端粒和端粒酶与癌症发生和治疗的关系研究进展
2014-01-02陈海伟杨静茹
陈海伟,杨静茹
(赤峰学院 生命科学学院,内蒙古 赤峰 024000)
端粒、端粒酶与肿瘤的研究一直以来都是科学家研究的热点,在2002年到2004年达到了高峰时期.2009年,美国科学家伊丽莎白·布莱克本、卡萝尔·格雷德和杰克·绍斯塔克三人因发现了端粒和端粒酶保护染色体这一机制获得了诺贝尔生理学或医学奖,更加引起了研究者们的关注[1].
机体受环境条件或自身的影响会发生一些遗传或表观遗传信息的改变,这些改变很多情况下会导致癌变,这是由于它们的改变占用了一些信号通路,多条信号通路的综合改变使有潜在癌变倾向的细胞无限制生长从而导致癌变,进一步扩散,最终导致机体死亡.癌基因是高度重新排列的基因,被赋予了复杂的转移性,还有局部复制而导致细胞数目改变的特征,这些改变是通过与癌症相关的基因的改变来实现的.一直以来,人们努力地去寻找造成癌基因不稳定的根本机制,最终揭示了在癌症发生中具有突出作用的重要结构——端粒,端粒是一种核蛋白结构,可以保护真核细胞线性染色体末端,由于真核细胞的染色体复制机制的原因,其末端在DNA复制中逐渐缩短,使得依赖细胞分裂的组织的更新令染色体的稳定性下降.很多相关研究证明了端粒缩短对组织衰老和过度增生性疾病的作用,表明端粒的状态与癌变风险密切相关.在大多数晚期癌症患者细胞中,端粒酶被激活进而维持端粒长度,最新的数据显示,端粒酶具有直接控制促癌通路的能力.本文综述了端粒和端粒酶在正常组织原代细胞中的作用以及在癌症的发展中所扮演的角色.
1 概述
1.1 端粒(Telomere)
1.1.1 概念
端粒是真核生物染色体(线性)末端的特殊结构,由富含鸟嘌呤G的DNA重复序列以其相关蛋白组成,是DNA蛋白复合物,对染色体结构的稳定以及细胞的正常有丝分裂功能的实现有非常重要的意义.
1.1.2 结构
端粒是由富含鸟嘌呤核苷酸的DNA重复序列构成,可作为大型蛋白的结合位点(见图1).在脊椎动物中,端粒由重复序列TTAGGG组成,包括双链部分以及单链部分,双链部分较长,单链端粒的3′端突出,称为3′悬突[2].单链端粒高度保守,末端可形成一个特殊的环状结构,称为t-loop,以此来固定染色体末端,保持其稳定性[3].双链的端粒序列可以特异性的结合双链端粒DNA结合蛋白,即端粒重复序列结合因子1(telomeric repeat binding factor 1,TRF1)和端粒重复序列结合因子2(telomeric repeat binding factor 2,TRF2).TRF2在端粒末端保护中起关键的作用,可以促进t-loop环的形成,是端粒长度的负性调节因子,通过抑制TRF2显性基因的过表达或者敲除此基因,可造成保护性的帽子结构缺失,表现为3'悬端的处理以及染色体末端的连接反应[4,5].TRF1不仅起到了调节端粒长度的作用,也可以通过调节端粒重复序列来促进DNA的复制[6,7,8].TRF1和TRF2专一性的与其他相 关 蛋白 TIN2(TRF1-interacting nuclear factor)、POT1(protection of telomeres 1)、TPP1和 RAP1(Repressor Activator Protein 1)相互作用,构成一个复合体共同调节端粒长度及活性[9].
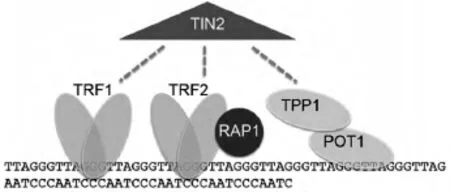
图1 端粒保护染色体末端
1.2 端粒酶(Telomerase)
1.2.1 概念
端粒酶又称端粒末端转移酶,其本质为核糖核蛋白.人体正常细胞中,端粒随着细胞有丝分裂代数的增加而逐渐缩短,每分裂一次约缩短50—200bp,到达临界水平时,细胞进入衰老期,停止继续分裂,但一些特殊细胞,比如绝大多数的癌细胞和永生化细胞,可通过复制维持其端粒长度,其催化不是常规的DNA聚合酶完成,只能由特殊的逆转录酶也就是端粒酶以自身RNA为模板来完成端粒的延长.
