三叶爬山虎叶片解剖结构和光合生理特性对3种生境的响应
2014-02-25吴涛耿云芬柴勇郝佳波袁春明
吴涛,耿云芬,柴勇,郝佳波,袁春明*
1. 云南省森林植物培育与开发利用重点实验室,国家林业局云南珍稀濒特森林植物保护和繁育实验室,云南 昆明 650201;2. 云南省林业科学院,云南 昆明 650201
三叶爬山虎叶片解剖结构和光合生理特性对3种生境的响应
吴涛1,2,耿云芬1,2,柴勇2,郝佳波1,2,袁春明1,2*
1. 云南省森林植物培育与开发利用重点实验室,国家林业局云南珍稀濒特森林植物保护和繁育实验室,云南 昆明 650201;2. 云南省林业科学院,云南 昆明 650201
藤本植物生活环境的时空变化较为剧烈,为适应异质性生境常表现出较大的可塑性,其形态解剖结构及光合生理特征被认为能很好地体现对异质生境的适应。为了明确藤本植物叶片结构和光合作用对不同生境光强的响应策略,以木质藤本三叶爬山虎(Parthenocissus himalayana)为对象,采用光合仪测定和解剖显微观察的方法研究了哀牢山亚热带湿性常绿阔叶林的林外(全光照)、林缘(遮荫)和林内(荫生)3种自然生境中三叶爬山虎的叶片解剖结构和光合生理特征的变化,以期阐述三叶爬山虎对不同光环境的生态适应能力及策略,为森林生态系统的管理和物种多样性的保护及群落优化配置提供理论依据。结果表明:从林内到林外随着生境光强增加,叶片厚度(157.77~299.17 μm)、上表皮厚度(21.30~28.40 μm)、栅栏组织厚度(30.83~124.65 μm)、栅栏组织细胞面积(430.95~652.97 μm2)显著增大(P<0.01),栅栏组织细胞长度(29.23~49.54 μm)和周长(86.58~155.17 μm)、下表皮厚度(16.14~19.01 μm)、气孔长度(24.13~27.10 μm)和气孔密度(86.20~129.41个·mm-2)呈显著上升趋势(P<0.05)。栅栏组织细胞宽度(19.67~22.81 μm)在3种生境中无显著差异。叶片解剖结构性状的平均可塑性值为0.37,其中最大值是栅栏组织细胞长度(0.67),最小值是气孔长度(0.11)。光饱和点(201.27~1299.17 μmol·m-2·s-1)、光补偿点(3.86~29.88 μmol·m-2·s-1)、饱和光强最大光合速率(2.20~12.03 μmol·m-2·s-1)、暗呼吸速率(0.17~2.19 μmol·m-2·s-1)、CO2补偿点(83.01~237.26 μmol·m-2·s-1)、饱和CO2最大净光合速率(2.07~25.49 μmol·m-2·s-1)、光呼吸速率(0.36~7.57 μmol·m-2·s-1)、初始羧化效率(0.006~0.035 μmol·μmol-1)随着生境光强的增高呈上升趋势,而表观量子效率(0.067~0.031 μmol·μmol-1)、CO2饱和点(2062.56~1385.31 μmol·m-2·s-1)呈下降趋势。与光合生理参数相关性显著的解剖性状主要是叶片厚度、上表皮厚度及其细胞周长/面积、栅栏组织细胞长度、栅栏组织厚度及维管束占叶脉面积的比例等。研究说明,三叶爬山虎在林外生境中具有明显的阳生叶特征,而在林内生境中具有明显的阴生叶特征,表现出对异质生境很强的适应性,从叶片解剖结构和光合生理特性方面解释了其在林外、林缘和林内均有分布的原因。
三叶爬山虎;叶片;解剖结构;光合特性;生境
