放牧对荒漠草原建群种和优势种生态位的影响
2014-02-25吕世杰刘红梅吴艳玲卫智军孙世贤王颖杰林森
吕世杰,刘红梅,吴艳玲,卫智军,孙世贤,王颖杰,林森
1. 内蒙古农业大学理学院,内蒙古 呼和浩特 010018;2. 内蒙古农业大学生态环境学院,内蒙古 呼和浩特 010019;3. 内蒙古林业科学研究院,内蒙古 呼和浩特 010010;4. 内蒙古师范大学生命科学与技术学院,内蒙古 呼和浩特 010022
放牧对荒漠草原建群种和优势种生态位的影响
吕世杰1,刘红梅2,3,吴艳玲2,4,卫智军2*,孙世贤2,王颖杰2,林森1
1. 内蒙古农业大学理学院,内蒙古 呼和浩特 010018;2. 内蒙古农业大学生态环境学院,内蒙古 呼和浩特 010019;3. 内蒙古林业科学研究院,内蒙古 呼和浩特 010010;4. 内蒙古师范大学生命科学与技术学院,内蒙古 呼和浩特 010022
短花针茅荒漠草原建群种为短花针茅(Stipa breviflora),优势种为无芒隐子草(Cleistogenes songorica)和碱韭(Allium polyrrhizum),3个种群地上现存量之和可达群落现存量的60%~80%,其数量消长、时空变化及结构的位移均会引起群落的巨大波动。为明确3个植物种群占有空间资源的能力、相互关系及其受放牧影响的变化规律,以内蒙古苏尼特右旗赛汉塔拉镇1999年建立的荒漠草原教学科研基地建群种和优势种为研究对象,于2012年8月在围封区(CK)和自由放牧区(CG)按样带法和样地法进行取样,运用生态位宽度指数、生态位重叠指数及灰色关联度进行数据分析,结果如下。受放牧影响,短花针茅和碱韭主要通过大幅度调节生态位宽度来保证植物种群存活及其在群落中的地位和作用,变化幅度分别为0.429~0.813和0.235~0.828;而无芒隐子草生态位宽度变化不大(为0.556~0.761),其对放牧影响的耐受性较强。在生态位重叠方面,短花针茅与无芒隐子草受放牧影响后分别与其他两植物种群重叠程度增加,而碱韭植物种群表现相反;生态位重叠程度变化与种对间的竞争能力有关,也与种对间的亲和能力有关,且存在不对称性。
短花针茅荒漠草原;建群种;优势种;生态宽度;生态位重叠,灰色关联分析
随着生态位理论的不断完善和发展,生态位研究已成为近代生态学理论的一个主要内容(李军玲等,2003;苏鹏飞等,2012),其在理解群落结构功能、群落内物种种间关系、生物多样性、群落动态演替和种群进化等方面具有重要的作用(张金屯,2011;刘贵河等,2013)。为此,放牧利用为主的草地生态系统,研究者对植物种群生态位研究更加关注放牧家畜影响下的植物种群生态位宽度和生态位重叠程度响应特点,且集中在放牧压(王仁忠,1997;董全民等,2007;刘贵河等,2013)和放牧制度(刘红梅,2011)两方面,以便探寻不同放牧压或不同放牧制度对草地植物种群的影响程度及其响应规律。
然而,植物种群生态位变化与其所处空间的资源状态密不可分,且同种间竞争紧密联系在一起(Silvertown,1983;Abrams,1987;汪殿蓓等,2005)。因此,当前生态位概念是指该种在群落中利用资源的能力,这种能力不但体现在该种个体在群落中的分布范围和生物量的占有上,而且也体现在资源有限时对环境的耐受性上(张金屯,2011)。所以,放牧草地植物种群生态位研究缺少与资源状态相结合的探讨,同时由于种间竞争关系存在不对称性(Weiner,1990;Connolly和Wayne,1996;Freckleton和 Watkinson,2001),国内外缺乏基于生态位理论分析为基础的种间竞争关系阐释,以生态位理论分析放牧草地植物种群生态位变化能够丰富这方面的研究内容。
