湖泊生态系统稳态转换理论与驱动因子研究进展
2014-02-25赵磊刘永李玉照朱翔邹锐
赵磊,刘永,李玉照,朱翔*,邹锐
1. 云南省环境科学研究院,云南省高原湖泊流域污染过程与管理重点实验室(筹建),云南 昆明 650034;2. 北京大学环境科学与工程学院,水沙科学教育部重点实验室,北京 100871
湖泊生态系统稳态转换理论与驱动因子研究进展
赵磊1,刘永2,李玉照2,朱翔1*,邹锐1
1. 云南省环境科学研究院,云南省高原湖泊流域污染过程与管理重点实验室(筹建),云南 昆明 650034;2. 北京大学环境科学与工程学院,水沙科学教育部重点实验室,北京 100871
湖泊生态系统会在长期的人为胁迫和短期的强扰动下发生稳态转换,稳态转换前后湖泊生态系统的结构和关键过程会发生明显的变化,探求浅水湖泊稳态转换驱动因子是科学合理确定湖泊管理策略的关键所在。对湖泊生态系统稳态转换的理论内涵、驱动机制进行了总结和探讨。湖泊生态系统稳态转换的概念主要含有发生的突然性和难以预知性、系统的结构与功能发生明显变化以及存在多稳态现象等内涵;具有非线性、多阈值、多稳态,以及修复过程中的迟滞效应等特征。湖泊生态系统稳态转换的驱动因子可分为外部驱动和内部驱动两种类型。外部驱动包括外源性氮磷负荷、气候变化、风浪、湖泊水位等因子;外源性氮磷负荷和气候变化的影响具有长期性和累积性,通过逐步削弱湖泊生态系统恢复力进而引发稳态转换;风浪、湖泊水位等为突发性因子,往往表现为稳态转换的直接诱因。内部驱动包括鱼类、水生植物等因子;鱼类主要通过对水生植物、湖泊底质、浮游动物等生态组分的影响引发湖泊生态系统稳态转换;水生植物对湖泊清水稳态可能不仅存在正反馈作用,也会在一定条件下存在负反馈作用。今后应加强沉水植物生长消亡的主要环境与生物要素综合作用机理、湖泊稳态类型与主控因素等方面的研究。
湖泊生态系统;稳态转换;驱动因子
湖泊富营养化是我国当前湖泊水环境面临的突出问题。例如长江中下游的众多湖泊多数已经富营养化或正在富营养化中(秦伯强, 2002);云南省九大高原湖泊中的浅水湖泊,例如:滇池、异龙湖等,均为重富营养化。由富营养化引起了水环境功能受损、水生生态退化、蓝藻水华频繁爆发、水质性缺水日趋严重等诸多问题。为遏制日趋严重的湖泊富营养化,我国各级政府投入了大量的财力物力,自“九五”以来,国家针对重点湖(库)制订了一系列的流域水污染防治规划(舒庆, 2008),仅以滇池为例,自“九五”以来的投资已达亿元。然而,至今为止治理效果尚不明显。缺乏系统的基础理论指导是制约湖泊富营养化治理的主要问题(秦伯强, 2007)。
富营养化是湖泊生态系统稳态转换的典型现象(Scheffer, 1993),湖泊富营养化过程可以归结为湖泊生态系统在长期的人为胁迫和短期的强扰动下由清水稳态转换为浊水稳态,生态系统结构与功能发生了根本性变化。以生态系统稳态转换理论指导富营养化湖泊治理与生态修复是解决当前水体富营养化的根本途径(刘永定等, 2007;秦伯强, 2007)。理解湖泊生态系统稳态转换的首要问题是明确稳态转换发生的胁迫因素和驱动机制。本文阐述了生态系统稳态转换的基本理论和核心内涵,总结归纳了湖泊生态系统稳态转换的驱动因子,以期为湖泊富营养化修复提供有益参考。
1 稳态转换概念与内涵
1.1 稳态转换概念
Holling对生态恢复力的研究,以及May阐述的生态系统多稳态变化和阈值概念初步奠定了生态学界对稳态转换的一致认知:持续的外来胁迫会降低生态系统的恢复力,从而使其超过阈值的范围并发生稳态变化(Holling, 1973; May, 1977)。Carpenter定义湖泊发生稳态转换是指生态系统的结构与动力学过程在短时间内发生突然变化,并且转化后的稳态能够持久保持(Carpenter, 2003);Collie在海洋生态系统稳态转换的研究中,将其定义为“海洋空间发生的低频率、高振幅的变动,并且该变动会在海洋的一些营养水平变动中得以传播,因而对生物量产生的影响尤为显著”(Collie等,2004);Cury提出“稳态转换是指海洋生态系统的结构和功能发生突然性改变,这些改变影响了一些生物的生存进而导致了一个稳定状态的生成”(Cury和Shannon,2004)。
尽管表述有所不同,但研究者对生态系统稳态转换内涵的认知基本一致,主要为:1)稳态转换是在短时间内发生的,具有突然性和难以预知性;2)系统的结构与功能发生明显变化,存在多稳态现象;3)转换后生成另外一种稳态,并且该稳态能够长期保持具有稳定性。
1.2 稳态转换类型
影响生态系统的因素,例如气候变化、土地利用变化、资源过度开发、污染物排放、生物多样性丧失等,大多是随时间逐渐变化的,具有线性特征(Vitousek, 1997)。然而,由于生态系统固有的复杂性、非线性、以及多阈值效应的特征,生态系统对环境因素的响应具有不同的形式。生态系统从一种稳定状态转变成另外一种稳定状态的过程可能是连续的渐变,也可能是不连续的突变或者其他变化形式。研究者多用抽象的数据模型描述多种生态系统稳态转换类型,其中最具代表性和影响力的是Scheffer提出的模型(Scheffer等,2001)。Scheffer在总结多种类型生态系统对环境变化响应形式的基础上,将稳态转换分为三种类型(图1):平滑型(a)、突变型(b)和不连续变化型(c)。
Scheffer等(2001)对三种稳态转换类型做了详细阐述。平滑型稳态转换是生态系统与驱动因素之间的响应基本呈线性关系,生态系统状态随着环境要素的变化而直接变化。突变型稳态转换中生态系统存在明显的阈值效应,驱动因素在逐渐的变化过程中对生态系统恢复力(Ecological resilience)产生损害,生态系统对干扰的承受能力逐渐降低,当驱动因素超过阈值时生态系统将会发生突然和本质的变化。与以上两种类型不同,当生态系统对驱动因素的响应为不连续型稳态转换时,即使环境要素处于同一水平,生态系统可能存在明显的截然不同的稳态(alternative stable state)。图1(c)描述的模型涵盖了稳态转换的3个基本内涵:多稳态(Alternative stable state),阈值(Threshold)和迟滞效应(hyperthsis)。图中上部实线与下部实线分别代表一种稳态,中部的虚线代表一种亚稳态。生态系统处于上部实线所代表的稳态时,当环境要素接近F2(阈值)时,环境要素微小的变化将促使生态系统发生突变,转变为下部实线所代表的稳态;如果生态系统要由下部实线代表的稳态恢复至上部实线代表的稳态,环境要素需要达到比灾变点F2小的多的F1,这是明显的迟滞效应。