木质藤本是森林植被中一类丰富多样但具有共同生长策略即攀援生长的植物类群,它们在森林的多样性、结构、功能和动态等方面具有重要的贡献和作用(王海艳和喻理飞,2010)。木质藤本植物由于受攀援能力和支柱木资源的限制,以及时空变化十分剧烈的生境特点,其形态和生理特征常发生明显的变化,表现出较大的可塑性。其中,叶片作为藤本植物对环境变化最为敏感的光合器官之一,其形态解剖结构及光合生理特征被认为最能体现环境因子的影响及植物对环境的适应(Mott等,1982;乌玉娜等,2010;庞杰等,2013)。亚热带常绿阔叶林林外、林缘、林内的生境具有较大的异质性,其中光照是影响藤本植物多度和分布的主要生境因子。木质藤本若要在这样较大异质性的生境中生长,较高的可塑性无疑将使其对环境具有较高的潜在适应能力,这对木质藤本在异质光环境中的生存至关重要(Strauss-Debenedetti和Bazzaz,1991)。
木质藤本以其特有的植物学特性和生态功能,在水土保持、防风固沙、减轻污染、垂直绿化和美化环境等方面异军突起,已经并将继续发挥其他植物不可替代的作用(李益锋等,2011)。然而,其有关研究工作在过去20年来才开始不断出现(Gentry,1991;蔡永立和宋永昌,2000)。目前有关木质藤本植物生态适应研究的一些结论主要来自于对热带藤本的研究,对其它地区藤本的研究则相对不足(蔡永立和宋永昌,2001);木质藤本植物间及其与木本植物间的比较生态研究较多,而对同种藤本植物不同生境下的光合生理和形态可塑性研究较少(夏江宝,2007;陈亚军等,2008);研究结果大多是在人工控制条件下获得的,缺少对木质藤本植物在野外原生环境中叶片形态结构及光合生理特征的观察和测定(江浩等,2011)。因此,本项研究以亚热带常绿阔叶林常见木质藤本三叶爬山虎(Parthenocissus himalayana)为材料,通过对其在不同生长光强的生境下叶片的光合作用和叶片解剖特征的研究,揭示其在不同光环境下的生态适应能力及策略,可为森林生态系统的管理和物种多样性的保护及群落优化配置提供科学依据。
1 材料与方法
1.1 研究地区自然概况
研究地点位于云南中南部景东县徐家坝的哀牢山国家自然保护区内的中国科学院哀牢山生态站附近的亚热带湿性常绿阔叶林,地理位置为24°32′ N,101°01′ E,海拔2450 m。本区属西南季风性气候,终年温凉潮湿。年平均温度为11.3 ℃,最冷月(1月)平均气温5.0 ℃,最热月(7月)平均气温15.3 ℃,极端最高气温25.0 ℃,极端最低气温-8.3 ℃。该区年降水量为1931.1 mm,雨季(5—10月)降水量占年降水总量的85%左右。年平均相对湿度为83%。≥10 ℃的的积温达3420 ℃,全年无霜期约180 d,日照时数为1239 h。林地土壤为山地黄棕壤。实验在8月上旬开展,最高温度为32.1 ℃,最低温度为16.0 ℃,正午最大光强可达2200 μmol·m-2·s-1;坡向为西坡,坡度10°~23°。土壤肥沃,偏酸性(pH为4.4~4.9),土壤表面由凋落物层(厚度3~7 cm)覆盖,土壤有机碳、全氮和全磷含量分别为12.91%、0.52%和0.06%,土壤表面的C/N比为14.4,30~50 cm深度C/N比为15.3(Liu等,2002.)。
研究地点所在研究区内有直径不小于1 cm、高度不小于2 m的木质藤本23种,隶属于16科、21属(袁春明等,2010)。在研究地点选取一个没有新近林窗干扰的森林边缘,并且其林缘、林内(林缘向内50 m外)和林外(林缘向外50 m外)均有自然生长的三叶爬山虎(直径1~2 cm)。林外、林缘、林内3种生境上午10:00时的光照强度和温度分别为1800 μmol·m-2·s-1和28 ℃、600 μmol·m-2·s-1和 25 ℃、100 μmol·m-2·s-1和22 ℃。