1.2.2 结构
目前对于端粒酶的研究大部分针对动物细胞,常用材料有小鼠、人类等,普遍认为,人类端粒酶主要包括三种成分:端粒酶RNA组分(Telomerase RNA Component,TERC)、端粒酶催化亚单位 (Telomere reverse transcriptase,TERT,端粒逆转录酶)和端粒酶相关蛋白 (telomerase associated proteinl,TP1).端粒酶是一种大型多亚基RNA酶,TERT和TERC构成催化中心[10,11,12,13](见图2).
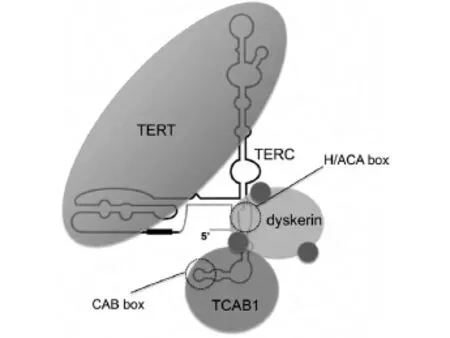
图2 端粒酶结构
在染色体末端,TERC作为逆转录模板使端粒重复序列延长.在体外TERT和TERC就足以使得端粒酶激活,而人体内,还需要其它酶的共同作用.TERC的5'端包含模板区,3'端包含两个序列作为其他端粒蛋白的结合位点.H/ACA box是蛋白dyskerin的结合位点[14,15,16],dyskerin对于端粒酶的组装和TERC的稳定性相当重要.Dyskerin还有三个相关蛋白,分别为NHP2、NOP10和GAR1.TCAB1是一种WD40重复序列蛋白质,在TERC中识别CAB box[17,18].以上两个蛋白相互作用去调节端粒酶的运行进而维持端粒稳定性.
1.3 端粒和端粒酶的主要功能及调节
1.3.1 端粒和端粒酶的功能
在近年来的研究中,研究者们一直致力于运用各种不同的模式生物进行实验,试图阐述端粒的组成结构,进而确定了端粒的主要功能.经过实验得出端粒功能主要有:保护染色体的线性末端,防止其降解和不必要的融合;运用其自身的结构能招募端粒酶,用来修复细胞分裂过程中出现的染色体末端缺失的重复序列;在细胞核中端粒能定位染色体末端,在减数分裂过程中这一作用至关重要,起到确保非错误性降解和等量的分裂的作用.相对哺乳动物和酵母的端粒的研究,关于植物端粒方面的研究还非常有限[19].
1.3.2 端粒酶调节端粒长度的机制
多数人体正常组织或者细胞不会有端粒酶的表达活性,一些有增殖潜能的细胞有低水平表达的端粒酶,如生殖细胞、造血干细胞、免疫记忆细胞、皮肤细胞等.以自身的RNA为模版,端粒酶能够逆转录合成具有DNA重复序列的端粒,从而使得端粒延长,保持染色体结构的相对恒定.
在端粒酶各个亚单位相互协作下可十分精确地调控端粒延伸的过程.首先,端粒酶逆转录酶TERT被激活,进而使端粒酶活化;其次,通过结合蛋白端粒酶结合于端粒,以端粒末端做为引物,与端粒酶RNA模板区的起始端结合,开始端粒DNA的合成,DNA链延长至模板区的末端时,延长的DNA末端与模板解开,新合成的端粒DNA仍然结合在端粒酶结合蛋白上,其末端又结合在端粒酶RNA模板区的起始位,如此反复进行,直至端粒达到适当长度时终止[20].
1.3.3 端粒酶活性的调节
对于端粒酶活性的调节方式,一般分为两种,一种是通过诱导端粒酶使其活性增强,目前是通过利用携带hTERT基因的载体转染干细胞的研究方法,从而激活端粒酶,使被转染的干细胞的端粒酶活性升高;另外一种是抑制端粒酶的活性,使端粒酶低水平的表达,甚至表达停止.研究发现,80%甚至以上的肿瘤细胞的端粒酶都处于活化状态.