图1 生态系统稳态转换的三种类型(改自Scheffer等, 2001)Fig. 1 Three types of ecosystem regime shifts (Modified from Scheffer et al, 2001)
1.3 稳态转换的内涵
1.3.1 多稳态(Alternative stable state)
多稳态是指在相同的外界环境条件下,有些生态系统有可能出现两种或多种结构与功能完全不同的稳定状态(Scheffer, 1993)。多稳态现象是生态系统发生稳态转换的基础,正是因为生态系统存在多稳态现象,生态系统才有可能发生稳态转换。在多稳态的概念中,外界环境条件可能是一种主导条件,也可能是多种环境条件,例如:捕食压力、营养盐负荷、森林大火等。在湖泊生态系统中,外界环境条件主要是外源性营养盐负荷、湖体磷含量等。结构不同指的是系统的结构发生较大变化,例如研究者一致认识到的浅水湖泊存在两种结构明显不同的稳定状态,一种是以大型水生植物为主导的清水稳态,一种是以藻类为主的浊水稳态(Bayley和Prather,2003);而功能不同则指的是随着结构的改变,系统相应的物质流、能量流以及信息流功能发生变化,结果也会造成生态系统服务价值的变化(冯剑丰等,2009)。
研究者多采用“杯中弹子模型”来描述生态系统的多稳态现象(Scheffer, 1990)。弹子代表生态系统所处的状态,杯子由底部和倾斜的斜面组成,当弹子处于杯子底部时表明生态系统处于平衡状态,杯子的斜面是一种不稳定平衡状态,称为吸引域(basins of attraction)。随着外界条件的逐渐变化时,杯子的形状会发生变化,这同时改变了稳定性(恢复时间)和弹性(决定稳定性的宽度)。当杯子中的谷峰消失时,弹子发生突然的跃迁。
1.3.2 生态系统恢复力(Ecological resilience)
Holling总结了恢复力的特征并指出,恢复力是动态系统远离平衡状态的行为,将生态系统恢复力定义为:系统在转变为一个具有不同的结构和功能的稳态之前所能够承受的扰动大小(Holling, 1973)。Folke对生态恢复力的定义为:系统承受扰动的能力,以及遭受变化时保持核心的功能、结构、特征和反馈机制稳定的自组织能力(Folke等,2004)。由于生态系统存在多稳态现象,并且生态系统始终处于动态变化之中,生态系统恢复力具备以下特征:生态恢复力是生态系统某一稳态对干扰的承受能力;生态恢复力并不是一个定值,相反具有明显的动态性,其大小会随着系统外界条件的变化而变化;生态系统恢复力具有难预测性。
1.3.3 阈值(Threshold)
1977年May在Nature上发表了《Thresholds and breakpoints in ecosystems with a multiplicity of stable states》,阐述了生态系统的多稳态变化和阈值概念,并指出:生态系统的恢复力可以衡量系统在受到外界胁迫时的承载容量,而阈值则反映生态系统可能发生状态变化的临界点(May, 1977),其影响延续至今,成为稳态转换的基础。综合人们的研究,阈值的定义为:在一些生态系统中,即使是环境状态的微小变化从而突破了关键的阈值都可能导致系统的剧烈转换,该关键阈值被称为灾变点(May, 1977;Scheffer等,2003;Kuznetsov, 1995)。
就湖泊而言,能够引起湖泊稳态转换的因素较多,例如:外部驱动的氮磷营养盐、气候变化、湖泊水位等,内部驱动的鱼类、水生植物等,因此,湖泊稳态转换的阈值是多维度的,可能是随时间而缓慢变化的环境因子,也可能是湖泊内部的生物因子(Carpenter和Lathrop,2008),并且这些因子之间存在复杂的相互作用。我们不能脱离湖泊特定的生态条件而确定具有统一性的阈值。
1.3.4 迟滞效应(hysteresis)
减少营养盐负荷,包括同时削减氮和磷并不一定能取得理想的结果。由于历史原因造成的“遗留负荷”的影响,系统可能无法按照预想轨迹恢复到未受损害时的状态而进入另一种状态或已改变了的基准状态,这种现象即为迟滞效应。在湖泊富营养化治理中,将湖体营养盐含量降低至受污染前的水平往往并不能抑制藻类爆发,使湖泊处于清水稳态。例如:荷兰Veluwe湖受污染前TP质量浓度约0.15 mg·L-1,经过多年的治理,湖体TP水平降至约0.1 mg·L-1时湖泊仍处于浊水稳态(Scheffer和Carpenter, 2003)。导致富营养化浅水湖泊迟滞效应的主要原因是由于受损湖泊在稳态转换前后生态系统的内部结构、驱动因素和关键过程均发生了显著变化,生态系统具有明显的不可逆性。
2 湖泊生态系统稳态转换驱动因素
2.1 外部驱动
外部驱动力能够在较大时间、空间尺度上引起生态系统的缓慢变化,并导致系统稳态转换。现有的研究表明,导致湖泊生态系统发生稳态转换的主要外部驱动力包括外源性营养盐输入、气候变化、水文等要素。
2.1.1 营养盐输入
淡水湖泊生态系统富营养化导致了许多湖泊水生植物消亡或覆盖度下降,使得许多原先以水生植物为主的草型生态系统转换为以浮游植物为主的藻型湖泊生态系统(Scheffer和Van Nes, 2007),是一种典型的生态系统稳态转换(Scheffer等, 1993),也是在湖泊研究和治理中的热点和难点。外源性营养盐输入是导致湖泊富营养化的主要原因,水生植物生长过程中需要氮来合成蛋白质,需要磷来组成DNA、RNA和传输能量,这两种物质是水生和陆地生态系统中主要的限制性营养盐(Conley等,2009)。营养盐胁迫导致的湖泊生态系统稳态转换在多个湖泊得到了证实(陈开宁等, 2006;Ibelings等,2007;Gonzales等, 2005)。荷兰Veluwe湖的长期观测数据表明,随着20世纪60年代至70年代TP逐步增加,当TP质量浓度增加到0.15 mg·L-1后,湖泊中水生植物覆盖度在减少,水生植物完全消亡时湖体的TP质量浓度高于0.20 mg·L-1(Ibelings等, 2007)。通过采取削减营养盐负荷,Veluwe湖逐步恢复至清水状态,这期间湖泊经历了浊水稳态、浊水与清水的过渡状态、清水稳态。丹麦204个浅水湖泊(水深小于5 m,面积大于5 hm2)的湖泊夏季总磷、总氮浓度与水生植物覆盖度的调查表明:当总磷浓度达到0.1 mg·L-1以上,总氮浓度达到2 mg·L-1以上时,水生植物的覆盖度几乎趋于零(Gonzales等, 2005)。
