海洋细菌Bacillus mojavensis 1A00437 β-1,3-1,4-葡聚糖酶基因的克隆、表达及功能研究
2014-04-02左怀雨王茂淋邵宗泽李光玉喻子牛张吉斌
左怀雨,王茂淋,邵宗泽,李光玉,徐 柳,喻子牛,张吉斌
(1.华中农业大学生命科学技术学院 农业微生物学国家重点实验室 微生物农药国家工程研究中心,湖北 武汉430070;2.国家海洋局第三海洋研究所 海洋生物遗传资源重点实验室,福建 厦门 361005)
β-1,3-1,4-葡聚糖是禾本科植物细胞壁的重要组成部分。β-1,3-1,4-葡聚糖酶(EC3.2.1.73)是解聚β-1,3-1,4-D-葡聚糖的活性最高的内切酶,属于糖基水解酶16家族(GHF16),可以精确水解和裂解地衣多糖或β-D-葡聚糖中位于β-1,3-糖苷键后的β-1,4-糖苷键。在啤酒发酵中,高分子量β-葡聚糖的残留会导致啤酒原液黏度和浊度的增大[1],黏度的增大会降低麦芽汁的过滤速率,而β-1,3-1,4-葡聚糖酶减小降低麦芽汁黏度。β-1,3-1,4-葡聚糖酶还被广泛用作饲料添加剂,以大麦为主的饲料中添加β-1,3-1,4-葡聚糖酶,饲料的能量利用率可提高13%,蛋白质利用率提高21%[2],显著提高了饲料利用率。
β-1,3-1,4-葡聚糖酶主要存在于一些植物组织中,如大麦、稻米等[3];另外,也来源于许多芽胞杆菌[3],如Bacillussubtilis、Bacillusamyloliquefaciens、Bacilluslicheniformis、Bacilluscirculans等,以及一些瘤胃细菌[4],如Ruminococcusflavofaciens、Bacteroidessuccinogenes、StreptococcusbovisJB1[5]等;最近,在一些真菌、高等植物中也有发现。
细菌β-1,3-1,4-葡聚糖酶在不同的宿主细胞中的表达均有报道,如Escherichiacoli、Bacillusspp.、Saccharomycescerevisiae、Pichiapastoris以及转基因大麦和烟草植物等。E.coli作为常规的原核表达宿主,常用于细菌基因的克隆和表达。Bacillusmojavensis1A00437是一株分离自太平洋的芽胞杆菌,其生长环境的特殊性使其具有特殊的研究价值。作者克隆了Bacillusmojavensis1A00437β-1,3-1,4-葡聚糖酶基因并在大肠杆菌中进行表达,研究其酶学特点,以期获得具有工业生产应用价值的葡聚糖酶。
1 实验
1.1 材料与试剂
Bacillusmojavensis1A00437由国家海洋局第三海洋研究所提供;E.coliDH5α、E.coliBL21(DE3)、小麦纹枯病菌(RhizoctoniasolaniAG8)及水稻纹枯病菌(RhizoctoniasolaniKühn),自行保存。
AxyPrepTMDNA Gel Extraction Kit、AxyPrepTMPCR Cleanup Kit,美国Axygen公司;Plasmid Midiprep Kit,北京庄盟国际生物基因科技有限公司;T4 DNA Ligase、Ex TaqTMPolymerase、RNase A、EcoRⅠ、NotⅠ,大连宝生物工程有限公司;pMD18-T、pET-28a,Takara公司;Yeast extract、Typtone,Oxiod公司;Agarose,Biowest公司;β-葡聚糖,Sigma公司;其它试剂均为国产分析纯。
1.2 方法
1.2.1引物设计与合成
通过在NCBI数据库上进行序列搜索及相似性比对,依据已登录的β-1,3-1,4-葡聚糖酶基因序列设计兼并引物并委托Invitrogen公司合成。引物序列如下:
bgl1:5′-ATGTCTTATCGTATGAAACGAGT-3′;
bgl2:5′-TTATTTTTTTGTATARCGYA-3′;
R=A/G,Y=C/T。
1.2.2基因组DNA 的制备及基因克隆
Bacillusmojavensis1A00437基因组DNA 制备参照细菌DNA抽提试剂盒说明书进行。以基因组DNA 为模板进行PCR 扩增。程序如下:94 ℃预变性5 min,94 ℃ 30 s,55 ℃ 30 s,72 ℃ 45 s;30 个循环后72 ℃延伸10 min。PCR 产物经1%琼脂糖凝胶电泳后回收并纯化(按照AxyPrepTMPCR Cleanup Kit试剂盒说明书进行)。回收产物与pMD18-T 载体连接过夜,连接产物转化Ca2+法制备的感受态细胞,37 ℃培养后菌落PCR 鉴定,阳性克隆质粒由上海英俊生物技术有限公司进行测序。
1.2.3序列分析
将测序得到的Bacillusmojavensis1A00437β-1,3-1,4-葡聚糖酶基因序列用softberry、ORF finder等软件分析其开放阅读框,用BLASTEN (http://www.ncbi.nlm.nih.gov/BLASTEN)进行序列相似性比对,Moe软件进行酶的结构模拟和关键作用位点的分析。