在正常组织细胞中(包括正常干细胞),端粒酶只是暂时性表达并且表达水平很低,同肿瘤细胞相,比其端粒比较长,肿瘤细胞的端粒酶的活化是肿瘤发生的一个重要步骤,且与肿瘤细胞的增殖、分化、转移、凋亡以及细胞周期有关,这就使人们把研究癌症治疗药物的眼光落到抑制端粒酶活性上,使其实现成为可能.目前对于该类药物的研究方向主要有酶直接抑制剂、端粒酶主动免疫治疗和干扰端粒酶反转录过程等几个方向[21].
1.4 端粒酶的检测技术
随着端粒酶的结构、功能及调控的进一步研究,对端粒酶在细胞永生化以及肿瘤癌变进程中的作用有了崭新的认识,因此对端粒酶的检测方法的研究成为研究端粒酶至关重要的部分.现在主要的技术有:端粒重复序列延伸法(TEA),端粒重复序列扩增法(TRAP),半定量TRAP法,细针抽吸端粒酶重复扩增法(FNA—TRAP),银染端粒重复序列扩增法(SS—TRAP),接近闪烁分析端粒重复序列扩增法,酶联免疫吸附端粒重复序列扩增法(TRAP—ELISA),原位TRAP法应,定量端粒重复序列扩增(RTQ—TRAP)[22].
1.5 端粒异常是原代细胞缺陷的关键驱动者
通过敲除TERC基因组分的端粒酶的小鼠模型,研究端粒逐渐缩短引起的端粒危机对生物体的影响.出乎意料的是,前几代均表现正常,从而一段时期内科学家轻率的认为端粒酶组分对于生命是可有可无的[23].后来发现这些小鼠的存活是因实验小鼠品系有较长的端粒,然而,第一代(G1)TERC基因敲除纯合体小鼠的持续繁殖产生了新生的TERC基因敲除纯合体小鼠,导致端粒逐步丧失,最终引起了高度增生的严重组织变性,第4代出现生育能力下降,到G5–G6TERC基因敲除纯合体小鼠出现不育现象,在睾丸组织中表现出较高的细胞凋亡和生殖细胞耗竭,出现骨髓造血细胞功能受损以及淋巴细胞增殖减弱.在G5–G6TERC基因敲除纯合体组织中,端粒逐渐缩短的细胞遗传学证据很清晰,包括染色体末端融合以及有丝分裂后期的染色体桥的出现.说明这些病症以及其严重程度是与端粒功能失调的程度紧密联系的.
在较晚世代的端粒酶基因敲除的小鼠中,严重退行性的组织缺损似乎与增殖损伤、细胞凋亡加强的原代细胞以及干细胞中自我更新缺陷有关联.这些缺陷是显而易见的,即原代细胞凋亡率较高,造成肠道腺细胞绒毛萎缩,在端粒酶缺失的小鼠中造血干细胞潜在减少.与野生型相比,经连续辐射的G1TERT基因敲除纯合体或者G1TERC基因敲除纯合体小鼠的造血干细胞造血能力下降,这种缺陷在较晚世代TERT基因敲除纯合体小鼠造血干细胞中更容易出现,具有较短端粒的较晚世代的TERT基因敲除纯合体小鼠与长端粒的野生型小鼠杂交产生的TERT基因敲除杂合体小鼠拥有一半正常的端粒和一半功能失调的短端粒,这些小鼠显现出与原来较晚世代TERT基因敲除纯合体小鼠相似的细胞凋亡和增殖缺陷,所以研究者一直认为系列的端粒功能失调是原代细胞出现缺陷的关键驱动者,更加突出了端粒在细胞中的重要地位.