尽管已有许多关于氮磷引起的湖泊藻类爆发、水生植物消亡的观测实证、模型模拟等方面的研究,但仍有几个问题仍然存在争议或者模糊不清(秦伯强, 2007; Conley等, 2009; Scheffer和Van Nes, 2007)。一是关于氮磷在湖泊富营养化过程究竟那种物质是主导的限制性因子或者是两者均需要得到控制(Conley等, 2009);二是氮磷胁迫下湖泊富营养化高等水生植物大范围消亡的机制;三是保证湖泊处于稳定的清水状态时水体中的氮磷含量需要降低到什么样的水平(灾变阈值与恢复阈值)(Scheffer和Van Nes, 2007)。
2.1.1.1 主导限制性因子
1977年Schindler在Scinece上发表了《Evolution of phosphorus limitation in lakes》,该文根据加拿大湖区227号湖长期的大规模实验结果,指出磷是湖泊富营养化主要的限制因子(Schindler, 1977)。此后,“削减磷负荷”成为北美和欧洲进行湖泊管理的主要策略,许多湖泊通过削减磷负荷改善了水质(Marsden, 1989; Jeppesen, 2007)。理论上来讲,当湖泊中富含磷而缺氮时,由于氮可以通过固定大气中的气态氮得到补充,导致可能出现具有固氮功能的蓝藻爆发,可以缓解湖泊中氮紧缺的问题,使湖泊仍处于磷限制的状态(Karl等, 2003)。Schindler在加拿大湖区227号湖的氮磷操控实验进一步证实了以上理论,在保持磷入湖量不变而持续削减氮入湖量的情况下,伴随着固氮蓝藻大量生长湖泊仍然处于富营养化状态(Schindler等, 2008)。Jeppesen在丹麦35个湖泊的调查指出,通过削减磷负荷,湖泊TP含量降低,进而湖泊中的浮游植物结构发生了变化,浅水湖泊中的硅藻、隐芽植物、金藻的生物量明显增加(Jeppesen等, 2005)。然而,最近的研究指出,氮对湖泊生态系统的作用可能被低估了。Gonzalez对丹麦浅水湖泊的调查发现,只有当夏季湖泊水体的TN质量浓度低于2 mg·L-1时大型水生植物的覆盖度才会比较高,而与TP浓度没有直接的关系,同时期TP的质量浓度范围为0.03~1.2 mg·L-1(Gonzalez等, 2005)。美国Apopka湖是一个典型的氮限制湖泊。Aldirdge针对Apopka湖的营养添加生物实验结果证明,在实验中添加磷不能刺激浮游植物生长,但浮游植物对添加氮有显著反应,并且湖泊中爆发的蓝藻并非固氮藻(Aldridge等, 1993)。而单纯的磷负荷控制策略也没有很好的改善Apopka湖的水质(Bachmann等, 1999)。针对这种争议,Conley等(2009a)、Schindler和Hechy(2009)在Sciences上分别发表了《Controlling eutrophication: nitrogen and phosphorus》和《Eutrophication: more nitrogen data needed》。Conley认为蓝藻可以通过固氮得到足够的氮源以支撑水体中利用磷源生长的藻类繁殖的观点并不总能在湖泊中得到证明,在一些湖泊中,底泥和水体中的磷循环迅速,浮游植物主要为不具有固氮功能的蓝藻,解决水陆系统中的富营养化问题需要合理的同时控制氮磷两种污染物;Schindler认为许多湖泊依靠仅控制磷的手段取得了成果,但还没有哪个水生系统的研究证实,减少氮的输入可以解决富营养化问题,我们还需要更多的有关氮的数据。显然,关于氮磷在湖泊富营养化过程究竟那种物质是主导的限制性因子或者是两者均需要得到控制的问题,是事关湖泊富营养化控制策略的基本问题,目前科学家尚未达成一致意见,也许我们还需要更多的实验、模型去回答这个问题。
2.1.1.2 氮磷胁迫下水生植物消亡机制
氮磷营养盐胁下水生植物消亡湖泊由清水稳态转换为浊水稳态的机制,是通过营养盐胁迫下湖体浮游植物、附着植物、浮游动物、沉积物、鱼类以及水生植物相互之间的正反馈作用实现的(图2)(Scheffer等, 2007; Zimmer, 2007; Anton等, 1989)。

图2 水生植物与富营养化关系的描述性模型(改自Anton Hough R, 1989)Fig. 2 A general descriptive model of aquatic plant relationships to eutrophication (Modified from Anton Hough R, 1989)
水生植物消亡的第一步是氮磷营养盐过量输入至湖泊导致藻类、附着植物大量生长,这个过程将降低湖泊的透光性(Bachmann, 2002),显著降低水生植物的光合作用(秦伯强等, 2006),降低水生植物的生物量、结构组成以及覆盖度。但是,这一过程并不能导致水生植物的完全消亡。就透光性而言,一方面在水生植物存在的情况下,藻类、附着植物的生物量难以大量生长(秦伯强等, 2006; Jones和Sayer, 2003);另一方面藻类对湖泊透光性的影响相对较小,并不足以将湖泊的透光性降低至水生植物完全无法生长的条件,例如太湖的研究发现,光的衰减主要受悬浮物和叶绿素a影响,其中起主导因素的悬浮物,浮游植物对光的衰减能够在一定程度上降低真光层深度,但水生植物依然能够生长(张运林等, 2006)。
随着水生植物覆盖度、生物量减少,水生植物对维系湖泊清水状态的正反馈作用减弱,浮游动物数量减少、体型小型化,附着植物生物量增加,底泥再悬浮及营养盐释放作用增加。浮游植物生物量和底泥再悬浮增加使得湖泊的透明度进一步减弱、浊度增加、透光性减弱,而水生植物在与藻类的光竞争中处于劣势(Bilotta和Brazier, 2008),水生植物的生长收到明显的光限制,这种情况下湖泊生态系统处于不稳定状态。
