TMSG-1蛋白不同功能域重组质粒构建及亚细胞定位
2014-03-22,,,,
,,,,
(青岛大学医学院,山东 青岛 266071 1 病理学教研室; 2 附属医院病理科; 3 附属医院血管外科)
肿瘤转移抑制基因1(TMSG-1)是马春树等[1]于2002年应用mRNA差异显示技术,在人前列腺癌细胞系中首先克隆出来的一种肿瘤转移抑制相关基因。TMSG-1定位于染色体1q21.3,转录生成的mRNA长约2.4 kb,编码蛋白质全长为380个氨基酸,相对分子质量为45 000。TMSG-1可以抑制前列腺癌、肺癌、乳癌、肝癌等多种肿瘤细胞的侵袭及转移能力,又被称为LASS2或CerS2,LASS2主要促进长链脂酰神经酰胺C22:0和C24:0-CoA特异的神经酰胺的合成[2]。目前,对于TMSG-1的研究主要集中于肿瘤转移抑制和神经酰胺合成方面,对该基因转录表达调控的研究鲜有报道。研究结果显示,TMSG-1蛋白含有Homeodomain结构域和一个潜在的核定位信号(NLS)[3],推测TMSG-1有可能作为转录因子发挥对特定基因表达的调节作用。本文把TMSG-1分成长度不同、包含不同结构域的片段,构建不同截短体的真核表达载体,并进一步研究各个截短体的亚细胞定位,探讨TMSG-1蛋白的转录活性。
1 材料与方法
1.1 实验材料
TMSG-1全长基因并带有Flag标签蛋白的pcDNA3质粒,来自北京大学医学院分子病理室。大肠杆菌感受态细胞DH5a、pfu DNA聚合酶、2×Taq PCR Master Mix及DNA Marker,均购自北京天根公司;细菌培养用LB培养基、Agarose及氨苄青霉素钠购自Amreso公司;限制性内切酶Xho Ⅰ和EcoR Ⅰ、T4 DNA连接酶及dNTP Mixture均购自Takara公司;DNA凝胶回收试剂盒及构建质粒提取试剂盒购自Takara公司;质粒中提精提试剂盒购自Promega公司;PCR引物由上海生工公司合成;所有重组质粒由上海生工公司测序鉴定;转染试剂Lipofectamine 2000购自Invitrogen公司;Flag鼠抗人单克隆抗体购自Sigma公司。
1.2 PCR扩增各截短体基因序列
构建不同截短体(图1),分别为TLC domain(T1)、Homeodomain+TLC domain(T2)、第一个跨膜区+Homeodomain(T3)、第一个跨膜区(T4)、Homeodomain(T5)及全长TMSG-1。根据Genbank提供的TMSG-1的cDNA序列设计引物,以带Flag标签蛋白的TMSG-1全长质粒为模板,引物信息见表1。上游和下游引物5′端分别加EcoR Ⅰ和Xho Ⅰ进行酶切。扩增体系100 μL,内含:全长质粒0.5 μL,上、下游引物(10 μmol/L)各1 μL,2.5 mmol/L的dNTPs 8 μL,10×PS Buffer 10 μL及DNA聚合酶(5×106U/L)0.5 μL,最后加水至100 μL。PCR反应条件:94 ℃ 3 min;94 ℃ 30 s,55 ℃ 30 s,72 ℃ 1 min或30 s,30个循环;72 ℃恒育8 min。产物经琼脂糖凝胶电泳鉴定结果。
1.3 截短体真核表达载体的构建