2 端粒、端粒酶与癌症的关系
2.1 正常细胞和癌细胞复制中端粒的差异性
在人类成纤维细胞原代培养实验中发现了端粒与癌细胞之间的联系,正常细胞表现出有限的复制能力,原代细胞一般分裂60–80代就进入衰老状态,直至死去,与此相反,癌细胞系在细胞培养时可以无限制的生长下去 (见图3).正常培养的人类成纤维细胞在分裂期间端粒是逐渐缩短的,这些缩短的端粒激活了细胞衰老程序,但是在肿瘤细胞中端粒却维持原始状态并不缩短.随着人成纤维细胞继续分裂,端粒逐渐缩短,端粒帽子结构功能损坏,导致染色体极度不稳定,最终细胞凋亡,这种状态称为危机.但是有少数细胞则可以通过一定的机制延长端粒而成为不受调控的细胞,比如人类成纤维细胞的衰老以及细胞出现的危机,可以通过重激活端粒酶进而合成端粒重复序列有效的化解,大部分癌症在发生时通过重新激活端粒酶的活性以维持端粒长度.
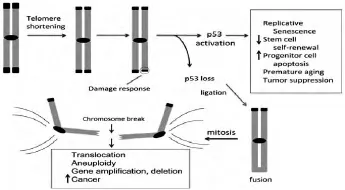
图3 端粒缩短激活p53并且通过基因扩增和缺失驱动上皮癌形成
2.2 端粒酶结构中TERT对其活性起关键作用
有学者研究了不同病变的胃黏膜端粒酶活性并对其亚单位进行检测,结果发现端粒酶活性均有不同程度的表达,而正常胃黏膜中则没有端粒酶的表达,且癌细胞中表达率最高.TERT主要在癌组织或部分癌前组织中表达,而其他两个亚基在所有情况下均有表达,TERT在癌组织中的表达与端粒酶活性有明显的相关性,由此提示在胃癌的发生过程中端粒酶可能具有极其重要的作用;可把TERT作为胃癌的诊断指标,同时也可以把TERT作为胃癌基因治疗的靶位点.张营等研究了在肝癌中端粒酶组分的表达情况发现,hTERTmRNA阳性表达普遍存在于肝癌组织中,说明了通过检测肝癌中hTERTmRNA表达情况可用来作为肝癌诊断及预后估计的新标志物.总之,这些研究结果说明对于端粒酶来说,其三种组分是缺一不可的,但是TERT只在有端粒酶活性的细胞中表达,相较于端粒酶其它两个组分来说,它的上调可能与端粒酶活性的增强有很大联系,从而稳定端粒长度,支持细胞无限的分裂.
2.3 端粒和端粒酶与P53的关系
2.3.1 端粒与P53蛋白通路及细胞衰老
端粒极度缩短诱导细胞和组织的改变,这个变化主要集中在p53抑癌蛋白这一细胞压力传感器上,它可以对癌基因的激活、DNA损伤信号和缺氧等各种刺激做出反应,起到强制细胞周期停止或使细胞凋亡的作用.实验证明,在端粒酶基因敲除的小鼠中,p53蛋白是稳定的,并且p53缺失显著削弱细胞周期阻滞、降低了细胞死亡率.在人工培养细胞的中,通过破坏端粒结合蛋白TRF2使得端粒帽子结构损坏,进而强烈的激活p53基因.以人类成纤维细胞中的端粒为例,端粒TRF2的功能被打乱,出现一些导致染色体内部双链断裂的典型DNA损伤蛋白,显示的病灶有组蛋白变体γH2AX、DNA损伤蛋白p53BP1、Mre11复杂化以及磷酸共济失调毛细血管扩张症突变(ataxia telangiectasia mutated,ATM)等,促进细胞衰亡.在端粒酶基因敲除小鼠繁殖的较晚世代中,表现出早衰综合症,包括毛发变得灰白无光泽,生存期缩短等不良反应,说明端粒缩短可能会引起许多老化现象,是重要的遗传毒性信号,这个信号可以激活DNA损伤信号,从而激活p53基因,加速细胞衰老进程;活性氧可以引起DNA损伤,进一步加强p53对DNA损伤检查点的反应,进一步加速端粒的侵蚀.
总之,这些数据证明在衰老的进程中,依赖于p53基因的DNA损伤信号的重要性,端粒功能失调会引起p53基因的表达,进而加速异常组织细胞的衰老或凋亡.