放牧利用为主的短花针茅荒漠草原地处亚洲中部草原区向荒漠区的过渡地带,由于其旱生生境及地域过渡性,显示出生态学上的独特性和脆弱性(卫智军等,2013);建群种为短花针茅(Stipa breviflora),优势种为无芒隐子草(Cleistogenes songorica)和碱韭(Allium polyrrhizum),3个种群地上现存量可达群落现存量的60%~80%,其数量消长、时空变化及结构的位移均会引起群落的巨大波动,构成了短花针茅+无芒隐子草+碱韭群落类型。通过研究3个种群生态位宽度和生态位重叠程度空间变化规律,结合种间关联度分析,可以阐释短花针茅荒漠草群落空间结构变化及波动情况,同时明确3个种群的生态位变化及其与种间关联的关系。
有鉴于此,本研究采用自由放牧作为试验处理,以短花针茅荒漠草原建群种和优势种为研究对象,选择代表性样地进行空间样带和空间范围取样,运用生态位宽度指数和生态位重叠指数,结合灰色关联度进行数据分析,阐明短花针茅荒漠草原建群种和优势种生态位受放牧影响与否的空间变化特点和规律,揭示3个植物种群种间关系及群落的波动情况,为草地环境保护和草地畜牧业可持续发展提供理论依据和数据支持。
1 试验设计与研究方法
1.1 试验地概况
放牧试验在1999年建立的内蒙古农业大学苏尼特右旗教学科研基地进行,行政区划属锡林郭勒盟苏尼特右旗赛汉塔拉镇哈登胡舒嘎查,地理位置为E112°47′16.9″,N42°16′26.2″。海拔在1100~1150 m之间,地势平缓且覆盖大量的第三纪中生代和第四纪残积物。土壤为淡栗钙土,是草原向荒漠过渡的地带性土壤,地表沙化严重。年均降水量183.0 mm,年蒸发量2793.4 mm,年平均气温5.8 ℃,日照时数3137.3 h,≥5 ℃的有效积温3426.0 ℃,≥10 ℃的有效积温2491 ℃,无霜期177 d,属中温型气候。试验区植被以亚洲中部荒漠草原植物种占主导地位,草层低矮,一般高度为10~35 cm,盖度为10%~50%。
1.2 试验设计及数据采集方法
放牧试验分别设对照(CK)和自由放牧(CG)2个试验处理,围栏封育区(CK)100 m×100 m,自由放牧区草场面积为438 hm2,全年实行自由放牧,全年载畜率为1.25只/hm2。
(1)样带法取样
在CK处理内和CG区选择一90 m×90 m代表性样地,分别以CK区或CG区样地西侧为初始样带,以西侧和南侧交汇处设为初始样点,每条样带共设10个1 m×1 m样方,每两个样方间隔9 m,2012年8月按种调查不同植物种群的株丛数。每两条样带同样间隔9 m,共设10条样带。
(2)样地法取样
将两区初始样带定为初始样地面积,以后依次增加一条样带,与初始样带及增加的样带合并成为新的样地面积。因此,第一条样带面积为90 m2,紧邻第一条样带的第二条样带与第一条样带同时作为新的样地空间范围,其面积为900 m2,以此类推。
样带法空间范围取样详见表1。
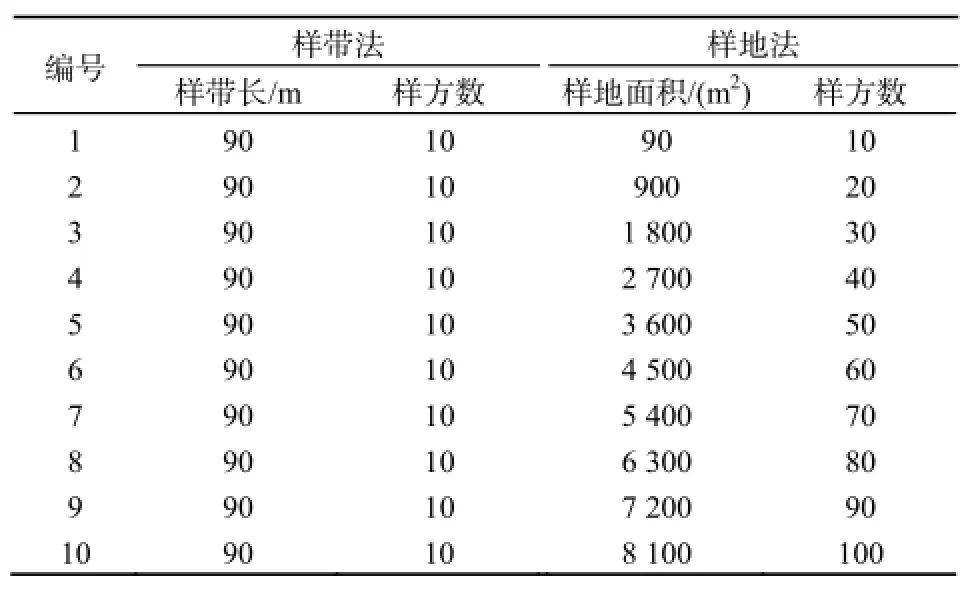
表1 样带法和样地法取样结果Table 1 Transect and plot sampling results
1.3 数据分析
(1)Levins生态位宽度指数(付卫国等,2008):
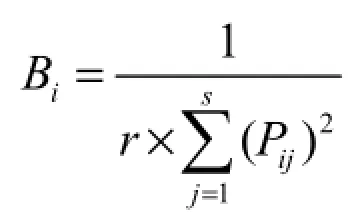
这里Bi为种i的生态位宽度;Pij=nij/Ni+,它代表种i在第j个资源状态下的个体数占该种所有个体数的比例,r为样方数。Levins生态位总宽度(牛克昌等,2009):
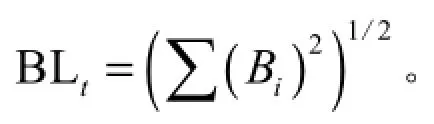
(2)Levins生态位重叠指数(张金屯,2011):
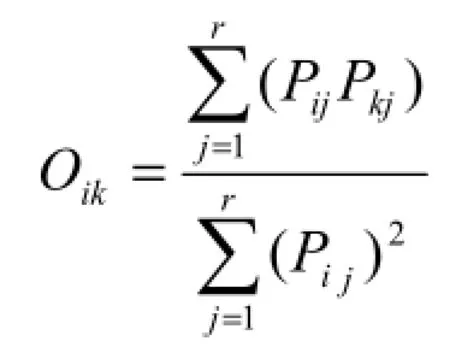
这里Oik代表种i的资源利用曲线与种k的资源利用曲线的重叠指数。从上式的分母可以看出,该指数实际上与种i的生态位宽度有关,由上式可知,Oik和Oki的值是不同的,含义也有异。Oik表示植物种群i占有其他植物种群j资源状态的比例情况,反之成立。
(3)关联度分析
将不同样线、样地范围的主要植物种群生态位宽度指数进行关联度分析,具体步骤如下:
①按照表1中序号计算得到的主要植物种群生态位宽度指数严格排列,构造成为原始数据集。数据集样式如下:
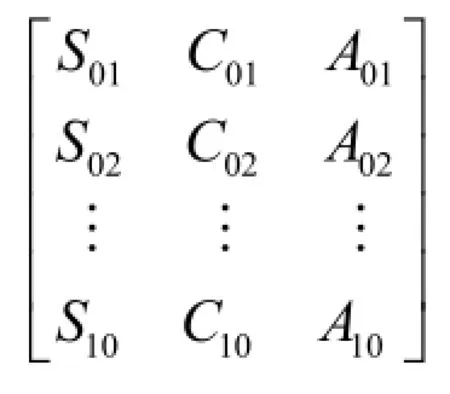
②采用均值化将原始数据进行变换,并进行初始化,得到绝对值差比较序列,样式如下:

③计算关联系数,公式为:
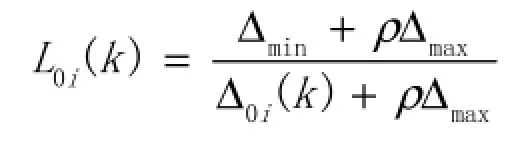
式中Δ0i(k)表示k种群两两比较序列的绝对差,ρ称为分辨系数,本研究中取值0.3,Δmax和Δmin分别表示比较序列绝对差中的最大值与最小值。
⑤“优劣”关系比较,若r0S=r0C,则称X序列{XS}对于序列{X0}等价于(或等于){XC}。若有r0S≥r0C,称序列{XS}对于序列{X0}优于或等于{XC}。若有r0S≤r0C,则称序列{XS}对于序列{X0}劣于或等于{XC}。
⑥返回②逐列进行,直到最后一列分析完毕,得到关联矩阵。
2 结果分析
2.1 植物种群在不同样带的生态位宽度变化

图1 不同样带主要植物种群生态位宽度变化Fig. 1 The niche width changes of main populations in different transect
将CK、CG处理内不同样带上建群种和优势种计算所得生态位宽度绘制成图,结果见图1。由图1可以看到,CK处理内碱韭的生态位宽度在不同样带上的差异最小,而短花针茅和无芒隐子草的生态位宽度在不同样带上的差异大于碱韭,这表明碱韭生态位宽度在CK处理内受不同样带资源差异影响较小,碱韭对空间资源差异的耐受性较强,其生态位宽度可以维持在相对稳定的水平上。在CG处理内,短花针茅植物种群受资源差异和放牧家畜同时影响,其生态位宽度在各个样带上相对稳定,无芒隐子草生态位宽度在样带间的波动情况大于短花针茅,但小于碱韭。
综合来看,短花针茅、无芒隐子草和碱韭生态位宽度均受植物种群所处资源状态的空间差异影响;但当存在放牧家畜干扰的时候,短花针茅、无芒隐子草和碱韭生态位宽度波动幅度发生变化,因此,建群种和优势种受生物和非生物因素同时影响,生态位宽度可能相同,也可能不同。
尽管不同样带态位宽度指数波动较大,且受放牧影响发生较大变化,但关联度分析结果显示,其存在明显的关联性(见表2)。在CK条件下,短花针茅与无芒隐子草的相互关联程度相等,均为0.4587;短花针茅与碱韭的关联度大于碱韭与短花针茅的关联度,分别为0.4529和0.4296;无芒隐子草与碱韭的关联度大于碱韭与无芒隐子草的关联度,分别为0.5697和0.5485。在CG条件下,短花针茅与无芒隐子草的关联度小于无芒隐子草与短花针茅的关联度,分别为0.6532和0.6570,说明受自由放牧影响,短花针茅与无芒隐子草之间生态位宽度变化由对称的转为非对称的,其种间关系表现也由对称性转为非对称性;短花针茅与碱韭的关联度不但也表现出不对称性,而且这种不对称性发生了逆转,这表明短花针茅和碱韭的生态宽度变化表现明显相反且变动幅度较大;无芒隐子草与碱韭的关联度和碱韭与无芒隐子草关联度由不对称转为对称,且关联程度增加。因此,3个植物种群3对6个关联度受放牧影响均已发生变化。同时可以看到,CG条件下种间生态位关联度均大于CK条件,表明自由放牧导致种间关联度增加。
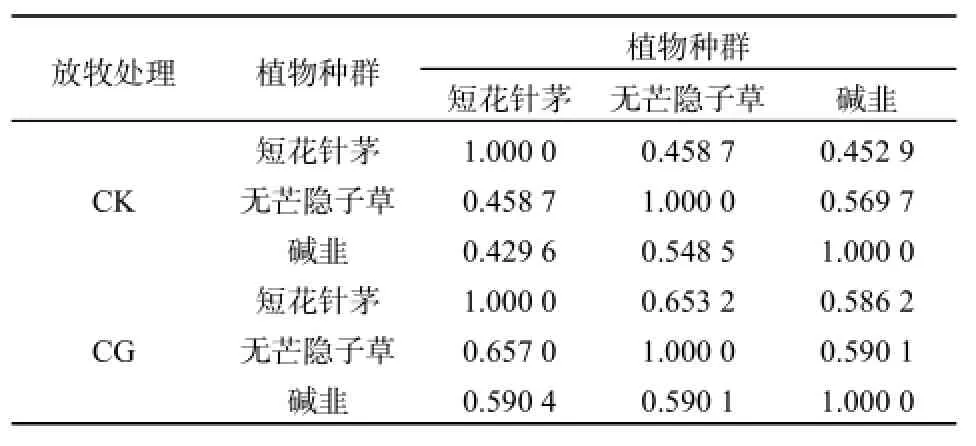
表2 不同样带主要植物种群生态位宽度关联情况Table 2 The grey correlation of niche width of main plant populations in different transect
短花针茅、无芒隐子草和碱韭在CK条件下的生态位总宽度为短花针茅<无芒隐子草<碱韭(见图2);在CG条件下,短花针茅、无芒隐子草和碱韭生态位总宽度为短花针茅>无芒隐子草>碱韭。这反映出建群种和优势种在不同试验处理区具有不同的资源占有能力,受放牧家畜的影像,建群种短花针茅生态位宽度增加,而碱韭的的生态位宽度明显下降,表明放牧可使短花针茅植物种群泛化种特征表现明显,而碱韭则由于生态位宽度变窄趋于特化种;无芒隐子草生态位总宽度变化不大,说明无芒隐子草对放牧家畜这一生物因素影响的耐受性比较强。