1.2.4原核表达
根据阳性克隆测序结果,设计β-1,3-1,4-葡聚糖酶成熟表达引物。在引物5′端加入EcoRⅠ和NotⅠ酶切位点,并委托Invitrogen公司合成。引物序列如下:
Fbgl1:5′-CCGGAATTCATGAGTTTGTCTGCAGTCACTTCTA-3′;
Rbgl2:5′-AATGCGGCCGCTTTTTTTGTATAGCGCACCCAG-3′。
以带有目的基因的重组质粒为模板进行PCR 扩增。程序如下:94 ℃预变性5 min,94 ℃ 30 s,55 ℃ 30 s,72 ℃ 45 s;30 个循环后72 ℃延伸10 min。PCR产物经1%琼脂糖凝胶电泳检测并回收纯化,同时与载体pET-28a进行EcoRⅠ及NotⅠ双酶切处理后于16 ℃连接过夜。转化E.coliDH5α感受态细胞及BL21感受态细胞。选取阳性克隆,经菌液PCR鉴定及双酶切鉴定正确无误后,以IPTG为诱导剂进行诱导表达,25 ℃诱导3 h。表达产物用SDS-PAGE检测。
1.2.5表达产物的分离纯化
大量诱导表达目的蛋白,4 ℃、5 000 r·min-1离心3 min收集菌体,去上清后加入1/10原培养液体积的Binding buffer悬浮细胞,超声破碎。收集粗蛋白液,4 ℃、12 000 r·min-1离心30 min,吸取上清液于干净的离心管中,用滤纸过滤后置于4 ℃备用。蛋白纯化按照GE公司的金属镍螯合琼脂糖凝胶6FF操作手册进行,SDS-PAGE 检测各段收集样品。对目的蛋白进行透析复性,用于后续实验。
1.2.6β-1,3-1,4-葡聚糖酶的酶活力测定
酶活力单位定义:在40 ℃、pH值5.5条件下,每分钟从浓度为5 mg·mL-1的β-葡聚糖溶液中释放1 μmol还原糖所需的酶量定义为一个酶活力单位(1 U)。
将1 mL稀释后的酶液和1 mL 8.0 g·L-1β-葡聚糖放入40 ℃水浴锅平衡30 min。采用DNS法测定还原糖含量。
1.2.7β-1,3-1,4-葡聚糖酶的酶学性质
通过pH值为4.0~8.0的乙酸-乙酸钠缓冲溶液稀释酶液测定最适pH值;pH值稳定性通过β-1,3-1,4-葡聚糖酶在pH值5.0、5.5、6.0、6.5、7.0的缓冲溶液中平衡不同时间(0 min,5 min,10 min,15 min,20 min,25 min,30 min)后测定,以0 min时的酶活为100% 酶活。稀释酶液与8.0 g·L-1β-葡聚糖底物混匀,50 ℃反应30 min,用DNS法测定并计算相对酶活。
将稀释酶液分别在20~100 ℃平衡30 min测定最适温度;将稀释酶液在40 ℃、50 ℃、60 ℃、70 ℃、80 ℃中分别保温0 min、10 min、20 min、30 min、40 min、50 min、60 min后测定热稳定性,以0 min时的酶活为100% 酶活;将稀释酶液与底物混合,50 ℃反应30 min,用DNS法测定并计算相对酶活。
金属离子与化学试剂的影响测定:在酶促反应体系中分别加入终浓度为1 mmol·L-1的金属离子(Zn2+、Cu2+、Mg2+、Ca2+、K+、Ni2+、Mn2+、Fe2+、Li+、Na+)和化学试剂(EDTA、SDS),以未加金属离子和化学试剂的酶活为100% 酶活,用DNS法测定并计算相对酶活。
底物专一性测定:选取β-葡聚糖、木聚糖、甘露聚糖、几丁质作为底物并配制成浓度为8.0 g·L-1的溶液,用DNS法测定并计算各底物反应条件下的相对酶活。
1.2.8β-1,3-1,4-葡聚糖酶抗真菌活性测定
采用打孔对峙法,将病原真菌菌块置于新鲜的固体培养基中央,并在培养基上用5 mm直径打孔器均匀打孔6个,挑出琼脂块备用。待真菌菌丝生长至6个孔约1 cm处,吸取一定量的不同蛋白液和无菌水,分别加入到琼脂平板已经制备好的孔中。培养48 h,观察抑制效果。
1.2.9β-1,3-1,4-葡聚糖酶对饲料消化率的影响
将麸皮、稻糠、玉米粉分别过100目筛,将稀释后的酶液和相应的饲料底物混匀放入50 ℃水浴锅平衡5 h。分别设置空白对照。通过DNS法测定还原糖含量,以确定β-1,3-1,4-葡聚糖酶对饲料消化率的影响。
2 结果与讨论
2.1 bgl基因克隆
以Bacillusmojavensis1A00437的总DNA为模板,经PCR扩增得到目的片段,约为700 bp(图1),与预测的大小相符。扩增后得到的PCR产物经过纯化后与pMD-18T载体连接,转化E.coliDH5α,菌落生长后经菌落PCR验证(图2),结果正确。
2.2 bgl序列分析
将测序得到的基因序列用softberry(http://linux1.softberry.com/berry.phtml)、ORF finder (http://www.ncbi.nlm.nih.gov/gorf/gorf.html)等软件分析其开放阅读框,最后确定该酶基因的ORF为681 bp;NCBI序列比对后构建进化树(图3),分析Bacillusmojavensis来源的bgl基因的同源性。