将上述凝胶回收的截短体PCR产物和空质粒pcDNA3.0-flag进行EcoR Ⅰ、Xho Ⅰ充分双酶切,酶切产物再次回收目的片段。再将纯化的截短体DNA片段与线性pcDNA3-flag按照1∶4(mol/L)比例进行16 ℃过夜连接,连接产物转化感受态细胞,经氨苄青霉素挑选阳性克隆,摇菌扩大培养后菌液提取质粒,经双酶切后凝胶电泳初步鉴定,鉴定正确的克隆进一步测序鉴定。

表1 TMSG-1真核表达载体构建引物表
1.4 Western blot鉴定截短体蛋白表达
转染前培养细胞增殖至90%且状态良好,应用Promega试剂盒中量精提质粒,转染步骤[4]按照Lipofectamine 2000说明书进行,每瓶细胞转染12 μg质粒。转染4 h后更换新培养基继续培养。转染48 h后用RIPA细胞裂解液收集总蛋白,Westren blot方法[5]鉴定截短体蛋白表达。
1.5 免疫荧光观察截短体亚细胞定位
截短体重组质粒瞬时转染24 h后的Hela细胞爬片过夜,冷丙酮固定20 min,PBS漂洗后用体积分数0.01的BSA常温封闭30 min,加入抗Flag一抗37 ℃孵育2 h,PBS漂洗后加入FITC标记的二抗,PBS漂洗后DAPI染核3 min,甲醇洗8 min,体积分数0.90甘油封片。奥林巴斯BX43荧光显微镜下观察,以出现绿色荧光信号为阳性表达。
2 结 果
2.1 PCR扩增各截短体重组质粒DNA片段
TMSG-1截短体T1长761 bp,为129~380位氨基酸,包括1个TLC结构域;T2长945 bp,为71~380位氨基酸,包括Homeodomain和TLC结构域;T3长390 bp,为1~128位氨基酸,内部包含1个Homeodomain结构域;T4长198 bp,为1~70位氨基酸,仅包含了结构域前端的跨膜序列;T5长185 bp,为71~128位氨基酸,为Homeodomain结构域。见图2。
2.2 重组质粒酶切鉴定
各截短体及TMSG-1全长重组质粒经EcoR Ⅰ和Xho1双酶切均得到2条带,分别为4 500 bp的质粒条带和TMSG-1基因各截短片段。酶切前后对比结果见图3。
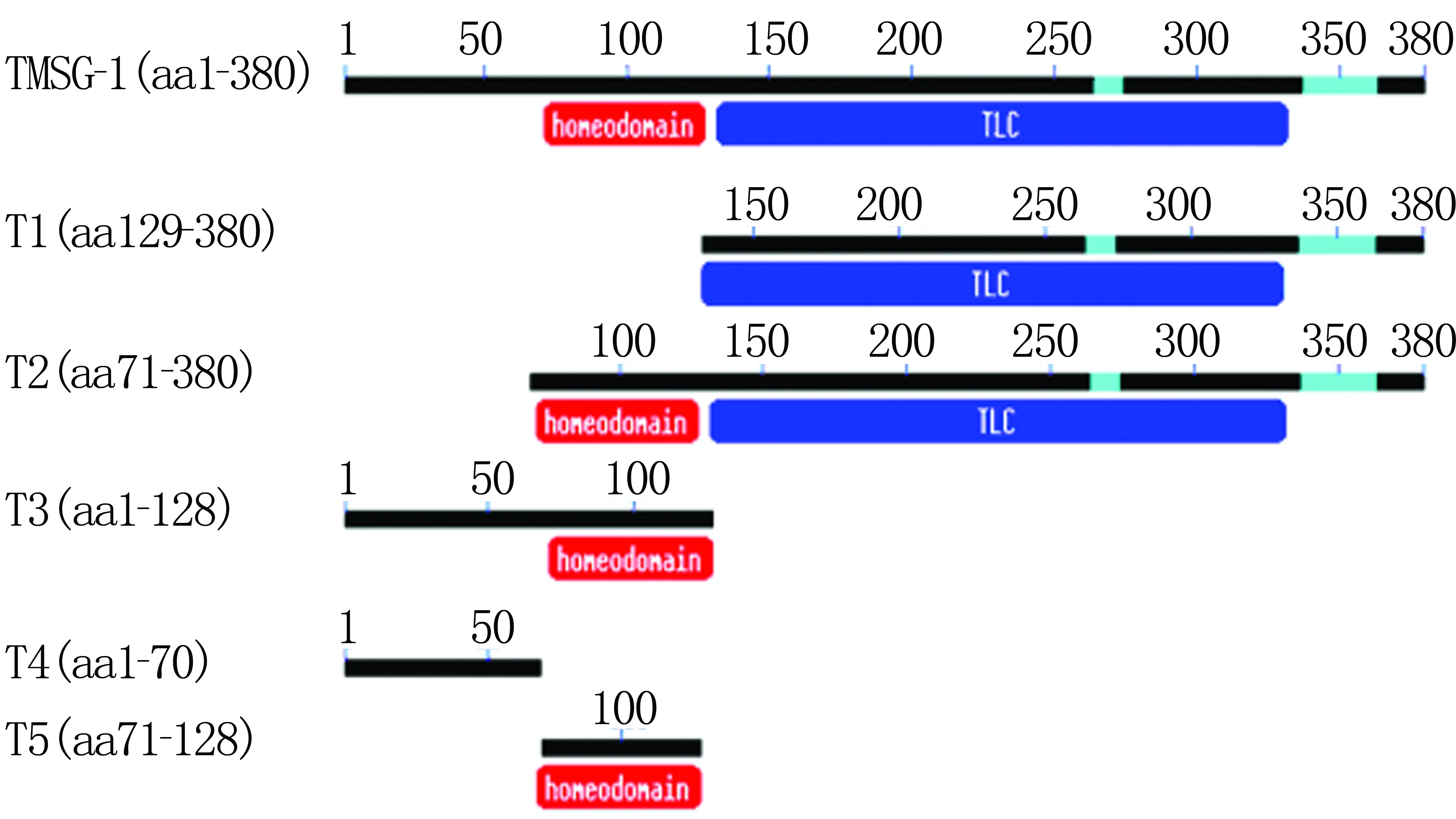
图1 TMSG-1各截短体示意图

图2 PCR扩增TMSG-1全长及各截短体基因片段
图3 各截短体及TMSG-1全长质粒酶切结果
每相邻两泳道代表酶切前后的重组质粒。酶切前仅一条大于4 500 bp的条带;酶切后可见两条带,大者为约4 500 bp的空质粒条带,小者分别对应各截短片段长度761、945、390、198、185及1 143 bp。
2.3 截短体蛋白表达
TMSG-1全长蛋白相对分子质量为45 000,根据氨基酸片段长度,预测截短体T1~T5 Flag融合蛋白相对分子质量约为2.5×104、3.4×104、1.6×104、1.0×104、1.0×104。Western blot获得蛋白条带位置与预测接近。见图4。
2.4 截短体亚细胞定位
含有Homeodomain结构域的截短体T2、T3及T5荧光信号主要位于细胞核内,细胞质内的荧光信号较弱,其中T2荧光信号在核内呈颗粒状或团块状弥漫分布,T3及T5主要呈团块状分布,类似核仁的形态,考虑为核仁定位;T1、T4、TMSG-1全长及空载体主要在胞质出现绿色荧光信号。见图5。

