中国沿海日本蟳地理群体的形态学变异研究
2013-10-21贾翠华韩志强
郑 伟,贾翠华,韩志强
(浙江海洋学院水产学院,浙江舟山 316022)
日本蟳Charybdis japonica属于梭子蟹科Portunidae、梭子蟹亚科Portuninae,蟳属Charybdis,俗称靠山红、石蟳仔、石蟹、岩蛑和海蟳。日本蟳属于广温广盐型分布种,在我国渤海、黄海、东海和南海广泛分布。它主要栖息于潮间带,属于沿岸定居性种类[1-5],是我国沿岸岩礁区最常见的蟹类之一。日本蟳为高蛋白、高脂肪食品,其肉质鲜美且营养丰富,肌肉含有17种氨基酸,且不饱和脂肪酸含量很高,具有很高的经济价值[6]。
目前对日本蟳的相关研究主要集中在生物学特性、营养成分组成及分子遗传等方面[7-11],有关日本蟳形态学的研究较少,仅有丁金强等[12]对中国沿海的莱州湾、大连、海州湾和象山湾4个日本蟳群体的形态差异进行了分析比较,认为不同海区的日本蟳群体间已经产生了一定程度的形态差异,但该研究采样群体较少,并且形态指标只测定了12项,部分附肢数据没有测定,可能对研究结果存在一定的影响。形态学方法作为传统的生物学测定方法在海洋生物的种群鉴定研究中得到广泛的应用。本研究采用单因子方差分析、聚类分析、主成分分析和判别分析四种多元分析方法对我国7个日本蟳群体的22项形态学差异进行综合分析,探讨日本蟳群体形态差异与地理距离之间的相关性,为日本蟳不同海区种群的鉴别研究提供基础资料,也为日本蟳资源保护和合理利用提供科学依据。
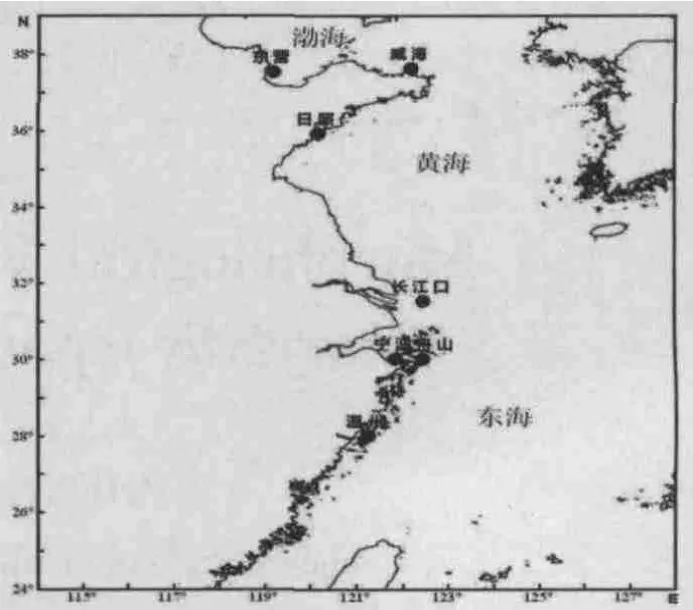
图1 日本蟳采样图Fig.1 Sampling locations of C.japonica
1 材料与方法
1.1 实验材料
本次实验采集了日本蟳东营、威海、日照、长江口、舟山、宁波和温州7个地理群体共154尾,各日本蟳群体采集的时间、地点及数量见表1。

表1 日本蟳各地理群体的采样信息Tab.1 Sample information of C.japonica populations
1.2 形态学参数的测定
通过参考高天翔等[13]和丁金强等[12]所测定的蟹类形态学参数,我们选定了以下日本蟳的22项形态学指标进行测量:头胸甲长(A)、头胸甲宽(B)、内头胸甲长(C)、额宽(D)、中额齿间距(E)、眼柄长(F)、眼眶宽(G)、额中央至 H 状沟中心长(H)、H 状沟至后缘长(I)、第一侧齿间长(J)、第二侧齿间长(K)、第三侧齿间长(L)、螯足长节长(M)、螯足不动指长(N)、螯足不动指宽(O)、第三颚足座节长度(P)、第三颚足座节宽(Q)、第三颚足长节长度(R)、第三颚足长节宽度(S)、第四步足长节长(T)、第四步足指节长(U)、第四步足指节宽(V),并使用游标卡尺(精确到0.01 mm)对以上形态学指标进行测量。
1.3 数据处理与分析