M.DL 20001,2.target gene
图1Bacillusmojavensis1A00437中bgl基因的克隆
Fig.1bglGeneclonedinBacillusmojavensis1A00437
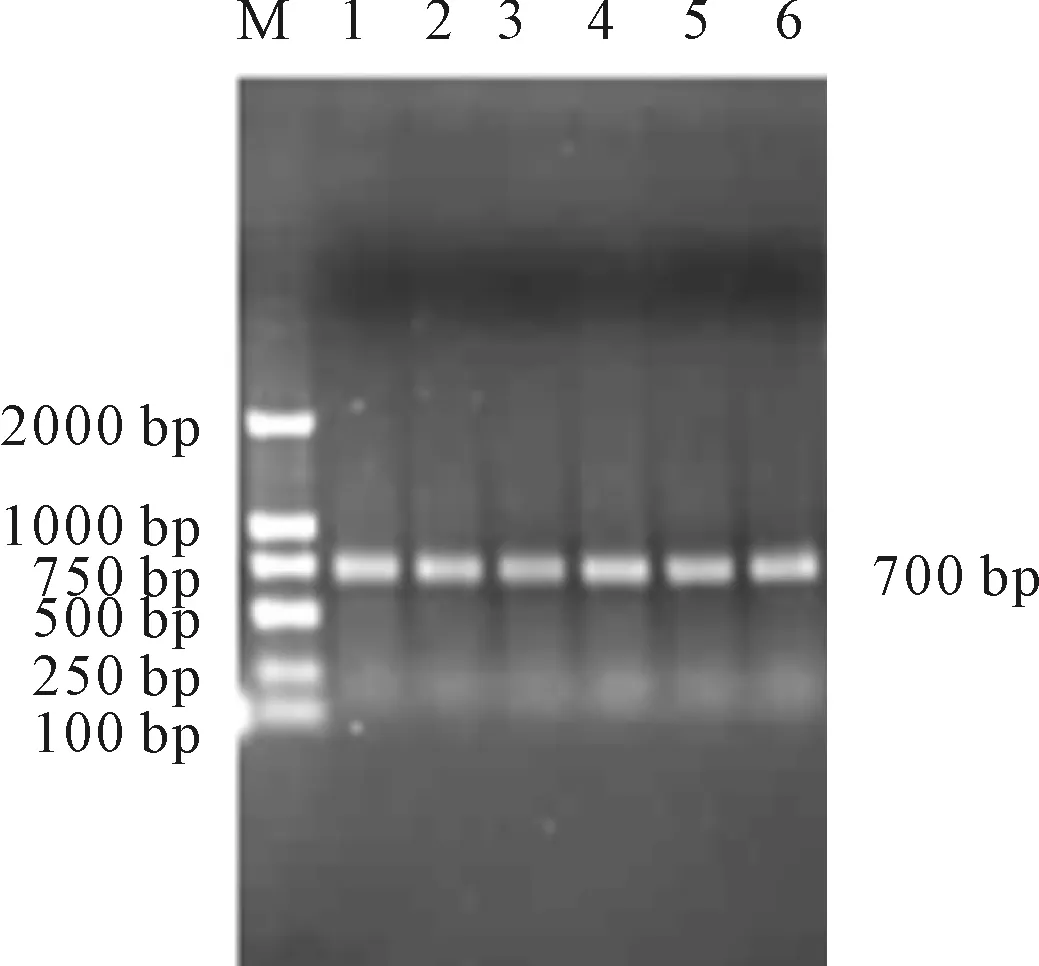
M.DL 2000 1~6.different colonies verified by colony PCR

