基于壳长频率分析的洪泽湖河蚬渔获物年龄结构、生长与死亡参数估计
2014-03-29毕婷婷侯刚张胜宇谢松光
毕婷婷侯 刚张胜宇谢松光
(1. 中国科学院水生生物研究所, 中国科学院水生生物多样性与保护重点实验室, 武汉 430072; 2. 中国科学院大学,北京 100049; 3. 广东海洋大学水产学院, 湛江 524025; 4. 江苏省洪泽湖渔业管理委员会办公室, 淮安223300; 5. 中国科学院水生生物研究所淮安研究中心, 苏北湖群渔业可持续发展与水环境保护重点实验室, 淮安 223002)
基于壳长频率分析的洪泽湖河蚬渔获物年龄结构、生长与死亡参数估计
毕婷婷1,2侯 刚1,3张胜宇4谢松光1,5
(1. 中国科学院水生生物研究所, 中国科学院水生生物多样性与保护重点实验室, 武汉 430072; 2. 中国科学院大学,北京 100049; 3. 广东海洋大学水产学院, 湛江 524025; 4. 江苏省洪泽湖渔业管理委员会办公室, 淮安223300; 5. 中国科学院水生生物研究所淮安研究中心, 苏北湖群渔业可持续发展与水环境保护重点实验室, 淮安 223002)
河蚬(Corbicula fluminea)广泛分布于东南亚、澳大利亚和非洲等地区的河流和淡水湖泊中[1,2], 它也是我国一些湖泊中(如太湖、洪泽湖)十分重要的经济贝类[3—6]。其中, 洪泽湖河蚬资源量曾达1×108kg以上, 分布遍及全湖除水草资源特别丰富的水域。但近年来, 由于外来污水进入湖体、过度捕捞以及捕捞生产中渔民2.27使用农药煤气等的影响,河蚬资源量下降, 2012年洪泽湖河蚬资源调查得出资源量为5.00×107kg, 比2011年的6.92×107kg下降28.80%, 比2010年的7.84×107kg下降36.20%, 分布区域明显缩小(江苏省洪泽湖渔业管理委员会办公室, 未发表数据)。
目前, 关于河蚬的研究主要集中在生物学特征、时空分布与环境因子的关系以及繁殖生物学特征方面[2,7—9],种群结构、生长和死亡的研究较少。种群的年龄结构、生长和死亡等基本参数是渔业资源管理的基础[10], 并且研究证明对于不能通过硬组织确定年龄的渔业对象, 体长频率分析方法是确定渔获物年龄结构的有效手段[10—12]。本文通过洪泽湖河蚬渔获物的壳长频数数据, 对河蚬渔获物的年龄结构、生长和死亡参数进行分析, 评估洪泽湖河蚬资源利用状况, 为制定合理的资源管理对策提供指导。
1 材料与方法
1.1 研究位置和采样方法
2011年8月至2012年7月逐月于洪泽湖河蚬主要分布水域(33°20′N,118°75′E)(图 1)采集河蚬, 采样工具为渔民捕捞河蚬用的耙栅, 孔径6 mm。河蚬洗净后置于敞口桶中、避光, 活体带回实验室进一步处理。
1.2 实验室处理
从每月采集的河蚬样品中随机选取195—200个个体,用游标卡尺测量壳长(L, 0.10 mm), 电子天平称取体重(W, 0.01 g), 采用幂函数方程拟合壳长-体重关系。
图1 洪泽湖中采样点的地理位置Fig. 1 Collection site of Corbicula fluminea in the Hongze Lake, Jiangsu Province, China
1.3 数据分析
根据河蚬生长的季节性特点, 采用von Bertalanffy季节性生长方程拟合河蚬生长方程[13,14]:
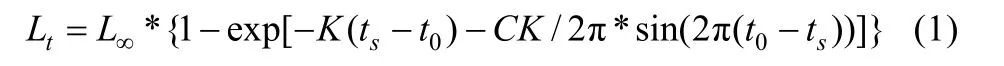
式中, L∞为渐近壳长, K为生长曲线的平均速率, t0为假设的理论生长起点年龄, ts为“夏季点”取0和1之间的值, C为季节性波动的幅度, 即为“振幅”, 通常取0和1之间的值, C值越大, 生长的季节性波动越大。
采用FISAT 软件中的ELEFAN I技术估算模型生长参数[15]。首先采用Powell-Wetherall 方法估测渐近壳长L∞[16]。以此为初始值, 拟合 von Bertalanffy 季节性生长方程的生长参数L∞、K、C和“冬季点”WP。
理论生长起点年龄t0采用Pauly经验公式估算[17]:
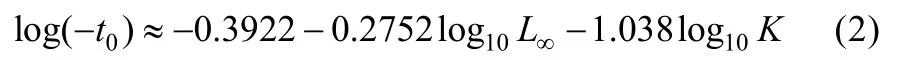
总死亡系数(Z)用 FISAT 软件中的长度变换渔获曲线法(Length-Converted Catch Curve)估算[17]。
自然死亡系数(M)采用以下公式计算[17]:
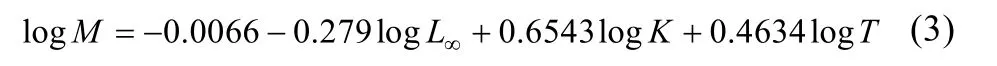
式中, T为河蚬栖息水域年平均水温。洪泽湖年平均水温取15.6℃[18]。
捕捞死亡系数(F)和开发率(E)由以下公式计算:

2 结果
共测定2390个河蚬个体, 壳长和体重范围分别为8.50—31.40 mm和0.37—11.34 g。壳长与体重呈幂函数关系(图2):

Powell-Wetherall方法估算初始L∞值为34.3 mm。Von Bertalanffy季节性生长方程生长参数的拟算值: L∞=35.80 mm, K=0.72, C=0.91, WP=0.09, Rn=0.275, t0≈–0.21a。从生长曲线图分析, 渔获物由3个世代构成; 低龄世代8、9月开始补充进入渔获物中; 生长曲线的斜率有明显的季节性波动:春季至夏季斜率逐渐升高(7月份最高), 秋季至冬季则逐渐降低(1月份最低)。低龄阶段壳长生长快(图 3)。
基于以上生长参数, 采用长度变换渔获曲线法获得总死亡系数( Z )为4.31(图4), 自然死亡系数( M )为1.05,捕捞死亡系数( F )为3.26, 开发率( E )为0.76。
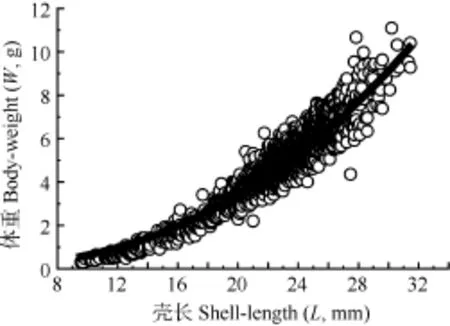