2.2 植物种群在不同样地范围的生态位宽度变化
由图3可知,在CK处理区内,建群种和优势种生态位宽度随样地范围的增大变化趋势不同,短花针茅略称横向的“S”型变化趋势,无芒隐子草呈弱的下降的变化趋势,碱韭呈对数曲线的变化趋势;但在样地范围大于6300 m2范围后,建群种和优势种生态位宽度趋于稳定,此时的样方数为80个,表明如果要获得CK处理内建群种和优势种比较真实的生态位宽度,其资源状态样点数应该大于等于80个。在CG处理内,将3个植物种群生态位宽度随样地范围增大的变化程度看,在样地范围大于6300 m2范围后,植物种群的生态位宽度也呈基本稳定状态。因此,短花针茅荒漠草原无论放牧与否,其生态位宽度取样的样点数应该为80个或以上。
对比CK和CG处理可以看到,受放牧家畜的影响(样地范围大于等于6300 m2),建群种和优势种生态位宽度的变化规律不一致。在CK处理区内,3个植物种群生态位宽度表现为短花针茅<无芒隐子草<碱韭,而CG处理区内短花针茅>无芒隐子草>碱韭,这一结果与图2中的结果一致。这说明,当采用多样带计算生态位总宽度与采用大样地多样点计算结果所表现的趋势是一致的。对比CK处理,CG处理由于存在放牧家畜干扰,建群种和优势种的生态位宽度发生变化,这一变化与植物种群对生物、非生物环境影响的耐受性有关,短花针茅和碱韭植物种群对外界环境的耐受性较小,适应外界环境变化主要依靠大幅度调节生态位宽度来实现(变化幅度分别为0.429~0.813和0.235~0.828),而无芒隐子草对外界环境的耐受性较大,其主要依靠维持稳定的生态位宽度来抵御外界环境的影响。
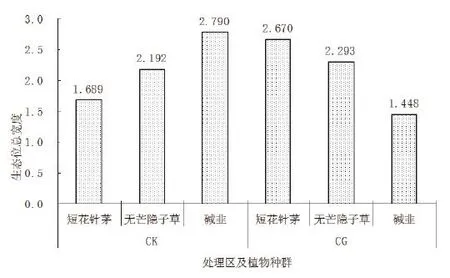
图2 主要植物种群生态位总宽度对放牧的响应Fig. 2 Effect of grazing on total niche width of main populations

图3 不同样地范围主要植物种群生态位宽度变化Fig. 3 The niche width changes of main populations in different plot
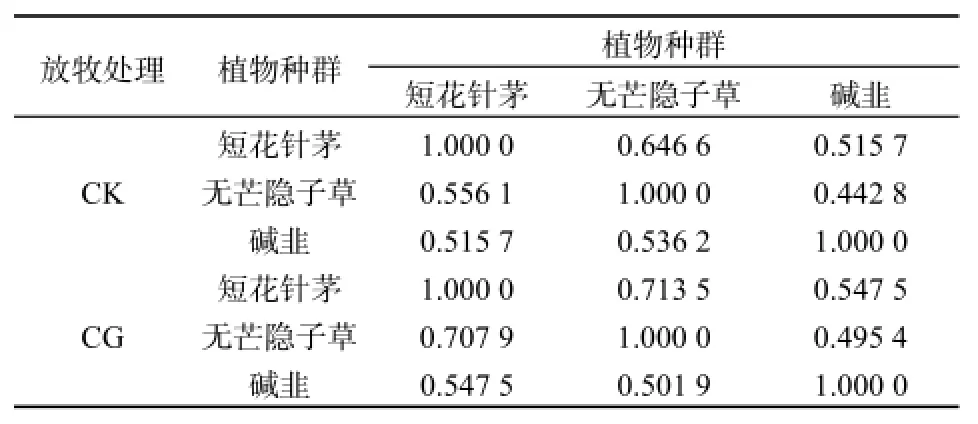
表3 不同样地范围主要植物种群生态位宽度关联情况Table 3 The grey correlation of niche width of main plant populations in different sample areas
尽管不同样地范围内主要植物种群生态位宽度指数变化趋势存在差异,但他们之间存在明显的关联性(见表3)。在CK条件下,短花针茅与无芒隐子草的关联度大于无芒隐子草与短花针茅的关联度,分别为0.6466和0.5561;短花针茅和碱韭之间的相互关联度相等,均为0.5157;无芒隐子草与碱韭的关联度小于碱韭与无芒隐子草的关联度,分别为0.4428和0.5326。在CG条件下,短花针茅与无芒隐子草的关联度大于无芒隐子草与短花针茅的关联度,分别为0.7135和0.7079,表明自由放牧没有改变种对间相互依赖的不对称性,但关联程度明显增大,其种间关系表现更为密切;短花针茅和碱韭之间的相互关联度相等,均为0.5475,这显示出自由放牧促进了短花针茅和碱韭的亲和性,导致其关联程度增大;无芒隐子草与碱韭的关联度小于碱韭与无芒隐子草的关联度,分别为0.4954和0.5019,说明自由放牧使得无芒隐子草与碱韭关联程度增大,而碱韭与无芒隐子草的关联程度减小,所以自由放牧对其相对依赖性的影响存在差别。
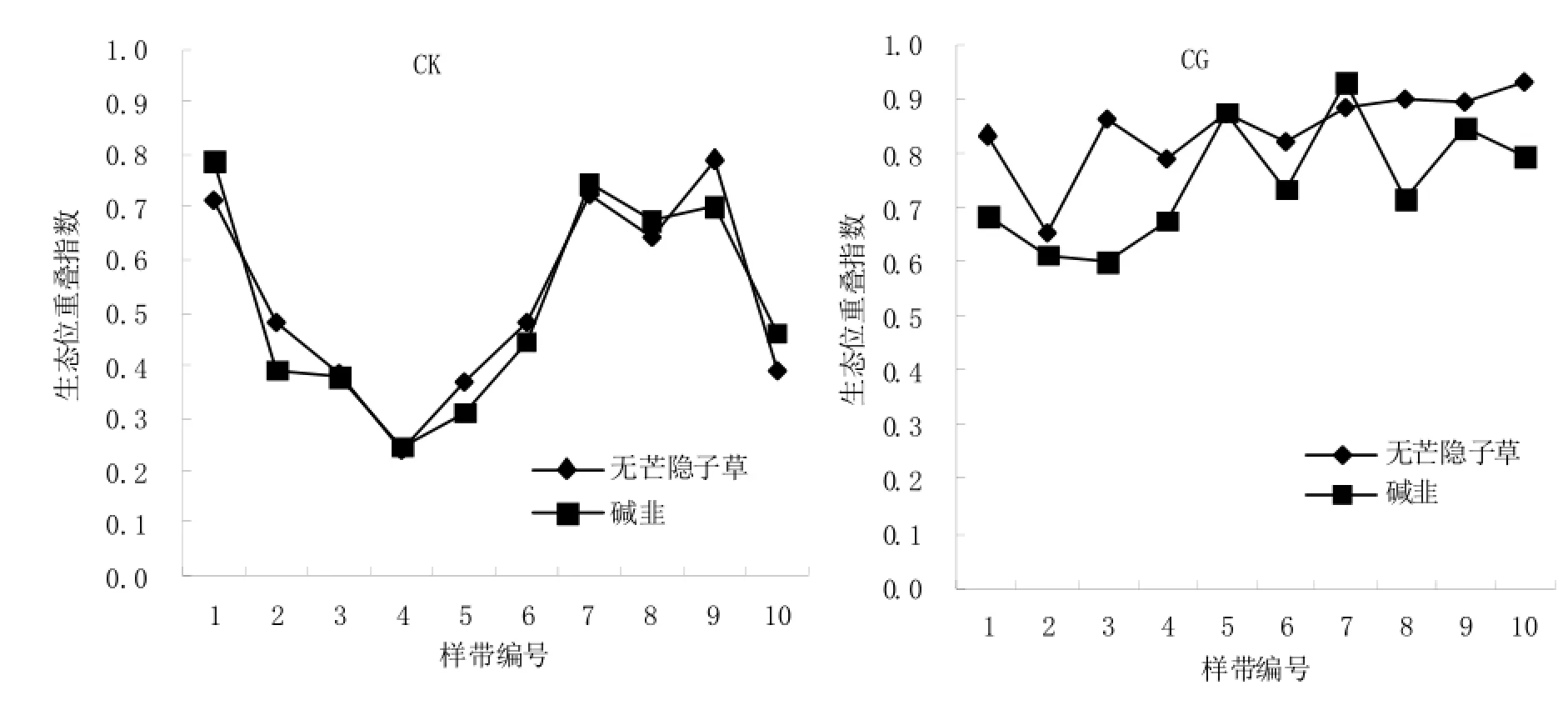
图4 短花针茅与无芒隐子草和碱韭的生态位重叠情况Fig. 4 The niche overlap of Stipa breviflora with Cleistogenes songorica and Allium polyrrhizum
2.3 植物种群在不同样带的生态位重叠程度