图3 β-1,3-1,4-葡聚糖酶的序列进化树
由图3可以看出,Bacillusmojavensis来源的bgl基因与Bacilluslicheniformis来源的bgl基因具有高度的相似性,可达99%。
通过对氨基酸序列的分析,确定该酶属于糖基水解酶16家族成员,理论pH值为6.05,蛋白大小为25.3 kDa,β-1,3-1,4-葡聚糖酶的保守区域为E-[LIV]-D-[LIVF]-x(0,1)-E-x(2)-[GQ]-[KRNF]-x-[PSTA],其中2个E是关键作用位点,本研究中酶的保守区域为EIDIEFLGKDT,2个通过粗体标明的E为该酶的活性位点氨基酸。该酶的保守区域和活性位点氨基酸如图4所示。其中,保守区域以灰色表示,活性位点为Glu 133和Glu 137。

A.the front of substrate binding pocket
2.3 目的基因亚克隆
分别使用限制性内切酶EcoRⅠ和NotⅠ双酶切pMD18-T-bgl(E,N)和表达载体pET28a,然后将含有酶切位点的目的基因bgl和载体片段进行连接,得到重组质粒pET28a-bgl(E,N)。构建好的载体图谱如图5所示。

图5 重组质粒pET28a-bgl(E,N)的构建
将构建好的pET28a-bgl(E,N)用EcoRⅠ和NotⅠ进行双酶切验证,结果见图6。
由图6可知,pET28a-bgl(E,N)经EcoRⅠ和NotⅠ双酶切后得到大小约为681 bp的bgl基因片段和大小约为5.3 kb的载体片段。载体测序结果表明,所有的目的基因在重组质粒中的克隆方向、插入位点和阅读框架正确。

M.DL20001,2.double-digest of pET28a-bgl(E,N)
图6重组质粒pET28a-bgl(E,N)双酶切
Fig.6DoubledigestionofrecombinantexpressionvectorpET28a-bgl(E,N)
2.4 β-1,3-1,4-葡聚糖酶分离纯化(图7)