1.2 实验材料
选取林外、林缘、林内3种生境的三叶爬山虎植株距离地面60~100 cm处枝条的顶端向下第2~3片成熟叶为光合测定对象,每株1片,每种生境3~5个重复。光合测定结束后,将该叶片于中央位置处沿中脉剪取约1 cm2的样品放入FAA固定液,用于叶片解剖结构观察。
1.3 研究方法
1.3.1 光合生理特征测定
2013年8月10—16日,采用Li-6400便携式光合测定仪(LI-COR,USA),配合Li-6400-02B红蓝光源和Li-6400-01液化CO2钢瓶注入系统,对3种生境内相同地面高度的三叶爬山虎叶片进行光合-光响应曲线(Pn-PAR)和光合-CO2响应曲线(Pn-Ci)的测定。Pn-PAR曲线测定时,设置光合有效辐射梯度为0、25、50、75、100、150、200、300、400、600、800、1200、1600、2000 μmol·m-2·s-1,调整CO2浓度为400 µmol·mol-1,运用自动测量程序测定;Pn-Ci曲线测定时,光合有效辐射设定为1200 μmol·m-2·s-1,设置CO2浓度梯度为400、300、200、150、100、50、0、400、600、800、1200、1600 μmol·mol-1,运用自动测量程序测定。选取无病害植株的相同叶位的叶片分别挂牌标记,测定3个重复,测定时保持叶片自然生长角度不变。测定时间为9:00—12:00。根据Ye等(2012)的非直角双曲线修正模型拟合Pn-PAR曲线,计算饱和光强最大光合速率(Pmax)、光饱和点(LSP)、光补偿点(LCP)、表观量子效率(AQY)和暗呼吸速率(Rd)。根据叶子飘和于强(2009)的非直角双曲线修正模型拟合Pn-Ci曲线,计算饱和CO2最大光合速率(Amax)、CO2饱和点(Csp)、CO2补偿点(Ccp)、表观羧化效率(CE)和光呼吸速率(Rp)。
1.3.2 叶片解剖结构观察
光合生理特征测定后,剪取该叶片主脉及其两侧1.0 cm×1.0 cm的小块,用FAA固定,3个重复。采用常规石蜡法制片,在Leica显微镜下观察和拍照,测量并记录叶片的各结构,所得数据为10个视野的平均值。所观测的叶片解剖结构特征包括叶片厚度、上表皮厚度、上表皮细胞面积、上表皮细胞周长、栅栏组织厚度、栅栏组织细胞宽度、栅栏组织细胞长度、栅栏组织细胞周长、栅栏组织细胞面积、海绵组织厚度、下表皮厚度、下表皮细胞周长、下表皮细胞面积、维管束面积占叶脉面积的比例、气孔长度、气孔密度,共16项。可塑性指数=(某一指标或参数在3种生境下的最大值 - 最小值)/最大值。
1.3.3 数据处理和分析
运用SPSS 19.0进行数据处理和模式拟合及相关参数的计算,对3种生境下各指标或参数进行方差分析时用LSD方法。
2 结果与分析
2.1 叶片解剖结构特征在不同生境下的差异
三叶爬山虎在林外、林缘和林内3种生境下的叶片解剖结构见图1,观测结果见表1。可以看出,其叶片类型属于异面叶,叶肉组织分化为明显的栅栏组织和海绵组织,但各特征在3种生境条件下存在明显的差异。三叶爬山虎叶脉背面的腺毛在林外生境下较林缘和林内更明显且常见。林外生境下,栅栏组织细胞紧邻上、下表皮均有分布,上表皮处排列紧密,2~3层,下表皮处排列稀疏,1~2层;林缘生境下,栅栏组织细胞紧邻上表皮单层分布且排列较为疏松;林内生境下,栅栏组织细胞单层分布且排列十分疏松。
