方园艺, 2008(9): 10-13.
[7] OKUT N, YIDIRIM B. Effects of different row spacing and nitrogen doses on certain agronomic characteristics of coriander(CoriandrumsativumL.)[J]. Pakistan Journal of Biological Sciences, 2005, 8(6): 901-904.
[8] DIEDERICHSEN A. Coriander (CoriandrumsativumL.). Promoting the Conservation and Use of Underutilized and Neglected Crops. 3[M]. Gatersleben: Institute of Plant Genetics and Crop Plant Research; Rome: International Plant Genetic Resources Institute, 1996.
[9] KUMAR A, SINGH R, CHHILLAR R K. Influence of omitting irrigation and nitrogen levels on growth, yield and water use efficiency of coriander (CoriandrumsativumL.)[J]. Acta Agronomica Hungarica, 2008, 56(1): 69-74.
[10] SILVA F, FERREIRA S, QUEIROZ J A, et al. Coriander (Co-riandrumsativumL.) essential oil: its antibacterial activity and mode of action evaluated by flow cytometry[J]. Journal of Medical Microbiology, 2011, 60(10): 1479-1486.
[11] MATASYOH J C, MAIYO Z C, NGURE R M, et al. Chemical composition and antimicrobial activity of the essential oil ofCoriandrumsativum[J]. Food Chemistry, 2009, 113(2): 526-529.
[12] AISSAOUI A, ZIZI S, ISRAILI Z H, et al. Hypoglycemic and hypolipidemic effects ofCoriandrumsativumL. in Meriones shawi rats[J]. Journal of Ethnopharmacology, 2011, 137(1): 652-661.
[13] MAHENDRA P, BISHT S. Anti-anxiety activity ofCoriandrumsativumassessed using different experimental anxiety models[J]. Indian Journal of Pharmacology, 2011, 43(5): 574-577.
[14] 陆占国, 郭红转, 李 伟. 芫荽根部芳香成分研究[J]. 化学与黏合, 2009, 29(2): 79-81.
[15] BURDOCK G A, CARABIN I G. Safety assessment of coriander (CoriandrumsativumL.) essential oil as a food ingredient[J]. Food and Chemical Toxicology, 2009, 47(1): 22-34.
[16] PRAKASH V. Leafy Spices[M]. Boca Raton: CRC Press, 1990.
[17] De SOUZA E L, STAMFORD T L M, LIMA E O, et al. Anti- microbial effectiveness of spices: an approach for use in food conservation systems[J]. Brazilian Archives of Biology and Technology, 2005, 48(4): 549-558.
[18] 滕红梅, 胡正海. 芫荽花蜜腺的发育解剖学研究[J]. 西北植物学报, 1998, 18(1): 41-46.
[21] 刘启新, 惠 红, 李碧媛, 等. 中国伞形科变豆菜亚科的果实解剖特征及其系统学意义[J]. 植物资源与环境学报, 2002, 11(4): 1-8.
[22] 王长宝, 马祥光, 何兴金. 伞形科柴胡属部分物种的果实特征及系统学意义[J]. 植物科学学报, 2011, 29(4): 399-408.
[23] LIU M, Van WYK B E, TILNEY P M. The taxonomic value of fruit structure in the Chinese endemic genusDickinsia(Apiaceae)[J]. Nordic Journal of Bonaty, 2002, 22(5): 603-607.
[24] 中国医学科学院药物研究所, 中国科学院南京中山植物园, 北京医学院药学系, 等. 中药志: 第二册[M]. 北京: 人民卫生出版社, 1959.
[25] 李正理. 植物制片技术[M]. 2版. 北京: 科学出版社, 1987.
[26] LIU M, PLUNKETT G M, Van WYK B E, et al. The phylogenetic significance of the carpophore in Apiaceae[J]. Annals of Botany, 2012, 110(8): 1531-1543.
[27] LIU M, Van WYK B E, TILNEY P M. Irregular vittae and druse crystals inSteganotaeniafruits support a taxonomic affinity with the subfamily Saniculoideae Apiaceae [J]. South African Journal of Botany, 2007, 73(2): 252-255.
[28] 宋春凤, 刘玉龙, 刘启新, 等. 伞形科植物明党参花后果实发育的解剖结构变化[J]. 植物资源与环境学报, 2011, 20(4): 1-7.
[29] 刘启新, 惠 红, 潘泽惠, 等. 中国伞形科天胡荽亚科果实解剖特征及其系统学意义[J]. 植物资源与环境学报, 2002, 11(3): 1-7.
[30] 何兴金, 王幼平, 溥发鼎, 等. 中国独活属果实的解剖学研究及对独活属的修订[J]. 云南植物研究, 1998, 20(3): 295-302.
[31] MSAADA K, TAARIT M B, HOSNI K, et al. Regional and maturational effects on essential oils yields and composition of coriander (CoriandrumsativumL.) fruits[J]. Scientia Horti-culturae, 2009, 122(1): 116-124.
[32] Van WYK B E, TILNEY P M. A revision of the genusMarlothiella(Apiaceae)[J]. South African Journal of Botany, 2007, 73(2): 208-213.