M.DL2000 1.the whole proteins of Bgl not-induced by IPTG
2.the whole proteins of Bgl induced by IPTG at 25 ℃3.purity protein
图7SDS-PAGE分析Bgl蛋白的表达
Fig.7AnalysisofBglproteinexpressionbySDS-PAGE
由图7可知,经IPTG诱导表达后,利用His标签分离纯化后得到目的蛋白,融合表达的目的蛋白的大小与预测大小相近。
2.5 β-1,3-1,4-葡聚糖酶的生物学特性
2.5.1pH值对β-1,3-1,4-葡聚糖酶的酶活力和稳定性的影响(图8)
由图8a可知,β-1,3-1,4-葡聚糖酶在pH值6.5的反应体系中具有最高的酶活力。
由图8b可知,β-1,3-1,4-葡聚糖酶在pH值5.0~7.0 的反应体系中保持70% 以上的酶活力,并且在pH值5.5的反应体系中相对酶活最高达到100%以上。
2.5.2温度对β-1,3-1,4-葡聚糖酶的酶活力和稳定性的影响(图9)

图8pH值对β-1,3-1,4-葡聚糖酶的酶活力(a)和稳定性(b)的影响
Fig.8EffectofpHvalueonactivity(a)andstability(b)ofβ-1,3-1,4-glucanase

由图9a可知,β-1,3-1,4-葡聚糖酶在60 ℃时具有最高的酶活力。
由图9b可知,β-1,3-1,4-葡聚糖酶在40~50 ℃区域内保温60 min后酶活力保持在80%以上;但当温度提高到60 ℃时,随着保温时间的延长酶活力发生波动,60 min后酶活力下降到30%以下。表明该酶在40~50 ℃时比较稳定,而在70~80 ℃时,酶活力在10 min内迅速下降到30%以下,极不稳定。
2.5.3金属离子及化学试剂对β-1,3-1,4-葡聚糖酶的酶活力的影响(图10)

图10 金属离子和化学试剂对β-1,3-1,4-葡聚糖酶的酶活力的影响
由图10可知,Ni2+和Fe2+显著增强酶活力,Cu2+、Ca2+、Mn2+对酶活力有明显的抑制作用,其余金属离子和化学试剂对酶活力无显著影响。
2.5.4β-1,3-1,4-葡聚糖酶底物专一性测定(表1)
表1不同底物的酶活力/(U·mL-1)
Tab.1Theenzymeactivityofdifferentsubstrates/(U·mL-1)

底物名称β⁃葡聚糖木聚糖甘露聚糖几丁质酶活力16400001700200021
由表1可知,木聚糖、甘露聚糖、几丁质与β-葡聚糖相比,酶解效率极低,表明β-1,3-1,4-葡聚糖酶具有严格的底物专一性。
2.6 β-1,3-1,4-葡聚糖酶的功能研究
2.6.1β-1,3-1,4-葡聚糖酶抗真菌活性测定(图11)
由图11可知,β-1,3-1,4-葡聚糖酶对小麦纹枯病菌及水稻纹枯病菌菌丝的生长有显著的抑制作用。
2.6.2β-1,3-1,4-葡聚糖酶对饲料消化率的影响(表2)
由表2可知,β-1,3-1,4-葡聚糖酶对稻糠、玉米粉具有一定的降解作用,能够分解其细胞壁结构,利于饲料释放其内容物,以提高饲料的能量利用率。该实验为β-1,3-1,4-葡聚糖酶用作饲料添加剂提供了理论基础。

A.水稻纹枯病菌 B.小麦纹枯病菌
表2β-1,3-1,4-葡聚糖酶降解饲料效果
Tab.2Feeddigestionresultsofrecombinantβ-1,3-1,4-glucanase