氮磷负荷增加或者减少能够使得湖泊发生稳态转换,然而生态系统稳态转换理论与大量的实例告诉我们,在一定的湖泊氮磷含量下,湖泊存在多个稳态,湖泊生态系统稳态存在一定的随机性,单纯的氮磷负荷增加或者减少并不一定能够导致湖泊生态系统稳态转换(Scheffer和Van Nes, 2007),一些突发性干扰也许在稳态转换中起着非常关键的作用(Scheffer和Van Nes, 2007),例如:风浪、湖泊水位升高、极端气候、大量草食性鱼类放养等,都可能使得湖泊水生植物大量消亡或者生长而发生稳态转换。实际上这也是湖泊生态修复的理论基础,即单纯的负荷削减并不能完全恢复良好湖泊生态系统,需要通过生态修复的协同手段,包括鱼类控制、水生植物修复,才能将湖泊由浊水稳态转换为清水稳态(秦伯强, 2007)。丹麦多个湖泊的长期观测表明单纯的负荷削减情况下湖泊的水生植物基本保持稳定,而同时进行负荷削减和生物操纵的湖泊水生植物具有明显增加(Søndergaard等, 1990;Perrow等, 1994;Jeppesen等, 2005)。
综合目前的研究,氮磷营养盐是湖泊生态系统稳态转换的必要先决条件,但不一定是充分必要条件,其所起的最主要作用是削弱湖泊生态系统的恢复力,为突发性诱因奠定了基础,进而引发了一系列湖泊生态系统的连锁反应,引起了湖泊光照、底泥环境、浮游植物、浮游动物、附着生物等生物与非生物因子结构组成上的变化,并最终导致了湖泊生态系统稳态转换的发生。
2.1.1.3 氮磷阈值
尽管针对氮磷在湖泊生态系统稳态转换中的具体机制仍然存在一些争议,但达成共识的是,氮磷是引起稳态转换的最重要因素,通过限制水体中营养物的输入,是控制富营养化及其负面影响最重要、最敏感,同时也是最可行的做法(Conley等,2009b)。对湖泊而言,主要存在两个氮磷阈值,一是由清水稳态转换为浊水稳态的灾变阈值,二是由浊水稳态转换为清水稳态的恢复阈值。灾变阈值是对湖泊生态系统作出预警的重要依据,恢复阈值是制定湖泊管理与恢复策略的重要基准。在针对氮磷阈值的研究中,研究者采用实验观测、统计分析、模型模拟等多种方法研究了湖泊的氮磷含量(负荷)阈值,取得了丰富的成果,例如:Solheim等(2008)针对欧洲1000个湖泊的调查发现,蓝藻与TP之间以及水生植物与TP之间均存在阈值现象,当湖泊TP高于12~20 µg·L-1时,湖体的蓝藻数量会大幅度增加,当湖泊TP高于20~40 µg·L-1时,水生植物会明显减少。Zimmer通过对美国72个浅水湖泊的调查指出,湖体TP质量浓度小于0.62 mg·L-1时湖泊能够保持清水状态(Zimmer等, 2009);经历了富营养化、水生植物消亡和逐渐恢复的Veluwe湖,其水生植物消亡于的湖体TP质量浓度高于0.20 mg·L-1,而大型水生植物恢复时湖体TP质量浓度小于0.10 mg·L-1(Ibelings等, 2007)。
2.1.2 气候变化
已有研究发现,气候变化会对湖泊的物理过程、地球生化循环以及生态系统的结构和功能产生直接的影响(Clair, 1998; Blenckner等, 2002; Carvalho和Kirika, 2003; Komatsu等, 2007);如在对美国五大湖的研究中,研究人员发现,气候变化会对五大湖的水位、流域污染物输入、水温、冰层面积、溶解氧、水温分层、生产力、生态系统结构、生物栖息地以及生态系统脆弱性等方面造成明显的影响,从而直接关系到美、加两国在五大湖区域的管理措施与对策(Chao等, 1999; Great Lakes Water Quality Board, 2003)。以湖泊水温升高为例,将带来水动力学过程的改变,加长湖泊分层的时间,提高湖泊沉积层中营养物质的释放速率和释放量以及浮游植物的生长速率,从而加剧湖泊的富营养化(Blenckner等, 2002;Malmaeusa等, 2006)。
IPCC 2008在发布的技术报告之六(Climate Change and Water)中,重点评估了气候变化所可能导致的降水强度和频率的增加,并且由此可能对影响水质并使许多水污染形式加重。以湖泊为例,这种影响机制主要表现在两方面:1)对流域地表过程和营养物质输入的影响;2)对湖泊水动力学过程和生态系统的直接影响(Bates等, 2008)。流域的水文过程对气候因素的影响十分敏感,观察试验表明,流域的降水强度和频率的增加会导致洪水和短时暴雨径流的产生,从而对地表产生短时的剧烈冲刷,增加水土流失和其他非点源污染输入的量,尤其是可溶污染物和被土壤颗粒吸附的污染物(Choi, 1998;Groisman等, 2001;Bouraoui等, 2004)。短时间大量输入的污染物量会对水质和生态系统造成严重的冲击,增加营养物质和沉积物的含量、降低水体透明度;对处于生态恢复进程中的湖泊而言,则有可能会造成生态系统逆转的严重后果(Malmaeusa等, 2006)。同时,降水强度和频率的增加还会造成一年内其他时段可能出现严重的干旱、低流量期延长,从而减少进入湖泊的地表径流,降低水体对污染物的稀释能力(Najjar, 1999; Kumagai等, 2003)。除了作用于流域的基础地表过程从而对湖泊产生间接的影响外,气候变化还通过对水温和水动力学过程等对湖泊产生直接的影响。气候变化将可能导致带来湖泊水温的升高和热稳定性的增加,从而降低水体中的含氧量,增加沉积物的磷(P)释放量。以安大略湖为例,研究证实,水温升高3~4 ℃将会带来1倍的P释放量增加(Nicholls, 1999)。在生态系统稳态转换中,气候变化也许将会缓慢但将导致生态系统发生不可逆的灾变。
2.1.3 其他因素
氮磷营养盐、气候变化均是缓慢变化的外界胁迫因素,能够从根本上改变湖泊生态系统恢复力,为湖泊生态系统稳态转换创造必要条件,而一些突发因素,诸如风浪、湖泊水位等因素将能够引起湖泊生态系统稳态转换。
风浪造成湖泊生态系统稳态转换的机制主要是大风暴对水生植物的损害,强烈的风浪将会使水生植物根茎折断,严重的甚至会连跟拔起(秦伯强, 2007)。典型的例子是美国的Appoka湖,该湖在1947年之前一直是清水草型湖泊,水生植物覆盖度达80%,1947年9月的一次强烈的风暴过程使得湖泊的水生植物大量消亡,1个星期后该湖首次出现蓝藻水华,之后一直处于浊水稳态(Bachmann等, 1999)。