本研究所观测的16项叶片解剖结构特征,除栅栏组织细胞宽度在3种生境中无显著差异外,其余15项指标均存在显著或极显著差异。随着生境光照强度的降低,叶片厚度、上表皮厚度、栅栏组织厚度、栅栏组织细胞面积呈极显著下降,栅栏组织细胞长度/周长、下表皮厚度、气孔长度和气孔密度整体上呈显著下降趋势。海绵组织厚度在林外和林内相近,二者之间差异不显著,但较林缘的高且差异显著。下表皮细胞周长及面积以林缘最高且与林外和林内差异显著,林内和林外之间差异不显著。主脉维管束面积占叶脉面积的比例为林外>林内>林缘,两两之间差异显著。
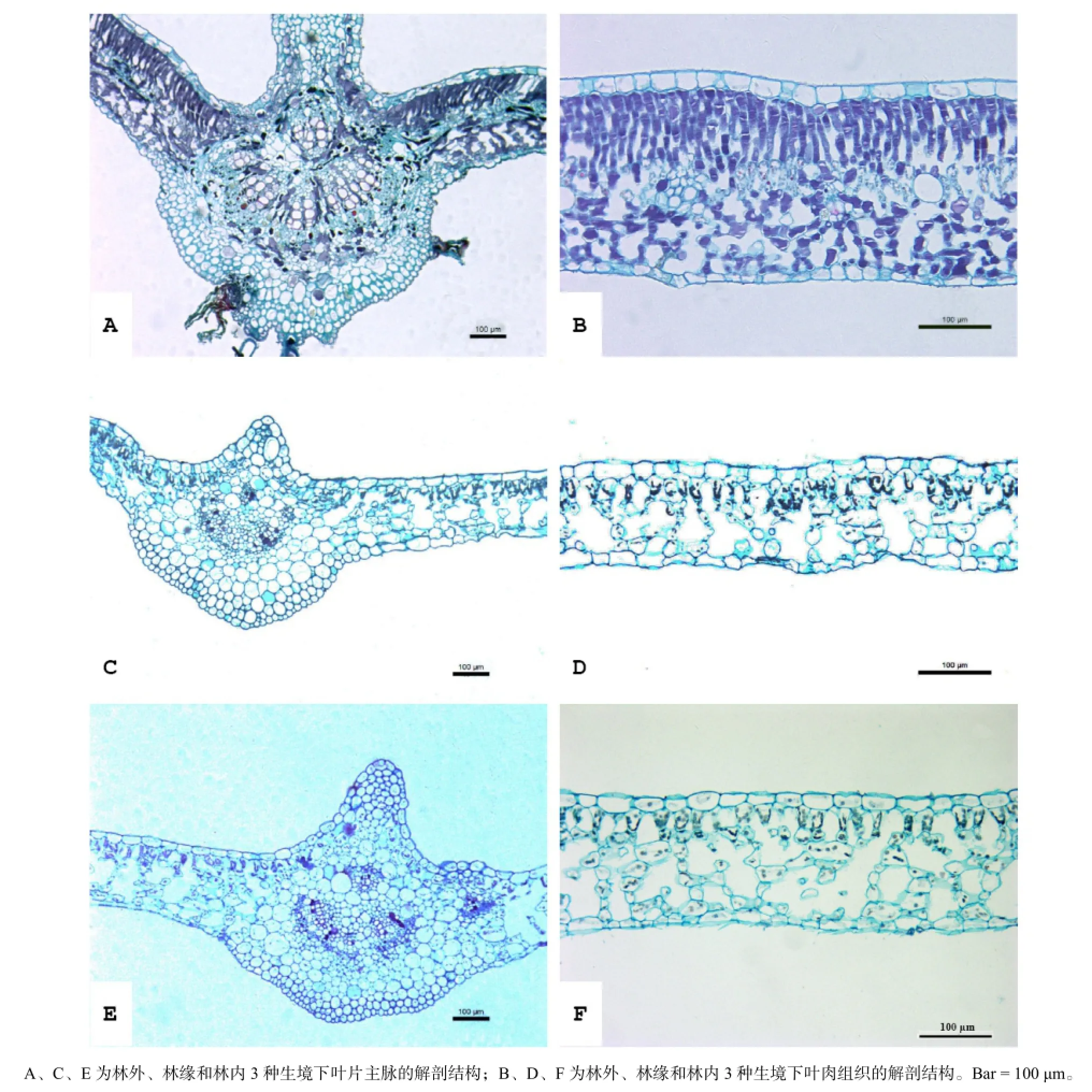
图1 三叶爬山虎在林外、林缘和林内3种生境下的叶片解剖结构Fig.1 Leaf anatomical structure of Parthenocissus himalayana in three different habitats

表1 三叶爬山虎在3种生境下的叶片解剖特征比较Table 1 The comparatives of leaf anatomical structure of Parthenocissus himalayana in three different light habitats
各指标的表型可塑性从大到小,依次为栅栏组织细胞长度>栅栏组织厚度>栅栏组织细胞周长>维管束面积占叶脉面积的比例>栅栏组织细胞面积>叶片厚度>上表皮细胞面积>下表皮细胞面积>气孔密度>上表皮细胞周长>上表皮厚度>栅栏组织细胞宽度>下表皮细胞周长>下表皮厚度>海绵组织厚度>气孔长度。综合方差分析和表型可塑性分析结果可以看出,栅栏组织细胞厚度/长度/周长/面积、叶片厚度、上表皮厚度、气孔密度和维管束面积占叶脉面积的比例是最能反映三叶爬山虎对生境响应的指标。
2.2 光合生理特性在不同生境下的差异
光补偿点越小,表明植物利用弱光的能力越强;而光饱和点越高则表明植物适应强光的能力越强。光合-光响应曲线参数(表2)表明,随着生境光强的降低,三叶爬山虎的光饱和点和光补偿点呈极显著下降趋势,而最大净光合速率、暗呼吸速率和表观量子效率呈显著下降趋势。三叶爬山虎林缘的最大净光合速率下降至林外的39%,表观量子效率降至其的56%,光饱和点降至其的59%,光补偿点降至其的25%,暗呼吸速率降至其的21%,表明三叶爬山虎从林外到林缘,主要通过降低光补偿点和暗呼吸速率来减缓其净光合速率的快速下降。
光合-CO2浓度响应曲线参数(表3)表明,三叶爬山虎林外的饱和CO2饱和净光合速率分别是林缘和林内的3.23倍和12.31倍,初始羧化效率分别是其4.38倍和5.83倍,光呼吸速率分别是其17.60倍和21.03倍,CO2补偿点分别是其3.32倍和2.86倍,而CO2饱和点分别是其78%和67%。参数显示,三叶爬山虎处于林缘和林内生境时,虽然饱和CO2净光合速率较林外生境显著降低,但其CO2补偿点也显著降低并且可利用的CO2浓度区间范围更大,光呼吸速率显著降低,即三叶爬山虎主要通过降低CO2补偿点和光呼吸速率来适应其饱和CO2净光合速率在林缘和林内生境时光强的减弱。

表2 三叶爬山虎在3种生境下的光合-光响应曲线特征参数Table 2 The parameters of light response curve (Pn-PAR) of Parthenocissus himalayana in three different light habitats

表3 三叶爬山虎在3种生境下的光合-CO2响应曲线特征参数Table 3 The parameters of CO2response curve (Pn-Ci) of Parthenocissus himalayana in three different light habitats