图4 Western blot鉴定TMSG-1全长及各截短体重组质粒蛋白表达
3 讨 论
TMSG-1是一种新的肿瘤转移抑制基因,它可以抑制多种肿瘤细胞的侵袭及转移,与肿瘤转移密切相关[6-9]。TMSG-1作为人LASS蛋白家族成员之一,其氨基端均含有1个Homeodomain结构域(LASS1除外)[10]。Homeodomain被证实是一个DNA结合功能域,含有Homeoprotein的蛋白可能与染色体DNA结合,具有转录因子活性,有转录激活或抑制作用,如Homeobox基因CDX2[11]。转录因子存在两种亚细胞定位模式,一种是细胞核分布,另一种是细胞膜或细胞质分布,在特定的信号刺激之下由细胞质进入细胞核发挥转录调节作用,核定位信号在这一转录因子调控机制中发挥了重要作用。另外,生物信息学预测也显示,TMSG-1蛋白Homeodomain结构域中的C末端119~128位氨基酸有连续4个精氨酸RRRR序列,这种结构通常被认定为核定位信号[3]。而潜在核定位信号的存在为TMSG-1有转录调节功能的设想提供有力支持。
为了明确TMSG-1蛋白是否能入细胞核,哪段区域能入核,本文初步针对其基因序列进行解体,成功构建不同长度的截短体,分别包含TMSG-1蛋白不同结构域,其中截短体T2、T3和T5均含有Homeodomain结构域,细胞免疫荧光观察显示,截短体T2、T3及T5定位于细胞核,其中T2在核内呈颗粒状弥漫分布,T3及T5则主要呈团块状定位于核仁。说明预测的Homeodomain结构域中的核定位信号发挥作用,引导截短体蛋白入核的可能性极大;T3及T5主要定位于核仁,进一步表明这一预测核定位信号极可能是核仁定位信号,类似情况以往有文献报道过[12]。
TMSG-1基因自1999年被成功克隆之后一直作为一种膜蛋白被研究,未见有其转录因子活性的报道。但是膜蛋白同样可以有转录因子活性,如甾醇调节因子结合蛋白(SREBP-1)本身是一种膜结合蛋白,在细胞中被SREBP裂解激活蛋白(SCAP)降解之后入核发挥转录因子功能[13]。已有研究结果显示,肿瘤细胞中含有Homeodomain结构域的截短体能够入核,且已知约39%的含有Homeoprotein的蛋白为转录因子,具有转录激活或者抑制作用[14]。因此,我们提出以下设想:TMSG-1蛋白有可能作为转录因子发挥作用,TMSG-1有可能和其他转录因子如NF-κB、STAT一样在肿瘤的发生发展过程中有着更为广泛的作用和更为重要的地位。基于这一目标,本文从基因结构入手,构建TMSG-1不同截短体的真核表达质粒,初步确定各截短体的亚细胞定位,为研究其转录功能打下重要基础。其确切的核定位信号及入核机制有待于进一步研究。
[参考文献]
[1]马春树,刘宇欣,郑杰,等. 应用mRNA差异显示技术克隆肿瘤转移相关基因TMSG-1[J]. 中国科学:C辑, 2002,32(3):270-275.
[2]PEWZNER-JUNG Y, BEN-DOR S, FUTERMAN A H. When do lasses (longevity assurance genes) become CerS (ceramide synthases)? Insights into the regulation of ceramide synthesis[J]. Journal of Biological Chemistry, 2006,281(35):25001-25005.