湖泊水位能够通过影响湖体的光照条件进而导致湖泊生态系统稳态转换。连续的高水位会导致浅水湖泊中适宜于水生植物生长的光照条件减弱,面积减小,进而在一定时期内导致水生植物覆盖度降低。例如:固城湖在20世纪90年代连续2年的高水位导致水生植物大范围的死亡,同时也使得湖泊水质下降(谷孝鸿等, 2002)。瑞士的Tämnaren湖由于1977年的高水位使得湖体的水生植物覆盖度由1973年的80%降低到1983年的14%(Wallsten和Forsgen, 1989)。美国Okeechobee湖28年的观测资料和操控实验显示,沉水植物生物量与高水位呈负相关,连续稳定的高水位将延缓水生植被的恢复(Havens等, 2001)。
需要指出的是:风浪、湖泊水位等突发性因素导致湖泊生态系统稳态转换的前提条件是湖泊生态系统已经处于不稳定的状态,虽然从表面上看突发性因素引起了稳态转换,但最根本的诱因仍然是氮磷营养盐的过量输入。
2.2 内部驱动
已有的研究发现生态系统内部驱动力会对生态系统的营养链级、营养物质循环产生直接的影响从而引起稳态转换。
2.2.1 鱼类
在内部驱动力中营养链级变化引起的稳态转换得到广泛关注(Lammens等, 2004)。鱼类,包括草食性鱼类(herbivorous fish)、浮游生物食性鱼类(Planktivorous fish)、底栖性鱼类(Benthivorous fish)和食鱼性鱼类(Piscivorous fish),通过对水生植物、湖泊底质、浮游动物的复杂的正反馈作用在湖泊稳态转换过程中关键作用(图3)。草食性鱼类通过对沉水植物的捕食直接影响湖泊沉水植物的生长,增加湖泊沉积物悬浮、营养盐释放,促进藻类生长(Gideon和William,2010;刘永等,2012)。底栖类鱼类会对湖泊沉积物产生扰动从而增加沉积物再悬浮和营养盐释放,减少湖体透明度,抑制沉水植物生长,例如:Eddy在Veluwe湖的研究发现商业性捕鱼导致底栖类鱼类的减少了约35%之后,Veluwe湖的沉水植物开始生长、湖体透明度增加(Lammens等, 2004)。鱼类主要通过对浮游生物种群密度的作用影响湖泊生态系统,研究发现随着浮游生物食性鱼类对浮游动物捕食量的减少,浮游动物生物量增加会使得藻类生物量减少。食鱼性鱼类作为湖泊的顶级捕食者会对其它鱼类产生影响,进而改变湖泊的营养链级。Carpenter等发现,Michigan湖、Tuesday湖和Peter湖中放养食鱼性鱼类后,滤食性鱼类数量显著减少而浮游动物大量繁殖,从而导致了Chla含量和初级生产力显著降低(Carpenter, 1988)。
2.2.2 水生植物
大型水生植物能够通过多种机制维系湖泊清水状态,包括为浮游动物提供庇护、增加沉积物的稳定性从而减少沉积物再悬浮、与浮游植物竞争营养及释放克藻物质等(Roger, 1999; Ellen, 1993; Van Nes, 2002)。然而,水生植物似乎也会通过对湖泊沉积物、营养物质滞留的负反馈作用导致生态系统稳态转换。历史数据显示英国的Alderfen Broad湖、荷兰Botshol湖约每7~8年会发生周期性的清水-草型和浊水-藻型的生态系统稳态转换(Martin等, 1994; Winnie等, 2005),然而并没有外部驱动力能够导致该现象。为揭示该周期性稳态转换的驱动力,Van Nes等(2007)分别建立了水生植物与湖体磷滞留关系模型、水生植物与厌氧状态沉积物磷释放模型,结果显示:两种模型都能够模拟湖体的周期性稳态转换,相对而言水生植物与厌氧状态沉积物磷释放模型的模拟结果更接近于实际情况。这种由于水生植物对湖体营养状态负反馈导致的稳态转换机制被称作“时间炸弹(time bomb)”(Van Nes, 2007)。水生植物的死亡后以有机质大量累积与湖泊底部,有机质的耗氧分解使得水-沉积物界面处于厌氧状态促使沉积物磷过量释放,而磷的释放促进了藻类生长并导致水生植物生长所需的透明度降低,最终致使水生植物消亡,湖泊由清水-草型转变为浊水-藻型。如果浊水-藻型状态下湖泊底质有机质能够大量逐步分解损失并产生好氧状态,沉积物磷释放量减少,湖泊又会由浊水-藻型转变为清水-草型(Scheffer和Van Nes, 2007)。上述发现告诉我们,水生植物对湖泊清水稳态不仅仅是通常认识中的正反馈作用,也存在负反馈作用,而沉积物在其中起着关键作用。然而,这种负反馈作用固然可能是湖泊生态系统中的普遍现象,但只能在一定条件下才会导致湖泊发生生态系统稳态转换。首先湖泊应处于不稳定的状态,其次水生植物要能够对湖泊有足够的影响力(Scheffer和Van Nes, 2007)。要满足上述条件,湖泊应该面积较小并且污染较为严重。
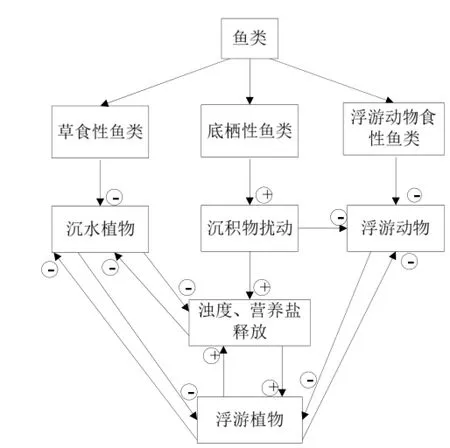
图3 鱼类在湖泊生态系统稳态转换中的正反馈作用Fig. 3 Positive feedback of fish in lake ecosystem regime shifts
3 研究展望
本文系统总结了湖泊生态系统稳态转换理论与驱动因子,湖泊生态系统稳态转换与外部环境因子和内部生物因子密切相关,并且这些因子存在复杂的相互反馈作用;此外,由于不同湖泊所处的演化进程、营养状态、物理形态等大相径庭,导致稳态转换的驱动因子存在极大的差异性。因此,湖泊稳态转换驱动因子具有明显的复杂性,尽管目前相关研究已取得了较多的成功,但仍存在较多的问题需要进一步的深化研究,概括起来主要包括3个方面:
1)影响沉水植物生长消亡的主要环境与生物要素机理研究。沉水植物生长消亡是浅水湖泊稳态转换的关键表征,沉水植物生长消亡受到光照、营养盐、沉积物、温度、藻类等多种环境与生物要素的影响,并且各种要素之间存在着相互作用与联系,应考虑多重因子之间的相互综合作用,科学地反映反映沉水植物生长消亡过程与环境及生物要素之间的相互关系。
2)湖泊生态系统有着丰富的多样性,对于处于富营养化状态的湖泊而言,由于特征营养元素、所处纬度、深度、流域特征等导致稳态转换过程的驱动因子和机理有所区别。应加强大样本的比较湖沼学研究,寻求不同类型湖泊所处的稳态,研究不同稳态的主控因素。
3)应用多种技术手段还原反演湖泊生态系统演化进程。
ALDRIDGE F J, SCHELSKE C L, CARRICK H J. 1993. Nutient limitation in a hypereutrophic Floride lake[J]. Archives of Hydrobiology, 127:21-37.