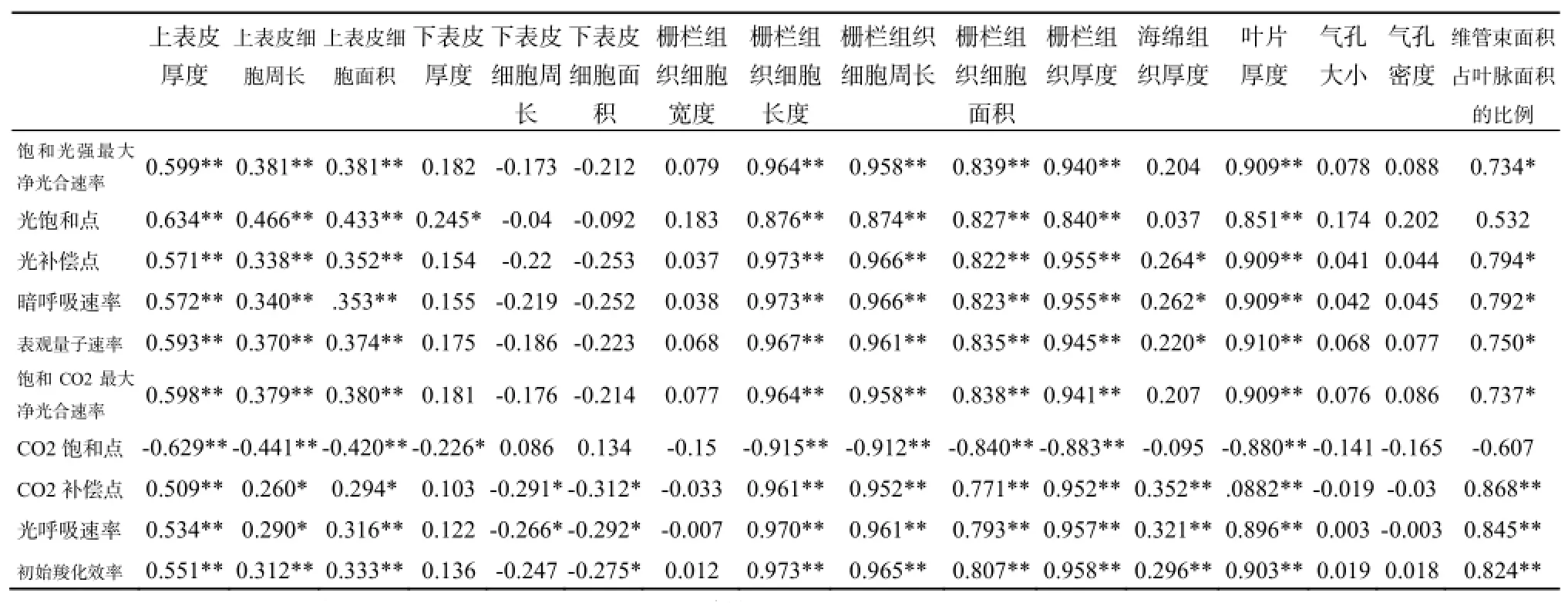
表4 不同生境下三叶爬山虎叶片解剖结构和光合生理参数之间的相关性分析Table 4 The correlation analysis between photosynthetic parameters and anatomical structures characteristics of Parthenocissus himalayana in three different light habitats
2.3 不同生境下三叶爬山虎叶片解剖结构和光合生理参数的关系
如表4所示,10个光合生理参数与16个叶片解剖性状之间的160对相关性分析结果中,显著性相关(P<0.05)18对,极显著相关(P<0.01)83对。其中,与光合生理参数相关性显著和极显著的解剖性状主要是上表皮厚度及其细胞周长/面积、栅栏组织细胞长度/周长/面积、叶片厚度、栅栏组织厚度及维管束占叶脉面积的比例;海绵组织厚度、气孔大小及密度、下表皮厚度及其细胞周长/面积、栅栏组织细胞宽度与光合生理参数几乎无显著(P<0.05)相关性。说明海绵组织厚度和气孔大小及其密度等解剖结构不影响三叶爬山虎在不同光强生境下的光合作用,而上表皮厚度及其细胞周长/面积、栅栏组织厚度及其细胞长度/周长/面积等解剖结构则有较明显的影响。
3 讨论
3.1 解剖结构对不同生境的响应
叶片解剖结构能够反映植物对环境变化的响应(李芳兰和包维楷,2005;季子敬等,2013)。厚叶结构、发达的栅栏组织、排列紧密平直的表皮细胞是阳生叶的典型特征,而发达的海绵组织、凸透形的表皮细胞则是叶片对于弱光、水分充足的一种适应特性(贺金生等,1994;王荣和郭志华,2007;陈模舜和柯世省,2013)。本项研究中,三叶爬山虎叶片厚度表现出随生境光强增强而变厚的趋势,且差异极显著;林外叶片厚度为林内的1.90倍。造成其差异的主要原因是栅栏组织厚度和表皮(尤其是上表皮)厚度的增加;栅栏组织厚度的增加主要由栅栏组织细胞层数增加所致,栅栏组织细胞长度也起到一定作用。海绵组织厚度表现出随生境光强减弱而增大的趋势,差异显著。此外,随生境光强减弱栅栏组织细胞排列呈现出由紧密向疏松变化的趋势;形状由长棒状趋于短锥状;表皮细胞的细胞壁由平直趋于弧形弯曲(图1)。这些结果表明,三叶爬山虎在林外生境中具有明显的阳生叶特征,而在林内生境中表现出明显的阴生叶特性,表现出对异质生境的良好适应性。
植物生长在高光强下气孔密度一般都比弱光强下高,如枫香(Liquidambar formosana) (王荣和郭志华,2007)和白三叶(Trifolium repens)(史刚荣和蔡庆生,2006),与本研究结果一致,推测这样有利于改善在较高光强下由于具有较大的叶肉表面积而造成CO2扩散受阻的情况(史刚荣和蔡庆生,2006)。但豚草(Ambrosia cordifolia)叶片在遮荫条件下为气孔下生,在高光强时则为气孔双生,且上表面的气孔密度在高光强时降低,而下表面的则增加(Mott等,1982)。这说明光强对气孔密度的影响因物种不同而不同。
3.2 光合特性对不同生境的响应