2.3.2 端粒缩短抑制肿瘤发生
为确定端粒耗损是否影响癌变过程,将TERC基因敲除纯合体与p53基因敲除纯合体小鼠进行杂交研究,结果发现,在早期阶段,端粒功能异常给细胞以及器官带来的不利影响可被p53基因敲除明显减弱.同代TREC和p53双基因敲除纯合体小鼠与只有TREC基因敲除纯合体的小鼠相比,前者存活时间长于后者,多数G5-G6 TREC和p53双基因敲除纯合体纯合体小鼠睾丸组织的凋亡被减弱,G6TREC和p53双基因敲除纯合体小鼠睾丸大小要比G6只有TREC基因敲除纯合体的小鼠的正常.由此说明,在TREC基因敲除纯合体小鼠中,抑制细胞生长和凋亡的反应可在p53的功能丧失的情况下完全丧失,导致器官衰亡消弱,与端粒相关的组织退化得到改善.虽然对G6TREC和p53双基因敲除纯合体小鼠与G6唯TREC基因敲除纯合体小鼠的睾丸进行比较发现后者更接近正常,但G6TREC和p53双基因敲除纯合体小鼠中约有一半小鼠的生殖小管表现出不同程度的萎缩,而且在G7-G8双基因敲除纯合体的小鼠中重新出现萎缩,由此表明小鼠体内还存在一些不依赖于p53信号介导的端粒功能异常反应.
Ink4a/Arf基因编码两个不同的肿瘤抑制蛋白:细胞周期蛋白依赖性激酶抑制剂p16和p53激活因子p19ARF,因子p19ARF对p53蛋白调节主要是通过阻断p53的降解,检测异常的细胞周期,致使p53稳定.Ink4a/Arf突变小鼠长出淋巴瘤与肉瘤,具有短潜伏期和高外显率;但是,在G4/G5TERC-/-Ink4a/Arf-/-的小鼠显示出降低肿瘤的发病率和增长潜伏期的功效,肿瘤的范围没有变化,显示出p53突变的组织中,端粒缺失对肿瘤有抑制作用,说明了在肿瘤发生过程中的依赖p53的端粒检查点的反应以及端粒维持的重要性.
以上实验表明,在肿瘤发生过程中依赖p53的端粒检查点的反应以及端粒维持的重要性,说明端粒缺陷会引起依赖p53的DNA损伤修复促进细胞衰亡,防止癌变的发生,而在p53缺陷可以显著削弱端粒失调所带来的细胞衰亡等不利现象.
3 端粒酶抑制剂——癌症抑制物质的研究成果
基于端粒酶在癌症中的重要作用,无数的研究者将端粒酶作为癌症的治疗点,对端粒酶抑制剂的研究,主要是有以下途径:(1)利用反义 hTR(human telomerase RNA,hTR)、反义寡核苷酸(Antisense oligonucleotides,ODN)、肽苷酸(peptide nucleic acid,PNAS)、锤头状核酶 (Hammerhead Ribozyme)等手段抑制端粒酶RNA的模板作用,进而抑制端粒酶活性.(2)对端粒酶蛋白组分的抑制剂:端粒酶蛋白抗体;蛋白激酶C(protein kinase C,PKC)调节剂;TERT抑制剂.(3)其他抑制剂.
在对治疗癌症的研究过程中发现了许多植物提取物对端粒酶有一定的作用,比如:目前已证实的植物生物碱传统中药苦参的主要生物碱之一苦参碱、喜树碱、小檗碱、长春碱、苦参碱、秋水仙碱、三尖杉酯碱等,小檗属植物的一种具有生物活性的成分小檗胺,白头翁根中分离提取的桦木酸衍生物2、3-羟基桦木酸,红纹马先蒿中所含的苯丙素苷类化台物毛蕊花苷,从我国和印度的传统民间草药珠子草的乙酸乙酯提取物中纯化得到的亚甲基双氧木质素,茶叶的主要活性成分茶多酚中的儿茶素类物质,从中国云南省所产的豆科藤本植物相思子的种子中分离得到的一种新的单体蛋白相思子蛋白P2,均有抑制端粒酶活性的作用.因而也可以作为癌症抑制的有效物质,但是仍然需要在临床中不断实验.