ANTON Hough R, FORNWALL M D, NEGELE B J, et al. 1989. Plant community dynamics in a chain of lakes: principal factors in the decline of rooted with eutrophication[J]. Hydrobiologia, 173:199-217.
BACHMANN R W, HORSBURGH C A, HOYER M V, et al. 2002. Relations betweent trophic state indicators and plant biomas in Florida lakes[J]. Hydrobiologia, 470:219-234.
BACHMANN R W, HOYER M V, CANFIELD D E. 1999. The restoration of Lake Apopka in relation to alternative stable states[J]. Hydrobiologia, 394:219-232.
BATES B C, KUNDZEWICZ Z W, WU S, et al. 2008. Climate Change and Water[R]. Technical Paper of the Intergovernmental Panel on Climate Change, IPCC Secretariat, Geneva, 210.
BAYLEY S E, PRATHER C M. 2003. DO wetland lakes exhibit alternative stable states? Submersed aquatic vegetation and chlorophy11 in western boreal shallow lakes[J]. Limnology and Oceanography, 48(6): 2335-2345.
BILOTTA G S, BRAZIER R E. 2008. Understanding the influence of suspended solids on water quality and aquatic[j]. Water Research, 42:2849-2861.
BLENCKNER T, OMSTEDT A, RUMMUKAINEN M. 2002. A Swedish case study of contemporary and possible future consequences of climate change on lake function[J]. Aquatic Science, 64:1-14.
BOLL T, JOHANSSON L S, LAURIDSEN T L, et al. 2012. Changes in benthic macroinvertebrate abundance and lake isotope(C, N) signals following biomanipulation: an 18-year study in shallow Lake Vaeng, Denmark[J]. Hydrobiologia, 686:135-145.
BOURAOUI F, GRIZZETTI B, GRANLUND K, et al. 2004. Impact of climate change on the water cycle and nutrient losses in a Finnish catchment[J]. Climatic Change, 66:109-126.
CARPENTER S R, BROCK W A. 2006. Rising variance: a leading indicator of ecological transition[J]. Ecology Letters, 9: 311-318.
CARPENTER S R, COLE J J, PACE, M L et al. 2011. Early warnings of regime shifts: a wholeecosystem experiment[J]. Science, 332:1079-1082.
CARPENTER S R, LATHROP R C. 2008. Probabilistic estimate of a threshold for eutrophication[J]. Ecosystems, 11: 601-613.
CARPENTER S R. 1988. Complex interactions in Lake communities[M]. Speringer-Verlag, New York, 283. 缺项
CARPENTER S R. 2003. Regime Shifts in Lake Ecosystems: Pattern and Variation[M]. Volume 15 in the Excellence in Ecology Series. Ecology Institute, Oldendorf/Luhe, Germany.
CARPENTER S R. 2005. Eutrophication of aquatic cosystems: bistability and soil phosphorus[J]. Proc Natl Acad Sci USA, 102: 10002-10005. CARVALHO L, KIRIKA A. 2003. Changes in shallow lake functioning: response to climate change and nutrient reduction[J]. Hydrobiologia, 506-509, 789-796.
CHAO P T, HOBBS B F, VENKATESH B N. 1999. How climate uncertainty should be included in Great Lakes management: Modeling workshop results[J]. Journal of The American Water Resources Association, 35(6): 1485-1497.
CHOI J S. 1998. Lake ecosystem responses to rapid climate change[J].Environmental Monitoring and Assessment, 49: 281-290.
CLAIR T A. 1998. Canadian freshwater wetlands and climate change: guest editorial[J]. Climate Change, 20: 163-165.
COLLIE J S, RICHARDSON K, STEELE J H. 2004. Regime shifts: can ecological theory illuminate the mechanisms[J].? Progress in Oceanography, 60(2/4): 281-302.
CONLEY D J, PAERL H W, HOWARTH R W, et al. 2009a. Controlling eutrophication: nitrogen and phosphorus[J]. Science, 323:1014-1015.
CONLEY D J, PAERL H W, HOWARTH R W, et al. 2009b. Eutrophication: Time to adjust expectations response[J]. Science, 324:724-725.
CURY P, SHANNON L. 2004. Regime shifts in upwelling ecosystems: observed changes and possible mechanisms in the northern and southern Benguela[J]. Progress in Oceanography, 60:223-243.
Ellen van Donk, Ramesh D. Gulati, Arjen Iedema, et al. 1993. Macrophyte-related shifts in the nitrogen and phosphorus contents of the different trophic levels in a biomanipulated shallow lakes[J]. Hydrobiologia, 251: 19-26.
FOLKE C, CARPENTER S, WALKER B, et al. 2004. Regime shifts, resilience, and biodiversity in ecosystem management[J]. Annual Review of Ecology and Systematics, 35:557-581.
GONZALEZ SMA, JAPPESEN E, GOMA J, et al. 2005. Does high nitrogen loading prevent clear water conditions in shallow lakes at moderately high phosphorous concentrations[J].? Freshwater Biology, 50:27-41.
Great Lakes Water Quality Board. 2003. Climate Change and Water Quality in the Great Lakes Basin: Report of the Great Lakes Water Quality Board to the International Joint Commission[R]. Canada: Great Lakes Water Quality Board, 135.
GROISMAN P Y, KNIGHT R W, KARL T R. 2001. Heavy precipitation and high streamflow in the contiguous United States: Trends in the twentieth century[J]. Bulletin of the American Meteorological Society, 82:219-246.
HARGEBY A, BLINDOW I, ANDERSSON G. 2007. Long-term patterns of shifts between clear and turbid states in lake Krankejön and lake Tåkern[J]. Ecosystems, 10:28-35.
HAVENS K E, AUMEN N G. 2000. Hypothesis driven experimental research is necessary for natural resource management[J]. Environmental Management, 25:1-7.
HAVENS K E, JIN K R, RODUSKY A J, et al. 2001. Hurricane effects on a shallow lake ecosystems and its response to a controlled manipulation of wate level[J]. The Scientific World, 1:44-70.
HOLLING C S. 1973. Resilience and Stability of Ecological Systems[J]. Annual Review of Ecology and Systematics, 4:1-2.