图 2 河蚬壳长(L)-体重(W)关系Fig. 2 Relationships between shell-length and body-weight of C. fluminea
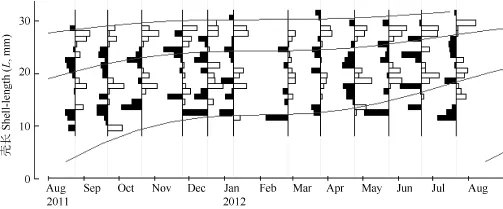
图3 河蚬壳长频数分布数据拟合的季节性生长曲线图(L∞=35.80 mm, K=0.72, C=0.91, WP=0.09, Rn=0.275)Fig. 3 Seasonalized von Bertalanffy growth curve (L∞=35.80 mm shell length, K= 0.72 per year, C=0.91, WP=0.09, Rn=0.275) of Corbicula fluminea as superimposed on the restructured shell length–frequency histogram
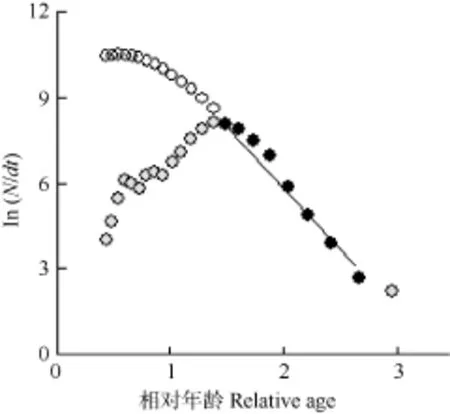
图4 变换体长渔获曲线估计河蚬总死亡系数Z=4.31Fig. 4 Estimated Z=4.31 per year base on length-coverted catch curve of Crobicula fluminea
3 讨论
洪泽湖河蚬壳长-体重符合幂函数关系, 幂指数 b为2.44。采用ELEFAN I技术分析洪泽湖河蚬壳长频率, 估算河蚬渔获物包括3个世代。壳长生长系数K为0.72, 振幅C为0.91, 冬季点WP为0.09, 种群的开发率E为0.76。
洪泽湖河蚬的繁殖季节集中, 贝壳的生长具有明显的季节性特点, 适于采用体长频率分析方法分析年龄结构[10]。但是采用体长频率分析方法估算渔获物年龄结构时, 由于高龄组体长分布常存在明显重叠, 可能低估年龄结构[8,10]; 另外, 由于捕捞的选择性, 低龄个体常不出现在渔获物中。基于本研究结果推测, 洪泽湖河蚬种群至少包括 3个年龄组, 与其他水体河蚬种群的年龄结构认识基本一致[8,9,19]。
幂指数 b值表明壳长属非匀速生长[20]。在一些河蚬入侵水域中K范围为0.070—0.65, Paraná River Delta中入侵种群振幅 C为 0.70, 相较而言, 洪泽湖河蚬生长较快,生长的季节性波动较大[8]。生长曲线分析表明春季至夏季生长率逐渐升高, 最大生长率约发生在7月, 秋季至冬季生长率逐渐降低, 最小生长率约发生在1月(冬季点)。从河蚬充分实现种群周年生长效率考虑, 河蚬捕捞在 7月之后比较恰当。本研究确定洪泽湖河蚬种群的开发率 E为0.76, 表明资源目前处于过度开发状态[21,22]。捕捞死亡系数上升是引起河蚬总死亡系数上升的主要因子, 为了降低河蚬死亡率恢复资源应降低捕捞量。
由于采用体长频率方法确定渔获物年龄结构的局限性, 需要进一步建立河蚬贝壳截面年龄分析方法来确证本研究的结果。
[1] Clavero M, Araujo R, Calzada J, et al. The first invasive bivalve in African fresh waters: invasion portrait and management options [J]. Aquatic Conservation: Marine and Freshwater Ecosystems, 2012, 22(2): 277—280
[2] Karatayev A Y, Padilla D K, Minchin D, et al. Changes in Global Economies and Trade: the Potential Spread of Exotic Freshwater Bivalves [J]. Biological Invasions, 2007, 9(2): 161—180
[3] Li D L, Zhang T, Yu J B, et al. Temporal and spatial distributional patterns of mollusca in a typical aquacultural lake—Datong Lake [J]. Acta Hydrobiologica Sinica, 2011, 35(6): 946—954 [李德亮, 张婷, 余建波, 等. 典型养殖湖泊大通湖软体动物的时空分布格局. 水生生物学报, 2011, 35(6): 946-954]
[4] Yan Y J, Li X Y. Secondary production of several dominant macrozoobenthos in Heizhuchong stream of Hanjiang River Basin [J]. Acta Hydrobiologica Sinica, 2007, 12(3): 1—306 [闫云君, 李晓宇. 汉江流域黑竹冲河部分优势大型底栖动物的周年生产量. 水生生物学报, 2007, 12(3): 1—306]
[5] Zhang C W, Zhang T L, Zhu T B, et al. Community structure of macrozoobenthos and its relationship with environmental factors in Lake Hongze [J]. Journal of Hydroecology, 2012, 33(3): 27—33 [张超文, 张堂林, 朱挺兵, 等. 洪泽湖大型底栖动物群落结构及其与环境因子的关系. 水生态学杂志, 2012, 33(3): 27—33]