饲料稻糠玉米粉麸皮还原糖得率/%12716304
2.7 讨论
邱思鑫等[6]研究发现,0.1~0.5 mmol·L-1的Ca2+对β-1,3-1,4-葡聚糖酶的酶活力有显著促进作用,0.2~0.4 mmol·L-1Fe2+可提高该酶的酶活力。谢焱等[7]研究发现,10 mmol·L-1的 Ca2+对β-1,3-1,4-葡聚糖酶的酶活力无显著影响,10 mmol·L-1的Fe2+对该酶的酶活力有显著的抑制作用。本实验研究发现,1 mmol·L-1Ca2+对酶活力有明显的抑制作用,1 mmol·L-1Fe2+对酶活力有显著促进作用。另外,Ni2+也可显著增强酶活力。造成这些差异的原因可能与菌株的生长环境相关,较特殊的海洋环境影响了菌株的某些遗传特性,在实际应用中可通过调节金属离子浓度提高酶活力。
姚乌兰等[8]从多粘类芽孢杆菌WY110(Paenibacilluspolymyxastrain WY110)菌株中分离纯化出一种β-1,3-1,4-葡聚糖酶P2蛋白,该酶具有抗稻瘟病菌的活性,而重组酶对稻瘟病菌无抗性,但对小麦纹枯病菌及水稻纹枯病菌具有明显抑制作用。P2蛋白与本研究中的重组酶相似性为77%,保守区域序列相同,不同之处应为蛋白结构序列。另有文献报道[9],重组β-1,3-1,4-葡聚糖酶对尖孢镰刀菌、香蕉炭疽病菌、荔枝炭疽病菌以及出芽短梗霉的菌丝生长具有明显的抑制活性。生物信息学分析发现,多数β-1,3-1,4-葡聚糖酶具有相同的保守区域及活性氨基酸,其功能不同可能是其蛋白结构不同造成的。造成该重组β-1,3-1,4-葡聚糖酶具有不同抗真菌活性的原因可能为酶的结构、底物专一性不同。另外,该重组β-1,3-1,4-葡聚糖酶对稻糠、玉米粉饲料具有一定的降解效果。
3 结论

参考文献:
[1]HAN F,ULLRICH S E,CHIRAT S,et al.Mapping ofβ-glucan content andβ-glucanase activity loci in barley grain and malt[J].Theoretical and Applied Genetics,1995,91(6-7):921-927.
[2]华卫东,史占全,严冰,等.β-葡聚糖酶在猪大麦基础日粮中的应用研究进展[J].饲料工业,1999,(8):15-17.
[3]YANG S Q,YAN Q J,JIANG Z Q,et al.Biochemical characterization of a novel thermostableβ-1,3-1,4-glucanase (lichenase) fromPaecilomycesthermophila[J].Journal of Agricultural and Food Chemistry,2008,56(13):5345-5351.
[4]BECKMANN L,SIMON O,VAHJEN W.Isolation and identification of mixed linkedβ-glucan degrading bacteria in the intestine of broiler chickens and partial characterization of respective 1,3-1,4-β-glucanase activities[J].Journal of Basic Microbiology,2006,46(3):175-185.
[5]EKINCI M S,MCCRAE S I,FLINT H J.Isolation and overexpression of a gene encoding an extracellularβ-(1,3-1,4)-glucanase fromStreptococcusbovisJB1[J].Applied and Environmental Microbiology,1997,63(10):3752-3756.
[6]邱思鑫,范晓静,胡方平,等.内生解淀粉芽胞杆菌β-1,3-1,4-葡聚糖酶基因(bglS)的克隆与表达及酶学特性分析[J].农业生物技术学报,2010,18(6):1173-1181.
[7]谢焱,顾国贤,李崎.重组β-1,3-1,4-葡聚糖酶的纯化及部分酶学性质研究[J].安徽农业科学,2008,36(20):8510-8512.
[8]姚乌兰,王云山,韩继刚,等.水稻生防菌株多粘类芽孢杆菌WY110抗菌蛋白的纯化及其基因克隆[J].遗传学报,2004,(9):878-887.
[9]文凤云,廖富蘋,林健荣,等.多粘类芽孢杆菌CP7β-1,3-1,4-葡聚糖酶基因的克隆、表达及应用[J].中国农业科学, 2010,(22):4614-4623.