[3]龚苗子,由江峰,崔湘林,等. 肿瘤转移抑制相关基因TMSG-1核仁定位信号序列的鉴定[J]. 中华病理学杂志, 2011,40(11):767-771.
[4]王娟,高美华,张蓓. CD59与CD3在T细胞活化信号转导中的协同作用[J]. 青岛大学医学院学报, 2012,48(4):287-290.
[5]李伟伟,高美华,张蓓,等. CD59配体肽基因对卵巢癌细胞CD59表达的影响[J]. 青岛大学医学院学报, 2010,46(4):301-303,306.
[6]苏静,由江峰,王洁良,等. 人肿瘤转移抑制基因1转染对人乳腺癌细胞MDA-MB-231体外生物学行为的影响[J]. 中华病理学杂志, 2007,36(10):672-676.
[7]边巴,马春树,由江峰,等. TMSG-1基因转染对肿瘤转移表型的影响[J]. 北京大学学报:医学版, 2003,35(1):18-22.
[8]苏静,由江峰,王洁良,等. 肿瘤转移抑制基因1抑制前列腺癌细胞增殖和侵袭能力[J]. 中华肿瘤杂志, 2008,30(6):404-407.
[9]谭宁,覃文新,万晓桢,等. LASS2基因抑制HCCLM3肝癌细胞的转移[J]. 肿瘤, 2007,27(5):335-340.
[10]PEWZNER-JUNG Y, PARK H, LAVIAD E L, et al. A critical role for ceramide synthase 2 in liver homeostasis Ⅰ. Alte-rations in lipid metabolic pathways[J]. Journal of Biological Chemistry, 2010,285(14):10902-10910.
[11]POMERANTZ J L, SHARP P A. Homeodomain determinants of major groove recogntion[J]. Biochemistry, 1994,33(36):10851-10858.
[12]LIU U, DU X, KE Y. Mapping mucleolar localization sequences of 1/A6DRIM[J]. FEBS Lett, 2006,580(5):1405-1410.
[13]BROWN M S, GOLDSTEIN J L. The SREBP pathway: regulation of cholesterol metabolism by proteolysis of a membrane-bound transcription factor[J]. Cell, 1997,89(3):331-340.
[14]DENG J N, LIU S X, ZOU L Q, et al. Lipolysis response to endoplasmic reticulum stress in adipose cells[J]. J Biol Chem, 2012,287(9):6240-6249.