[6] Zhang H C, Chen M, Fan H F, et al. Climatic background of modern Corbicula fluminea and the stable isotopes of shells from the representative areas in continental China [J]. Marine Geology & Quaternary Geology, 2007, 27(3): 77—84 [张虎才, 陈明, 樊红芳, 等. 河蚬分布的气候环境及壳体稳定同位素. 海洋地质与第四纪地质, 2007, 27(3): 77—84]
[7] Araujo R, Moreno D, Ramos M A. The asiatic clam Corbicula fluminea (Müller, 1774) (Bivalvia: Corbiculidae) in Europe [J]. American Malacological Bulletin, 1993, 10: 39—49
[8] Cataldo D, Boltovskoy D. Population dynamics of Corbicula fluminea (Bivalvia) in the Paraná River Delta (Argentina) [J]. Hydrobiologia, 1999, 380: 153—163
[9] Mouthon J. Life cycle and populations dynamics of the Asian clam Corbicula fluminea (Bivalvia: Corbiculidae) in the Saone River at Lyon (France) [J]. Hydrobiologia, 2001, 452: 109—119
[10] Sparre P, Venema S C. Introduction to Stock Assessment of Tropical Fish [M]. Beijing: Chinese Agricultural Science and Technology Press. 1992, 407 [ Sparre P, Venema S C. 热带鱼类资源评估导论. 北京:中国农业科技出版社. 1992, 407]
[11] Li Z. The study on the performances of length-frequency analysis methods on the simulated and real fishery data sets [D]. Thesis for Master of Science. Ocean University of China, Qingdao. 2006 [李壮. 渔业体长频数分析法在模拟和实际渔业中的应用. 硕士学位论文, 中国海洋大学, 青岛. 2006]
[12] Pauly D, David N. ELEFAN I, a BASIC program for the objective extraction of growth parameters from length-frequency data [J]. Berichte der Deutschen Wissenschaftlichen Kommission für Meeresforschung, 1981, 28(4): 205—211
[13] Ituarte C F. Growth dynamics in a natural population of Cor-bicula fluminea (Bivalvia, Sphaeriacea) at punta atalaya, río de la plata, Argentina [J]. Studies on Neotropical Fauna and Environment, 1985, 20(4): 217—225
[14] Pauly D. A review of the ELEFAN System for Analysis of length–frequency data in Fish and Aquatic Invertebrates [A]. In: Pauly D, Morgan G R (Eds.), Length-Based Methods in Fisheries Research [C]. Manila: ICLARM Conference Proceedings 13. 1987, 7—34
[15] Gayanilo F C, Sparre P, Pauly D. FAO-ICLARM stock assessment tools II User’s guide [M]. Rome: FAO Computerized Information Series (Fisheries), No. 8, Revised version. 2005, 168
[16] Pauly D. On improving operation and use of the ELEFAN programs, part 2, improving the estimation of L∞[J]. Fishbyte, 1986, 4(1): 18—20
[17] Pauly D. On the interrelationships between natural mortality, growth parameters, and mean environmental temperature in 175 fish stocks [J]. Journal du Conseil / Conseil Permanent International pour l'Exploration de la Mer, 1980, 39(2): 175—192