IBELINGS B W, PORTIELJE R, LAMMENS E, et al. 2007. Resilience of alternative stable states during the recovery of shallow lakes from eutrophication: Lake Veluwe as a case study[J]. Ecosystems, 10:4-16.
JEPPESEN E, JENSEN J P, BRANCO C W C. 2007. Restoration of shallow lakes by nutrient contrlo and biomanipulation-the successful strategy varies with lake size and climate[J]. Hydrobiologia, 581:269-285.
JEPPESEN E, JENSEN J P, KRISTENSEN P, et al. 1990. Fish manipulation as s lake restoration tool in shallow, eutrophic, temperate lakes 2: threshold levels, long-term stability and conclusions[J]. Hydrobiologia, (200/201): 219-227.
JEPPESEN E, SØNDERGAARD M, JENSEN J P, et al. 2005. Lake responses to reduced nutrient loading – an analysis of contemporary long-term data from 35 case studies[J]. Freshwter Biology, 50: 1747-1771.
JEPPOESEN E, JENSEN J P, KRISTENSEN P, et al. 1990. Fish manipulation as a lake restoration tool in shallow, eutrophic, temperate lakes 2: threshold levedl, long-term stability and conclusions[J]. Hydrobilologia, (200/201): 21-227.
JOHN D I, GERALD L W, PAUL F T. 1977. The Motion of a Small Sphere in a Rotating Velocity Field: A Possible Mechanism for Suspending Particles in Turbulence[J]. Journal of geophysical research, 82(15): 2096-2100.
JONES J I, SAYER C D. 2003. Does the fish-invertebrate-periphyton cascade precipitate plant loss in shallow[J]? Ecology, 84(8):2155-2167.
KAMENIR Y, WINDER M, DUBINSKY Z, et al. 2008. Lake Tahoe vs. Kinneret phytoplankton: comparison of long-term taxonomic size structure consistency[J]. Aquatic Sciences, 70:195-203.
KARL E. Havens, R. Thomas James, THERESE L. East, et al. 2003. N∶P ratios, light limitation, and cyanobacterial dominance in a subtropical lake impacted by non-point source nutrient pollution[J]. Environmental Pollution, 122: 379-390.
KOMATSU E, FUKUSHIMA T, HARASAWA H. 2007. A modeling approach to forecast the effect of long-term climate change on lake water quality[J]. Ecological Modelling, 209(2/4): 351-366.
KOSTEN S, LACEROT G, JEPPESEN E, et al. 209. Effects of submerged vegetation on water clarity across climates[J]. Ecosystems, 12:1117-1129.
KUMAGAI M, ISHIKAWA K, CHUNMENG J. 2003. Dynamics and biogeochemical significance of the physical environment in Lake Biwa[J]. Lakes Reserviors Research and Management, 7:345-348.
KUZNETSOV Y A. 1995. Elements of applied bifurcation theory[M]. Springer-Verlag
LAMMENS E, VAN NES E H, MEIJER M L, et al. 2004. Effects of commercial fishery on the bream population and the expansion of Chara aspera in Lake Veluwe[J]. Ecological Modelling, 177:233-244.
MALMAEUSA J M, BLENCKNERA T, MARKENSTENB H, et al. 2006. Lake phosphorus dynamics and climate warming: a mechanistic model approach[J]. Ecological Modelling, 190 (1/2): 1-14.
MARSDEN S. 1989. Lake restoration by reducing external phosphorus loading: the influence of sediment phosphorus release[J]. Freshwater Biology, 21:139-162.
MARTIN R. Perrow, Brian Moss, Julia Stansfield. 1994. Trophic interactions in a shallow lake following a reduction in nutrient loading: a long-term study[J]. Hydrobiologia, (275/276): 43-52.
MAY R M. 1977. Thresholds and breakpoints in ecosystems with a multiplicity of stable states[J]. Nature, 269: 471-477.
NAJJAR R G. 1999. The water balance of the Susquehanna River basin and its response to climate change[J]. Journal of Hydrology, 219:7-19.
NICHOLLS K H. 1999. Effects of temperature and other factors on summer phosphorus in the inner Bay of Quinte, Lake Ontario: implications for climate warming[J]. Journal of Great Lakes Research, 25(5): 250-262.
PERROW M R, MOSS B, STANSFIELD J. 1994. Trophic interactions in a shallow lake following a reduction in nutrient loading: a long-term study[J]. Hydrobiologia, (275/276): 43-52.
Roger W. Bachmann, Mark V. Hoyer, Daniel E. Canfield. 1999. The restoration of Lake Apopka in relation to alternative stable states[J]. Hydrobiologia, 394: 219-232.
SCHEFFER M, BAKEMA A H, WORTELBOER F G. 1993. MEGAPLANT-a simulation model for the dynamics of submerged plants[J]. Aquatic Botany, 45:341-356.
SCHEFFER M, BAKEMA A H, WORTELBOER F G. 2000. MEGAPLANT-Calado G, Duarte P. Modelling growth of Ruppia cirrhosa[J]. Aquatic Botany, 68:29-44.
SCHEFFER M, CARPENTER S R. 2003. Catastrophic regime shifts in ecosystems: linking theory to observation[J]. TRENDS in Ecology and Evolution, 18(12):648-656.
SCHEFFER M, CARPENTER S R. 2003. Catastrophic regime shifts in ecosystems: linking theory to observation[J]. Trends in ecology and evolution, 18(12):648-656.
SCHEFFER M, CARPENTER S, JONATHAN A F, et al. 2001. Catastrophic shifts in ecosystems[J]. Nature, 413:591-596.
SCHEFFER M, EGBERT H. van Nes. 2007. Shallow lakes theory revisited: various alternative regimes driven by climate, nutrients, depth and lake size[J]. Hydrobiologia, 584:455-466.
SCHEFFER M, HOSPER S H, MEIJER M-L, et al. 1993. Alternative equilibria in shallow lakes[J]. Trends in Ecology and Evolution, 8: 275-279.
SCHEFFER M, SZABÓ S, GRAGNANI A, et al. 2003. Floating plant dominance as a stable state[J]. PNAS, 100(7): 4040-4045.
SCHEFFER M. 1990. Multiplicity of stable states in a freshwater system[J]. Hydrobiologia, (200/201): 475-486.
SCHINDLER D W, HECHY R E. 2009. Eutrophication: more nitrogen data needed[J]. Science, 324:721-722.
SCHINDLER D W, HECKY R E, FINDLAY D L, et al. 2008. Eutrophication of lakes cannot be controlled by reducing nitrogen input: Results of a 37-year whole-ecosystem experiment[J]. PNAS, 105: 11254-11258.