4 结论和展望
从一系列人类系统和小鼠模型的研究表明,端粒酶对调节端粒的长度有重要作用,端粒功能的完好对于细胞增殖是至关重要的.在端粒酶中,TERT是关键的亚基,在端粒酶活性中起主要作用.端粒功能失调在原代细胞出现缺陷中扮演着重要的角色.端粒、端粒酶通过与p53等抑癌基因的相互作用,证明了其在细胞衰亡和癌变中的关键地位,并激起了人们对端粒酶抑制剂的研究兴趣.
端粒与端粒酶对癌症的重要性已被确定,但是更具体的作用机制仍然不清晰,端粒酶抑制剂的研究大部分也处于实验阶段,继续端粒与端粒酶这个领域的研究将会得到更新、更重要的成果,例如它们如何调节原代细胞、组织、有机体的年龄以及癌基因是如何发展等方面探索.
〔1〕孔令平,汪华侨.端粒和端粒酶与衰老、癌症的潜在关系——2009年诺贝尔生理学或医学奖简介[J].自然杂志,第31卷.第6期.
〔2〕Palm,W.et al.(2008)How shelterin protects mammalian telomeres.Annu.Rev.Genet.,42,301–334.
〔3〕Griffith,J.D.et al.(1999)Mammalian telomeres end in a large duplex loop.Cell,97,503–514.
〔4〕van Steensel,B.et al.(1998)TRF2 protects human telomeres from end-toend fusions.Cell,92,401–413.
〔5〕Celli,G.B.et al.(2005)DNA processing is not required for ATMmediated telomere damage response after TRF2 deletion.Nat.Cell Biol.,7,712–718.
〔6〕van Steensel,B.et al.(1997)Control of telomere length by the human telomeric protein TRF1.Nature,385,740–743.
〔7〕Smogorzewska,A.et al.(2000)Control of human telomere length by TRF1 and TRF2.Mol.Cell.Biol.,20,1659–1668.
〔8〕Sfeir,A.et al.(2009)Mammalian telomeres resemble fragile sites and require TRF1 for efficient replication.Cell,138,90–103.
〔9〕de Lange,T.(2005)Shelterin:the protein complex that shapes and safeguards human telomeres.Genes Dev.,19,2100–2110.
〔10〕Greider,C.W.et al.(1989)A telomeric sequence in the RNA of Tetrahymena telomerase required for telomere repeat synthesis.Nature,337,331–337.
〔11〕Lingner,J.et al.(1997)Reverse transcriptase motifs in the catalytic subunit of telomerase.Science,276,561-567.
〔12〕Nakamura,T.M.et al.(1997)Telomerase catalytic subunit homologs from fission yeast and human.Science,277,955–959.
〔13〕Meyerson,M.et al.(1997)hEST2,the putative human telomerase catalytic subunit gene,isup-regulated in tumor cellsand during immortalization.Cell,90,785-795.
〔14〕Mitchell,J.R.et al.(1999)A telomerase component is defective in the human disease dyskeratosis congenita.Nature,402,551–555.
〔15〕Cohen,S.B.et al.(2007)Protein composition of catalytically active human telomerase from immortal cells.Science,315,1850–1853.
〔16〕Venteicher,A.S.et al.(2008)Identification of ATPases pontin and reptin as telomerase components essential for holoenzyme assembly.Cell,132,945–957.
〔17〕Venteicher,A.S.et al.(2009)A human telomerase holoenzyme protein required for Cajal body localization and telomere synthesis.Science,323,644–648.
〔18〕Tycowski,K.T.et al.(2009)A conserved WD40 protein binds the Cajal body localization signal of scaRNP particles.Mol.Cell,34,47–57.
〔19〕刘由页,宋涵,李凤兰,等.植物端粒与端粒酶研究进展[J].北京林业大学学报,2012(5).
〔20〕王联群,刘德伍.端粒、端粒酶与干细胞[J].中国组织工程研究与临床康复,2009(10).
〔21〕惠子健.端粒酶的研究进展[J].山西职工医学院学报,2010(1).
〔22〕洪日,陈红风.端粒酶检测技术新进展[J].临床和实验医学杂志,2007(11):163-164.
〔23〕Blasco,M.A.et al.(1997)Telomere shortening and tumor formation by mouse cells lacking telomerase RNA.Cell,91,25–34.