[18] Yang S J. Protection and sustainable use of biodiversity of West Lakeside zone of Hongze Lake [J]. Bulletin of Soil and Water Conservation, 2003, 23(5): 62—69 [杨士建. 洪泽湖西部湖滨的生物多样性保护与可持续利用. 水土保持通报, 2003, 23(5): 62—69]
[19] Sousa R, Antunes C, Guilhermino L. Ecology of the invasive Asian clam Corbicula fluminea (Müller, 1774) in aquatic ecosystems: an overview [J]. Annales de Limnologie-International Journal of Limnology, 2008, 44(2): 85—94
[20] Ecoutin J M, Albaret, J J, Trape S. Length–weight relationships for fish populations of a relatively undisturbed tropical estuary: the Gambia [J]. Fisheries Research, 2005, 72: 347—351
[21] Uneke B I, Nwani C D, Okogwu O, et al. Growth, mortality, recruitment and yield of Pellonula leonensis Boulenger, 1917 (Osteichthyes: Clupeidae) in a tropical flood river system [J]. Journal of Fisheries International, 2010, 5(1): 19—26
[22] Ye J Q, Xu Z L, Chen J J, et al. Resources status analysis of large yellow croaker in Guanjinyang using von Bertalanffy growth equation and fishing mortality parameters [J]. Journal of Fisheries of China, 2012, 36(2): 238—246 [叶金清,徐兆礼, 陈佳杰, 等. 基于生长和死亡参数变化的官井洋大黄鱼资源现状分析. 水产学报, 2012, 36(2): 238—246]
STUDY ON AGE STRUCTURE, GROWTH AND MORTALITY OF CORBICULA FLUMINEA IN HONGZE LAKE, CHINA, THROUGH SHELL LENGTH-FREQUENCY DATA ANALYSIS
BI Ting-Ting1,2, HOU Gang1,3, ZHANG Sheng-Yu4and XIE Song-Guang1,5
(1. The Key Laboratory of Aquatic Biodiversity and Conservation of Chinese Academy of Sciences, Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan 430072, China; 2. University of Chinese Academy of Sciences, Beijing 100049, China; 3. Fisheries College, Guangdong Ocean University, Zhanjiang 524025, China; 4. Hongze Lake Fisheries Administration Committee Office of Jiangsu Province, Huai’an 223300, China; 5. Key Laboratory of Sustainable Fisheries and Environmental Protection for Lake of Northern Jiang-su, Huai’an Research Center, Institute of Hydrobiology, Chinese Academy of Sciences, Huai’an 223002, China)
河蚬; 壳长频数; 年龄结构; 生长; 死亡
Corbicula fluminea; Shell length-frequency data; Age structure; Growth; Mortality
Q959.21
A
1000-3207(2014)04-0797-04
10.7541/2014.111
2013-02-26;
2013-11-15
淮安市重点实验室项目(HAP201204); 淮安市科技支撑项目(SN1182; SN12100)资助
毕婷婷(1989—), 女, 安徽长丰人; 硕士; 主要从事动物学研究。E-mail: bijun515@163.com
谢松光, E-mail: xiesg@ihb.ac.cn