光合-光响应和光合-CO2响应参数提供了植物对变化的环境条件适应能力的光合特性信息,反映了植物对光照强度和生境大气CO2浓度的适应能力,在一定程度上决定了植物的分布模式和物种丰度。植物的光饱和点(LSP)表示对光照强度的需要量,其值越高表示对光强的需求越大。植物光补偿点(LCP)和表观量子效率(AQY)反映其对弱光的适应能力,LCP值越小、AQY越大,表明植物利用弱光的能力越强(柴胜丰等,2013;陈模舜和柯世省,2013)。本项研究中,三叶爬山虎的LSP在3种光照生境下差异显著,林外的LSP是林缘的1.7倍、林内的6.5倍。林外的光补偿点LCP显著高于林缘,但林缘和林内差异不显著。三叶爬山虎在林外的LSP高而林内的LCP低,说明其对光强范围的适应能力广;其在林内的LSP和LCP均显著低于林外,而林缘和林内的AQY显著高于林外,说明其对弱光的利用能力强,既能适应林外的全光照光强又能适应林内遮荫条件。这与夏江宝(2007)的研究结果一致,其认为三叶爬山虎耐荫性较强,维持较高光合速率的适宜光强范围为600~1600 µmol·m-2·s-1。
3.3 三叶爬山虎对生境光强的适应策略
植物表型可塑性与其环境适应能力关系紧密,是衡量植物对环境响应的重要指标,可塑性值大在一定程度上表明植物适应异质生境的能力强(薛静,2010)。枫香的叶片解剖特征在异质光生境间的总体可塑性值为0.30(王荣和郭志华,2007),白三叶的为0.26(史刚荣和蔡庆生,2006)。三叶爬山虎的总体可塑性值为0.37,大于枫香和白三叶,一定程度上表明藤本植物对异质光生境的适应能力强于木本植物和草本植物。另外,三叶爬山虎的可塑性值最大的是栅栏组织细胞长度,可塑性值较小的是气孔长度、海绵组织厚度和下表皮厚度,均与枫香和白三叶相同,表明植物在叶片解剖结构上响应异质光生境的策略相同,主要是通过增加栅栏组织细胞长度和栅栏组织厚度增加叶片厚度,从而适应更强的光照环境。栅栏组织细胞长度和层数在植物适应异质光环境方面起到了极其重要的作用。
三叶爬山虎在哀牢山常绿阔叶林的分布模式为在林窗边缘(等同于本研究中的林缘)和林内分布较多而在林窗(等同于本研究中的林外)较少(袁春明等,2008)。本项研究中,在10:00~14:00光照较强时,林外生境的光强有时会超过光饱和点,林缘生境的光强接近但小于光饱和点,林内生境的光强则仅约为光饱和点的一半,三叶爬山虎在林外比林缘和林内更整齐平直的上表皮细胞、更长和更多层数的栅栏组织细胞、更大的光饱和点,这些均可视为其对林外高光强和林内弱光强的适应,因此可以认为三叶爬山虎能够适应林外、林缘和林内的光照条件,而其在林外分布较少可能是攀附生长所需的支持物缺少的关系(袁春明等,2010)。
4 结论
1)三叶爬山虎表现出对异质光强生境的良好适应性,在林外全光照生境下呈现厚叶结构、发达的栅栏组织、排列紧密平直的表皮细胞等阳生叶特征;在林内荫蔽生境下呈现叶片变薄、海绵组织发达、栅栏组织细胞排列疏松、表皮细胞壁弧形弯曲等阴生叶特征。造成三叶爬山虎叶片厚度随生境光强的增强而显著变厚的主要原因是栅栏组织厚度和表皮(尤其是上表皮)厚度的增加;同时,栅栏组织厚度的增加主要由栅栏组织细胞层数增加所致,栅栏组织细胞长度也起到一定作用。
2)从林外到林内,三叶爬山虎主要通过降低光补偿点、暗呼吸速率、CO2补偿点和光呼吸速率,增加CO2浓度使用区间来适应逐渐减弱的光强生境。
3)相关性分析表明,与光合生理参数相关性极显著/显著的叶片解剖性状主要是上表皮厚度及其细胞周长/面积、栅栏组织细胞长度/周长/面积、叶片厚度、栅栏组织厚度及维管束占叶脉面积的比例;栅栏组织细胞宽度、气孔大小及密度与光合生理参数的相关性不显著。
4)可塑性值分析表明,三叶爬山虎从叶片解剖结构上响应异质光生境的策略主要是通过增加栅栏组织细胞长度和栅栏组织厚度以增加叶片厚度,从而适应更强的光照环境。栅栏组织细胞长度和层数在植物适应异质光环境方面起到了极其重要的作用。
致谢:试验样地选择和光合生理参数的测定得到中国科学院哀牢山森林生态系统研究站的大力支持,特表感谢!