不同样带上短花针茅植物种群与无芒隐子草和碱韭的生态位重叠情况见图4,在CK条件下,短花针茅与无芒隐子草和碱韭的生态位重叠程度随样带的变化趋势与短花针茅针茅植物种群的生态位宽度表现基本一致,而CG处理内短花针茅与无芒隐子草和碱韭的生态位重叠程度随样带的变化不同于短花针茅针茅植物种群生态位宽度变化。这说明在CK条件下,建群种短花针茅植物种群与优势种无芒隐子草和碱韭的生态位重叠情况主要受短花针茅植物种群的资源利用情况决定;当存在放牧家畜干扰,建群种与两优势种的生态位重叠除受短花针茅植物种群的资源利用情况决定外,家畜的选择性采食和践踏等牧食行为也会对其生态位重叠情况产生明显影响。
结合短花针茅与无芒隐子草和碱韭的关联度(表2)可知,其种间相对关联程度影响着其生态位重叠程度,即CK条件下短花针茅与无芒隐子草和碱韭的关联程度接近,其生态位重叠程度也比较接近;CG条件下短花针茅与无芒隐子草的关联度大于其与碱韭的关联度,他们之间的生态位重叠程度表现亦是如此。因此,短花针茅植物种群生态位宽度变化趋势影响着其与优势种生态位重叠变化趋势,关联程度的大小影响着生态位重叠程度。
不同样带上无芒隐子草与短花针茅和碱韭的生态位重叠情况见图5,在CK处理内,无芒隐子草与短花针茅和碱韭的生态位重叠情况与无芒隐子草生态位宽度变化趋势基本一致,且无芒隐子草与其他两植物种群生态位重叠程度基本相同。在CG处理内,无芒隐子草与短花针茅和碱韭的生态位重叠情况不完全由无芒隐子草生态位宽度决定,放牧家畜的影响使得无芒隐子草与其他两植物种群的生态位重叠情况变得错综复杂。根据关联度分析(表2)结果可知,无芒隐子草与其他两植物种群的关联程度影响着其生态位重叠程度。因此无芒隐子草与其他植物种群的生态位重叠变花同样受本身的生态位宽度和他们之间的关联程度影响。
不同样带上碱韭与短花针茅和无芒隐子草之间生态位重叠情况无论是在CK处理区内还是在CG处理区内,其生态位重叠程度情况均比较复杂(图6);且CK处理区内碱韭与短花针茅和无芒隐子草的重叠程度总体上高于CG处理。根据关联度分析结果可知,CK区的碱韭与短花针茅和无芒隐子草的关联度大于CG区,这表明其种间关联程度与生态位重叠程度存在正相关。
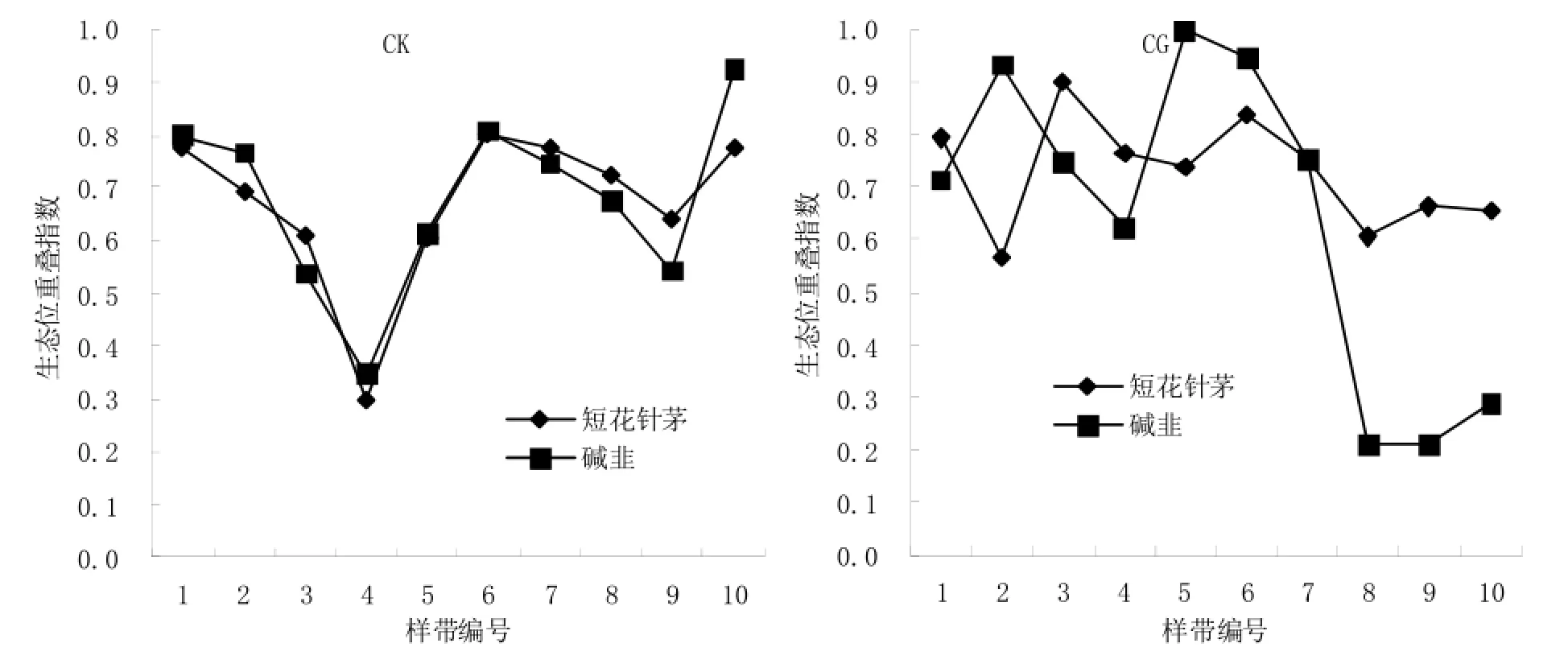
图5 无芒隐子草与短花针茅和碱韭的生态位重叠情况Fig. 5 The niche overlap of Cleistogenes songorica with Stipa breviflora and Allium polyrrhizum
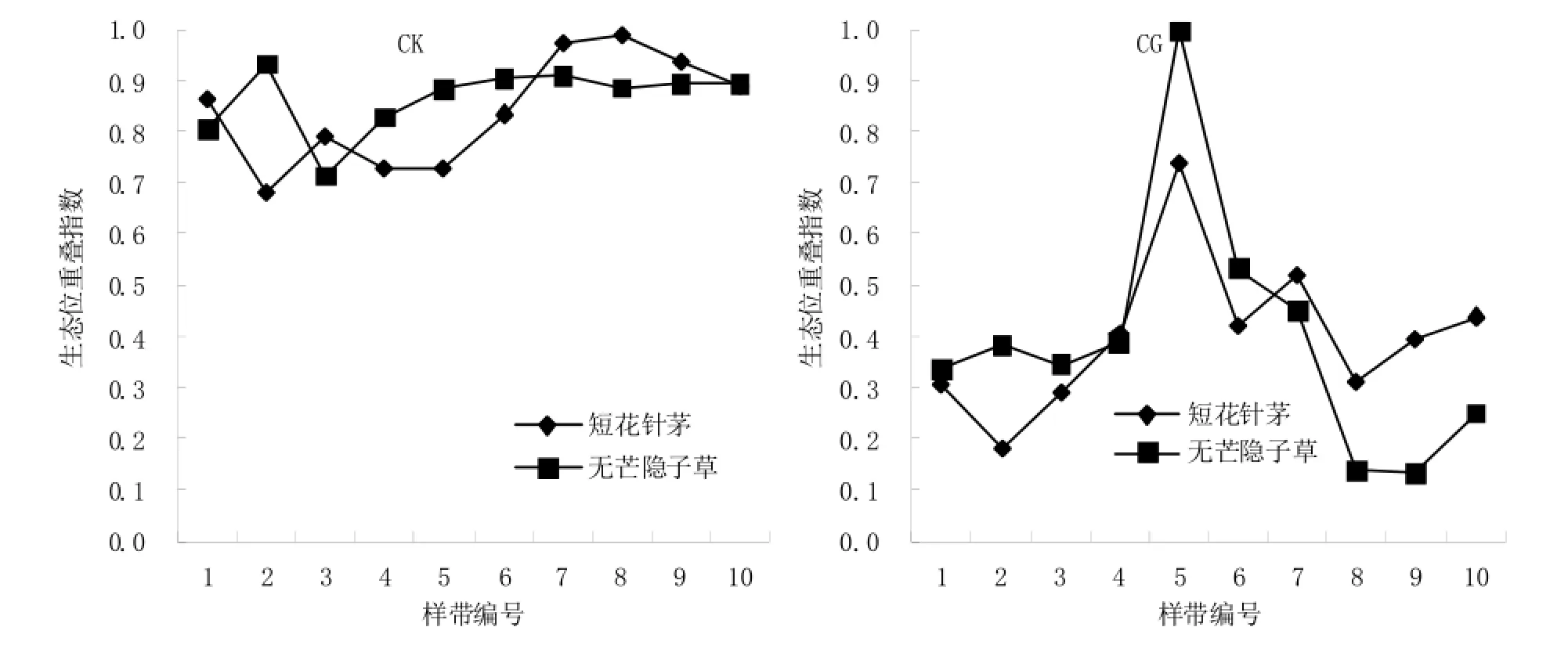
图6 碱韭与短花针茅和无芒隐子草的生态位重叠情况Fig. 6 The niche overlap of Allium polyrrhizum with Stipa breviflora and Cleistogenes songorica
综合图4~图6和表2来看,在3个植物种群种对之间的重叠程度存在差异,这种差异首先是由于植物种群占有资源能力不同导致了生态位宽度不同,进而影响其生态位重叠程度发生变化,同时植物种群间的相互关联性差异也对其生态位重叠程度产生影响。
2.4 植物种群在不同样地范围的生态位重叠程度
在CK处理区内,短花针茅植物种群与无芒隐子草和碱韭的生态位重叠程度随着样地范围的增加呈现先降低后增加并趋于稳定的变化趋势(如图7所示);在CG处理区内,短花针茅与无芒隐子草和碱韭的生态位重叠程度基本维持在某一相对稳定的水平。受放牧家畜的干扰,CG处理内短花针茅与无芒隐子草和碱韭的生态位重叠程度相对CK处理增大,这表明短花针茅可以通过增加与无芒隐子草和碱韭的生态位重叠程度来抵抗家畜牧食行为的干扰,短花针茅与无芒隐子草和碱韭的种间竞争关系减弱,种间的亲和性增强,表3中种间关联度分析证实了这一研究结果。
在CK处理区和CG处理区内,短花针茅与无芒隐子草的生态位重叠程度总体上均大于短花针茅与碱韭的生态位重叠程度,说明在短花针茅荒漠草原区,无论放牧与否,短花针茅与碱韭的之间的种间竞争强度大于短花针茅与无芒隐子草的种间竞争强度,这与种间关联度分析结果(表3)相一致。