SOLHEIM A L, REKOLAINEN S, MOE S J, et al. 2008. Ecological threshold responses in European lakes and their application for Water Framework Directive (WFD) implementation: synthesis of lakes results from the REBECCA project[J]. Aquatic Ecology, 42: 317-334.
SØNDERGAARD M, JEPPESEN E, MORTENSEN E, et al. 1990. Phytoplankton biomass reduction after planktivorous fish reduction in a shallow, eutrophic lake: a combined effect of reduced internal P-loading and increased zooplankton grazing[J]. Hydrobillogia, (200/201): 229-240.
VAN DEN BERG M S, SCHEFFER M, VAN NES E, et al. 1999. Dynamics and stability of Chara sp. and Potanmogeton petinatus in a shallow lake changing in eutrophication level[J]. Hydrobiologia, (408/409): 335-342.
VAN NES E H, RIP W J, SCHEFFER M. 2007. A theory for cyclic shifts between alternative states in shallow lakes[J]. Ecosystems, 10:17-27.
VAN NES E H, SCHEFFER M, VAN DEN BERG M S, et al. 2002. Dominance of charophytes in eutriphic shallow lakes-when should we expect it to be an alternative stable state[J].? Aquatic Botany, 72: 275-296.
VITOUSEK P M, MOONEY H A, LUBCHENCO J, et al. 1997. Human domination of earth’s ecosystems[J]. Science, 277: 494-499.
WALLSTEN M, FORSGEN P. 1989. The effects of increased water level on aquatic macrophytes[J]. J. Aquat. Plant Manage, 27: 32-37.
WALTER K D, WILLIAM H C, KEITH G, et al. 2010. Thresholds, breakpoints, and nonlinearity in freshwaters as related to management[J]. North American Benthological Society, 29(3): 988-997.
WINNIE J Rip, MAARTEN Ouboter, EGBERT H. van Nes, et al. 2005. Oscillation of a shallow lake ecosystem upon redunction in external phosphorus load[J]. Archives of Hydrobiology, 164(3):387-409.
YACOBI Y Z. 2006. Temporal and vertical variation of chlorophyⅡa concentration, phytoplankton photosynthetic activity and light attenuation in Lake Kinneret: possibilities and limitations for simulation by remote sensing[J]. Jouranl of Plankton Research, 28(8): 725-736.
ZIMMER K D, HANSON M A, HERWIG B R, et al. 2009. Thresholds and stability of alternative regimes in shallow Prairie-Parkland Lakes of central North America[J]. Ecosystems, 12:843-852.
ZIMMER K D, HERVIG B R, LAURICH L M. 2006. Nutrient excretion by fish in wetland ecosystems and its potential to support algal production[J]. Limnology and Oceanography, 51(1):197-207.
陈开宁, 包先明, 史龙新, 等. 2006. 太湖五里湖重建示范工程-大型围隔实验[J]. 湖泊科学, 18(2): 139-149.
冯剑丰, 王洪礼, 朱琳. 2009. 生态系统多稳态研究进展[J]. 生态环境学报, 18(4):1553-1559.
刘永定, 常毅峰, 潘晓洁, 等. 2007. 淡水生态系统稳态转换理论及其指导意义[D]. 中国海洋湖沼学会藻类分会第七届会员大会暨第十四次学术讨论会论文摘 要集.
秦伯强, 宋玉芝, 高光. 2006. 附着生物在浅水富营养化湖泊藻-草型生态系统转化过程中的作用[J]. 中国科学C辑: 生命科学, 36(3): 283-288.
秦伯强. 2002. 长江中下游湖泊富营养化发生机制与控制途径初探[J].湖泊科学, 14(3):193-201.
秦伯强. 2007. 湖泊生态恢复的基本原理与实现[J]. 生态学报, 27(11): 4848-4858.
舒庆. 2008. “十一五”环境规划汇编[M]. 北京: 红旗出版社.
张运林, 秦伯强, 胡维平, 等. 2006. 太湖典型湖区真光层深度的时空变化及其生态意义[J]. 中国科学D辑: 地球科学, 36(3):287-296.
Survey on Theory and Driving Factors of Regime Shifts on Lake Ecosystems
ZHAO Lei1, LIU Yong2, LI Yuzhao2, ZHU Xiang1*, ZOU Rui1
1. Yunnan Institute for Environmental Sciences, Yunnan Key Laboratory of Pollution Process and Management of Plateau Lake-Watershed (Prepare to Construct), Kunming 650034, China; 2. College of Environmental Science and Engineering, Peking University, The Key Laboratory of Water and Sediment Sciences, Ministry of Education, Beijing 100871, China
The lake ecosystem can undergo regime shifts triggered by long-term human threats and short-term strong disturbance. Before and after regime shifts, the ecosystem structure and key processes will change dramatically. For scientific management and decision making on lakes, it is therefore essential to reveal the mechanism and explore the driving factors of regime shifts. We concluded and discussed the concept and driving factors of regime shifts. Changing dramatically and difficult to forecast, ecosystem structure and key processes change, alternative stable state are main connotations of the concept of regime shifts. The characteristics of regime shifts include non-linear, alternative stable state, multiple thresholds, and hysteresis during restoration. There are external and internal factors in driving shifts between alternative states. The external driving factors include external nutrient loadings, climate change, storm, and water level. The influence of external nutrient loadings and climate change on lake ecosystem are long-term and accumulative. They can weaken the ecological resilience of lake to trigger regime shifts. Storm and water level are short-term factors and can trigger regime shifts shortly. The internal driving factors include fish, macrophyte. Fish can impose influence on macrophyte, lake sediment and zooplankton to trigger regime shifts. There is not only positive feedback effect of macrophyte on clear state of lake, also under certain conditions there is negative feedback effect. Some potential research focus was proposed based on the intensive literature review, including: (a) integrated influence of environmental and biological factors on macrophyte, (b) dominated states and key processes controlling the state based on large scale lake survey.
Lake ecosystem; regime shifts; driving factor
X171
A
1674-5906(2014)10-1697-11
赵磊,刘永,李玉照,朱翔,邹锐. 湖泊生态系统稳态转换理论与驱动因子研究进展[J]. 生态环境学报, 2014, 23(10): 1697-1707.
ZHAO Lei, LIU Yong, LI Yuzhao, ZHU Xiang, ZOU Rui. Survey on theory and driving factors of regime shifts on lake ecosystems [J]. Ecology and Environmental Sciences, 2014, 23(10): 1697-1707.
国家自然科学基金项目(41301599;4110180);国家水体污染控制与治理科技重大专项(2013ZX07102-006-002)
赵磊(1978年生),男,高级工程师,博士,主要从事湖泊污染过程机理与模拟研究。E-mail: zlzny@163.com
*通信作者(责任作者),E-mail: zx@yies.org.cn
2014-05-20