GENTRY A H. 1991. The distribution and evolution of climbing plants[A]//PUTZ F E and MOONEY H A. The Biology of Vines[M]. Cambridge: Cambridge University Press: 3-49.
LIU W Y, FOX J E D, XU Z F. 2002. Biomass and nutrient accumulation in montane evergreen broad-leaved forest (Lithocarpus xylocarpus type) in Ailao Mountains, SW China[J]. Forest Ecology and Management, 158: 223-235.
MOTT K A, Gibson A G, O' Leary J W. 1982. The adaptive significance of amphistomatic leaves [J]. Plant Cell Environ, 9: 455-460.
STRAUSS-DEBENEDETTI S, Bazzaz F A. 1991. Plasticity and acclimation to light in tropical Moraceae of different successional positions [J]. Oecologia, 87: 377-387.
YE Z P, YU Q, KANG H J. 2012. Evaluation of photosynthetic electron flow using simultaneous measurements of gas exchange and chlorophyll fluorescence under photorespiratory conditions[J].Photosynthetica, 50(3): 472-476.
YE Z P. 2012. Nonlinear optical absorption of photosynthetic pigment molecules in leaves[J]. Photosynthesis Research, 112(1): 31-37.
蔡永立, 宋永昌. 2000. 中国亚热带东部藤本植物的多样性[J]. 武汉植物研究, 18(5):15-18.
蔡永立, 宋永昌. 2001. 浙江天童常绿阔叶林藤本植物的适应生态学 I.叶片解剖特征的比较[J]. 植物生态学报, 25(1): 90-98.
柴胜丰, 庄雪影, 韦霄, 等. 2013. 光照强度对濒危植物毛瓣金花茶光合生理特性的影响[J] 西北植物学报, 33(3): 547-554.
陈模舜, 柯世省. 2013. 天台鹅耳枥叶片的解剖结构和光合特性对光照的适应[J]. 林业科学, 49(2): 46-53.
陈亚军, 朱师丹, 曹坤芳. 2008. 两种光照下木质藤本和树木幼苗的生理生态学特征[J]. 生态学报, 28(12): 6034-6042.
贺金生, 陈伟烈, 王勋陵. 1994. 高山栎叶的形态结构及其与生态环境的关系[J]. 植物生态学报, 18(3): 219-227.
季子敬, 全先奎, 王传宽. 2013. 兴安落叶松针叶解剖结构变化及其光合能力对气候变化的适应性[J]. 生态学报, 33(21): 6967-6974.
江浩, 周国逸, 黄钰辉, 等. 2011. 南亚热带常绿阔叶林林冠不同部位藤本植物的光合生理特征及其对环境因子的适应[J]. 植物生态学报, 35 (5): 567-576.
李芳兰, 包维楷. 2005. 2011. 植物叶片形态解剖结构对环境变化的响应与适应[J]. 植物学通报, 22(Z): 118-127.
李益锋, 陶抵辉, 王绍卿. 2011. 藤本植物异军突起 藤本产业值得开发[J]. 江苏农业科学, 39(4): 5-8.
庞杰, 张凤兰, 郝丽珍, 等. 2013. 沙芥幼苗叶片解剖结构和光合作用对干旱胁迫的响应[J]. 生态环境学报, 22(4): 575-581.
史刚荣, 蔡庆生. 2006. 白三叶叶片解剖可塑性及其对光强的响应[J].草地学报, 14(4): 301-305.
王海艳, 喻理飞. 2010. 不同光环境下3种藤本植物幼苗的光合特性[J].浙江林学院学报, 27(6): 858-864.