在CK处理区内(见图8),无芒隐子草与短花针茅和碱韭的生态位重叠程度随样地面积的增大呈现先减小后稳定的变化趋势,在CG处理区内,无芒隐子草与短花针茅之间的生态位重叠程度基本稳定在0.7~0.8之间,略呈下降的变化趋势,而无芒隐子草与碱韭的生态位重叠程度波动情况比较复杂,这一复杂程度与无芒隐子草生态位宽度变化密不可分。
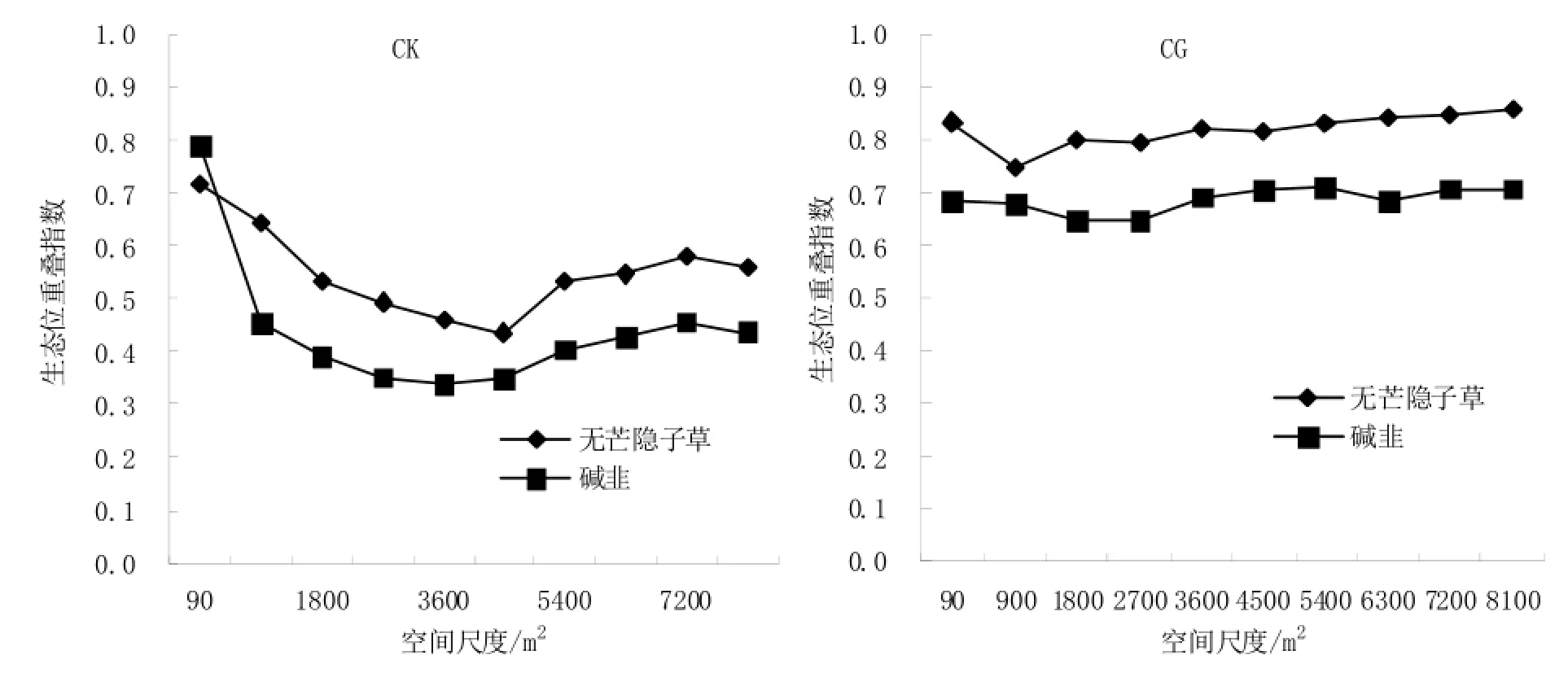
图7 短花针茅与无芒隐子草和碱韭的生态位重叠情况Fig. 7 The niche overlap of Stipa breviflora with Cleistogenes songorica and Allium polyrrhizum
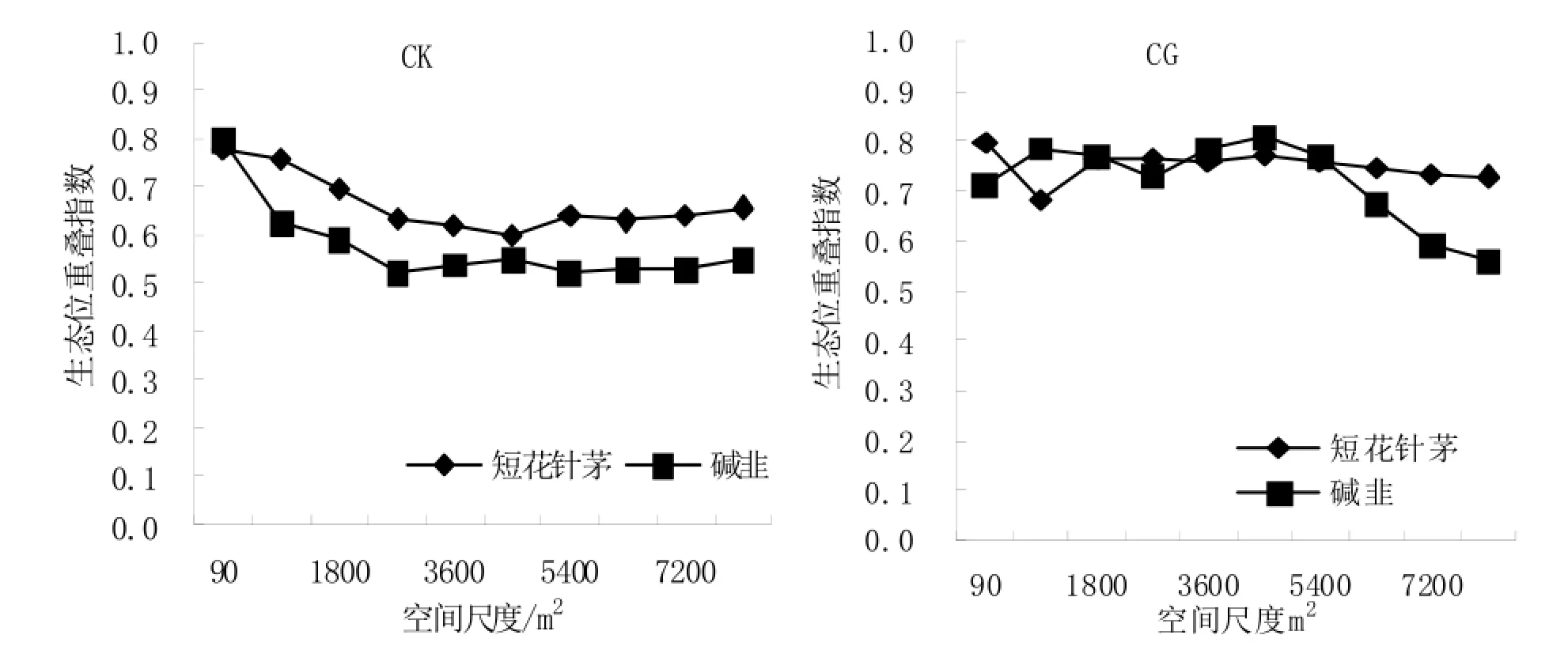
图8 无芒隐子草与短花针茅和碱韭的生态位重叠情况Fig. 8 The niche overlap of Cleistogenes songorica with Stipa breviflora and Allium polyrrhizum
综合表3分析结果来看,受放牧家畜影响后,无芒隐子草与短花针茅和碱韭的关联程度增加,生态位重叠程度总体也呈增加的变化趋势,说明无芒隐子草与短花针茅植物种群一样,通过减少与其他植物种群竞争关系并增加与其他植物种群的生态位重叠程度来抵御放牧家畜的干扰。
在CK处理区内,碱韭与短花针茅和无芒因子草之间的生态位重叠程度随着样地面积的增加呈现先减小后增加并逐渐趋于稳定的变化趋势,且碱韭与短花针茅植物种群之间的生态位重叠程度小于碱韭与无芒隐子草的生态位重叠程度(见图9),说明CK区内碱韭与短花针茅植物种群之间的竞争强度弱于碱韭与无芒隐子草之间的竞争强度;CG区亦是如此。受放牧家畜的影响,碱韭植物种群与其他两植物种群的重叠程度下降,表明放牧导致碱韭与其他两植物种群的种间竞争作用增强,根据生态位宽度可知,竞争结果导致碱韭生态位宽度大幅度下降。
综合图7~图9和表3可以看到,植物种群生态位重叠程度随着样地面积的增大变化规律各不相同,但当样地面积大于等于6300 m2以后,3个植物种群间生态位重叠程度趋于稳定。生态位重叠程度不但与植物种群生态位宽度有关,与种对间种间关联度也存在直接关系。
3 讨论
3.1 植物种群生态位宽度与其生态适应性和分布幅度
植物种群生态位宽度揭示了其对资源的利用能力,表征了其生态适应性和分布幅度(奚为民,1993;李登武等,2006),本研究对短花针茅荒漠草原建群种和优势种的生态位宽度进行了分析,结果证实了3个植物种群具有较强的生态适应性,因而其资源利用能力强、生存机会多,分布范围广。植物种群生态宽度在反应种群生态适应性时,除了体现在资源利用能力和分布范围上,还体现在生态位调控能力上,如短花针茅和碱韭植物种群主要依靠大幅度调节生态位宽度来适应放牧干扰,而无芒隐子草主要是维持生态位宽度相对稳定来适应放牧干扰。在资源状态相同的情况,不同植物种群受生物和非生物因素同时影响,生态位宽度可能相同,也可能不同(张金屯,2011),本研究结果(同一样带不同植物种群生态位宽度)支持这一观点。
3.2 生态位重叠程度与种对间的竞争强度及亲和关系
植物种群生态位变化与其所处空间的资源状密不可分,且同种间竞争紧密联系在一起(Silvertown,1983;Abrams,1987;汪殿蓓等,2005)。种群间生态位重叠程度较大,种群间的竞争作用比较强烈,反之种群间竞争作用比较小,原因是当空间资源状态有限时,生态位重叠程度大的植物种群因资源限制而发生竞争作用,竞争作用的结果是导致生态位分离,种间生态位重叠程度变小(Hardin,1960;张金屯,2011)。本研究结果显示,不同植物种群由于本身的植物学特征和生物学特性差异,受家畜放牧影响,种群生态位重叠程度增加,并不是增加种间的竞争作用,而是种群间通过增加生态位重叠程度来保证物种存活和相对稳定的生物量来抵御放牧的影响。因此,单一的从生态位重叠程度大小来判断种间竞争强度需要谨慎考虑,国外越来越多的研究证明,种群间的关系不但存在竞争关系(Silvertown,1983;Abrams,1987),也存在亲和关系(Gross,2008;Martorell和Freckleton,2014),且在一定程度上可以相互转化(Zavala等,2005;Martorell和Freckleton,2014),因此我们认为放牧导致无芒隐子草和短花针茅与其他植物种群间生态位重叠程度变大,是由于亲和关系增强共同抵御外界环境干扰的一种响应;碱韭与其他两植物种群生态位重叠程度下降是由于放牧干扰导致其亲和作用下降,抵御外界环境干扰的能力下降。

图9 碱韭与短花针茅和无芒隐子草的生态位重叠情况Fig. 9 The niche overlap of Allium polyrrhizum with Stipa breviflora and Cleistogenes songorica
3.3 生态位重叠的不对称性与种间关系的不对称性
种间竞争关系存在不对称性,这已经得到国外研究者的证实(Weiner,1990;Connolly和Wayne,1996;Freckleton和Watkinson,2001)。通过本研究的种群生态位重叠情况可以看到,生态位重叠程度在种对间存在差异,植物种群竞争能力也相应地存在差别;由于生态位重叠程度可解释为种间竞争作用或反映亲和作用大小,因此,生态位重叠反映的种群间亲和关系也存在不对称性,且这种不对称性的表现是由于种群本身占有资源的能力决定的,种对间植物种群占有资源能力存在差异会导致其竞争能力或亲和能力产生不对称性,这一研究结果与希吉日塔娜等(希吉日塔娜等,2013)的研究结果相一致,他们的关系可用图10表示。

图10 植物种群Levins生态位重叠图示Fig. 10 Levins niche overlap of plant populations
3.4 资源异质和外界干扰对种群生态位及群落特征的影响
由于生态位与其所处空间的资源状态密不可分(汪殿蓓等,2005),因此,讨论植物群落种群间生态位变化情况,应该考虑空间资源异质性影响,空间取样范围和取样的样点数都应该考虑。所以在研究生态位宽度的时候,应该首先分析种群生态位宽度和重叠程度随取样范围或样点数的变化规律,并在其相对稳定状态分析其变化特点和规律,以便使分析结果能够代表草地植物种群生态位真实情况,而不是局部资源空间的种群生态位表现。同时,由于3个植物种群数量消涨、时空变化及结构的位移均会引起群落的巨大波动(卫智军等,2013),使得短花针茅荒漠草原在不同季节、不同年度受不同干扰因素、干扰强度影响均会表现出不同的群落水平结构,总体上,2012年8月份CK处理区植物种群数量碱韭>无芒隐子草>短花针茅,CG处理区反之;受草地土壤养分资源异质性和其他植物群落数量消涨变化影响,3个植物种群水平结构的数量变化存在差异,形成短花针茅荒漠草原特有的群落类型和景观特征。
4 结论
1)样地范围大于或等于6300 m2(样点数大于或等于80),植物种群生态位宽度和重叠情况趋于稳定,在此取样条件下研究短花针茅荒漠草原建群种和优势种生态位变化特点和规律具有代表性意义。
2)放牧使短花针茅植物种群生态位宽度得到大幅度提升,而碱韭生态位宽度大幅度下降;无芒隐子草受放牧影响生态位宽度变化不大。
3)CG处理区短花针茅、无芒隐子草植物种群分别与其他两植物种群生态位重叠程度大于CK处理,碱韭植物种群与其他两植物种群生态位重叠程度小于CK处理。
ABRAMS P A. 1987. Alternative models of character displacement and niche shift. I. Adaptive shifts in resource use when there is competition for nutritionally nonsubstitutable resources[J]. Evolution, 41(3): 651-661.