王荣, 郭志华. 2007. 不同光环境下枫香幼苗的叶片解剖结构[J]. 生态学杂志, 26(11): 1719-1724.
乌玉娜, 陶建平, 赵科, 等. 2010. 海南霸王岭天然次生林边缘效应下木质藤本的变化[J]. 林业科学, 46(5): 1-6.
夏江宝. 2007. 北方主要木质藤本植物光合效率及其对水分与光照的响应[D]. 泰安: 山东农业大学: 1-286.
薛静,王国骄,李建东,等. 2010. 不同水分条件下三裂叶豚草叶解剖结构的生态适应性[J]. 生态环境学报, 19(3): 686-691.
叶子飘, 于强. 2009. 植物气孔导度的机理模型[J]. 植物生态学报, 33 (4): 772-782.
袁春明, 刘文耀, 杨国平, 等. 2010. 哀牢山湿性常绿阔叶林木质藤本植物的物种多样性及其与支柱木的关系. 林业科学, 46(1): 15-22.
袁春明, 刘文耀, 杨国平. 2008. 哀牢山湿性常绿阔叶林林窗木质藤本植物的物种组成与多样性[J]. 山地学报, 26(1): 29-35.
Response of Leaf Anatomical Structure and Photosynthesis Characteristics of Parthenocissus himalayana to Three Habitat Types
WU Tao1,2, GENG Yunfen1,2, CHAI Yong2, HAO Jiabo1,2, YUAN Chunming1,2*
1. Yunnan Laboratory for Conservation of Rare, Endangered & Endemic Forest Plants, Public Key Laboratory of the State Forestry Administration; Yunnan Provincial Key Laboratory of Cultivation and Exploitation of Forest Plants, Kunming 650201, China; 2. Yunnan Academy of Forestry, Kunming 650201, China
Temporal and spatial changes of habitats of vine plants are often intense, so plants should exhibit great plasticity in order to adapt to heterogeneous habitats. The anatomical structures and photosynthetic characteristics of leaves are considered to greatly reflect the adaptation of plants to different environmental regimes. The objective of the present study was to compare the anatomic and photosynthetic properties of the leaves of liana Parthenocissus himalayana grown in different locations (field, forest edge, and forest interior in the Ailao Mountains, Yunnan, China) with paraffin sectioning and Li-6400 photosynthetic apparatus. The reactions of P. himalayana to different light environments and its survival mechanisms were evaluated. A significant correlation was found between the anatomical structure characteristics and the photosynthesis physiological properties. The intensification of light, from the forest interior to the field, caused a considerable increase in the thickness of the whole leaf and the tissues within upper epidermis, palisade tissue, and palisade cells area, from 157.77 to 299.17 μm, 21.30 to 28.40 μm, 30.83 to 124.65 μm, 430.95 to 652.97 μm2, respectively. There was also a significant rise in the length (from 29.23 to 49.54 μm) and circumference of palisade cell (from 86.58 to 155.17 μm). The thickness of the lower epidermis and the stomatal size and density were substantially increased. There were no significant differences in width of palisade cells (19.67-22.81 μm) among different habitats. The average value of plasticity indexes of leaf anatomical characteristics was 0.37, among which the length of palisade cells had the maximum value (0.67) and the size of stoma had the minimum one (0.11). The light saturation point (LSP), light compensation point (LCP), and the maximum photosynthetic rate under saturation light intensity (Pmax) were markedly elevated. Higher were also the values of a number of indicators, including the dark respiration rate (Rd), CO2compensation point (Ccp), light respiration rate (Rp), and initial carboxylation efficiency (CE). These results indicated that Parthenocissus himalayana have a remarkable light adaptability in heterogeneous habitats in terms of leaf anatomical structure and photosynthetic characteristics.
Parthenocissus himalayana; leaf; anatomical structure; photosynthesis characteristics; habitat
Q945.79
A
1674-5906(2014)10-1586-07
吴涛,耿云芬,柴勇,郝佳波,袁春明. 三叶爬山虎叶片解剖结构和光合生理特性对3种生境的响应[J]. 生态环境学报, 2014, 23(10): 1586-1592.
WU Tao, GENG Yunfen, CHAI Yong, HAO Jiabo, YUAN Chunming. Response of leaf anatomical structure and photosynthesis characteristics of Parthenocissus himalayana to three habitat types [J]. Ecology and Environmental Sciences, 2014, 23(10): 1586-1592.
国家自然科学基金项目(31160136)
吴涛(1979年生),男,助理研究员,博士,主要从事林木遗传育种和植物生理学研究。E-mail:ynafwt@126.com
*通信作者:袁春明(1964年生),男,副研究员,博士,主要从事植物生态、恢复生态和保护生物学等方面的研究。E-mail:yuanchunming1017@163.com
2014-09-09