CONNOLLY J, WAYNE P. 1996. Asymmetric competition between plant species[J]. Oecologia, 108(2): 311-320.
FRECKLETON R P, WATKINSON A R. 2001. Asymmetric competition between plant species[J]. Functional Ecology, 15(5): 615-623.
GROSS K. 2008. Positive interactions among competitors can produce species‐rich communities. Ecology Letters, 11(9): 929-936.
HARDIN G. 1960. The competitive exclusion principle [J]. Science, 131(3409): 1292-1297.
MARTORELL C, FRECKLETON R P. 2014. Testing the roles of competition, facilitation and stochasticity on community structure in a species‐rich assemblage [J]. Journal of Ecology, 102(1): 74-85.
SILVERTOWN J W. 1983. The distribution of plants in limestone pavement: tests of species interaction and niche separation against null hypotheses[J]. The Journal of Ecology, 71(3): 819-828.
WEINER J. 1990. Asymmetric competition in plant populations[J]. Trends in Ecology & Evolution, 5(11): 360-364.
ZAVALA M A, BRAVO DE LA PARRA R. 2005. A mechanistic model of tree competition and facilitation for Mediterranean forests: scaling from leaf physiology to stand dynamics[J]. Ecological Modelling, 188(1): 76-92.
董全民, 赵新全, 马玉寿. 2007. 放牧率对高寒混播草地主要植物种群生态位的影响[J]. 中国生态农业学报, 15(5):1-6.
付为国, 李萍萍, 卞新民, 等. 2008. 镇江内江湿地植物群落演替进程中种群生态位动态[J]. 生态环境, 17(1): 278-284.
李登武, 张文辉, 任争争. 2006. 黄土沟壑区狼牙刺群落优势种群生态位研究[J]. 应用生态学报, 16(12): 2231-2235.
李军玲, 张金屯, 郭逍宇. 2003. 关帝山亚高山灌丛草甸群落优势种群的生态位研究[J]. 西北植物学报, 23(12): 2081-2088.
刘贵河,王国杰,汪诗平,等. 2013. 内蒙古荒漠草原主要草食动物食性及其营养生态位[J]. 生态学报, 33(3): 856-866.
刘红梅. 2011. 短花针茅草原群落特征与草地空间异质性对不同放牧制度的响应[D]. 呼和浩特: 内蒙古农业大学: 48-58.
牛克昌, 刘怿宁, 沈泽昊, 等. 2009. 群落构建的中性理论和生态位理论[J]. 生物多样性, 17(6): 579-593.
苏鹏飞, 张克斌, 王瑞斌, 等. 2012. 人工封育区沙化草地植被生态位研究[J]. 生态环境学报, 21(3): 422-427.
汪殿蓓, 暨淑仪, 陈飞鹏, 等. 2005. 不同水平的仙湖苏铁种群在土壤资源上的生态位宽度[J]. 生态环境, 14(6): 913-916
王仁忠. 1997. 放牧影响下羊草草地主要植物种群生态位宽度与生态位重叠的研究[J]. 植物生态学报, 21(4): 304-311.
卫智军, 韩国栋, 赵钢, 等. 2013. 中国荒漠草原生态系统研究[M]. 北京: 科学出版社.
希吉日塔娜, 吕世杰, 卫智军, 等. 2013. 不同放牧制度下短花针茅荒漠草原植物种群作用和种间关系分析[J]. 生态环境学报, 22(6): 976-982.
奚为民. 1993. 怀柔山区灌丛群落优势种群生态位的研究[J]. 植物生态学与地植物学学报, 17(4): 324-330.
张金屯. 2011. 数量生态学[M]. 第二版. 北京: 科学出版社, 113-123.
Effect of Grazing on Niche of Constructive and Dominant Species in the Desert Steppe
LV Shijie, LIU Hongmei, WU Yanling, WEI Zhijun, SUN Shixian, WANG Yingjie, LIN Sen
1. College of Science, Inner Mongolia Agricultural University, Huhhot 010018, China;
2. College of Ecology and Environmental Science, Inner Mongolia Agricultural University, Huhhot 010019, China 3. Academy of Forestry Sciences, Huhhot 010010, China; 4. College of Life Science and Technology, Inner Mongolia Normal University, Huhhot 010022, China
Stipa breviflora was the constructive specie, and Cleistogenes songorica and Allium polyrrhizum were dominant species of Stipa breviflora desert steppe. Their aboveground present biomass reached to 60%~80% of the total biomass, of which quantities, time and space changes, and structure displacement causeed large fluctuations in communities. Niche breadth and niche overlap index were analyzed to make sure the capacity of resources consumption, the relationships and variation impacts by grazing in Stipa breviflora desert steppe in Inner Mongolia Saihantala research station,established in 1999. The samples were collected using the transect and plot method in contrast area (CK) and continuous grazing area (CG), in August 2012. The results were shown as follows. Stipa breviflora and Allium polyrrhizum affected by grazing mainly adjusted niche width to ensure plant populations survival and status or roles in the community, and their variation range were 0.429 to 0.813 and 0.235 to 0.828, respectively. Cleistogenes songorica did not change in niche width (0.556 to 0.761), and showed a strong resistance to grazing. Stipa breviflora and Cleistogenes songorica increased niche overlap extent with the other two populations due to grazing, whereas Allium polyrrhizum was the opposite. There was a relationship between niche overlap extent and populations competitive ability, and also linked with the populations affinity that exists asymmetry.
Stipa breviflora desert steppe, constructive specie, dominant specie, ecological width, niche overlap
Q948
A
1674-5906(2014)10-1600-10
吕世杰,刘红梅,吴艳玲,卫智军,孙世贤,王颖杰,林森. 放牧对荒漠草原建群种和优势种生态位的影响[J]. 生态环境学报, 2014, 23(10): 1600-1609.
LV Shijie, LIU Hongmei, WU Yanling, WEI Zhijun, SUN Shixian, WANG Yingjie, LIN Sen. Effect of grazing on niche of constructive and dominant species in the desert steppe [J]. Ecology and Environmental Sciences, 2014, 23(10): 1600-1609.
公益性行业(农业)科研专项(201003019);国家重点基础研究计划(973计划)项目(2014CB138800);科技部“十二五”农村领域国家科技计划课题(2012BAD13B00)
吕世杰(1978年生),男,讲师,主要从事生物统计和数量生态方面的研究。E-mail:lshj123@163.com
*通信作者:nmndwzj@163.com
2014-07-22