为了校正日本蟳样品个体大小对形态学参数的影响,我们将所测得的22项形态学参数转化为15项形态比例参数:C/A、D/B、E/B、F/G、H/B、I/B、J/B、K/B、L/B、M/O、N/O、P/Q、R/S、T/V、U/V。应用 SPSS18.0 软件对日本蟳各群体进行聚类分析、主成分分析和判别分析。
1.3.1 单因子方差分析
运用SPSS18.0对双斑蟳群体的15项形态比例参数进行单因子方差分析,并对各项形态比例参数进行多重比较,具有方差齐性的变量采用LSD法,不具备方差齐性的变量采用Tambane’s T2法。7个地理群体按海区归并划分为渤海、黄海和东海组群,计算差异系数,根据MAYR等提出的75%规则,对它们之间的形态学特征差异进行归类。差异系数的计算公式如下:
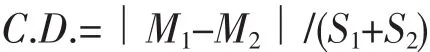
式中,M1、M2和S1、S2分别表示两群体性状的平均值和标准差。
1.3.2 聚类分析
运用SPSS18.0中的分析-分类-系统聚类功能对7个日本蟳群体进行聚类分析,首先计算各个群体15项形态比例参数的均值,然后利用欧氏距离的最短系统聚类法构建聚类树。
1.3.3 主成分分析
运用SPSS18.0对日本蟳群体的15项形态比例参数进行主成分分析,得到各成分的特征值、负荷值和贡献率,特征值>1的成分定为主成分,然后根据各主成分的负荷值绘制主成分散点图。
1.3.4 判别分析
运用SPSS18.0软件中的分析-分类-判别功能建立7个日本蟳群体的判别公式,并计算判别准确率和综合判别率:
判别准确率P(%)=(判别正确的个数/实际个数)×100%。
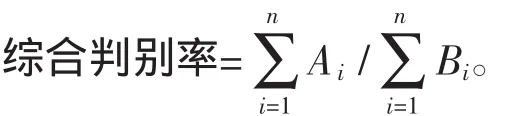
式中,实际个数是指该群体所测量的个体数,判别个数是指被判为该群体的个体数,Ai为第i个群体中判别正确的个体数,Bi为第i个群体的实际个数,n为群体总数。
2 结果
2.1 单因子方差分析
对日本蟳7个地理群体的15项形态比例参数进行单因子方差分析,从表2中可以明显看出,不同海区的日本蟳群体间形态学比例参数均存在显著差异,相同海区内群体的形态学比例参数差异基本不显著。通过对各地理群体间差异系数的计算,初步对群体间差异性进行归类。由表3可知,日本蟳渤海、黄海和东海群体间的各项形态差异均小于1.28,日本蟳不同海区群体间的形态差异属于不同地理群体差异水平。
2.2 聚类分析
采用欧氏距离的最短系统聚类法,对15项形态比例参数的均值进行聚类分析,得到因形态学差异的不同而初步分类的聚类树。从图2中可看出长江口的日本蟳群体单独聚为一支,舟山群体、温州群体和宁波群体首先聚为一小支,随后与威海和东营的日本蟳群体聚集在一起,日照群体随即与它们聚为一支。从聚类图中能看出,当不考虑长江口群体时,各日本蟳群体大致按分布海区的不同而聚集,并且无与地理距离相关的明显聚类,这可能是由于长江口群体位于海流与冲淡水的交汇处,地理环境因素较为复杂,影响了长江口群体的形态学差异性;温州群体与东营群体地理距离最远,但是在聚类分析图中表现不明显,不同海区之间群体存在一定差异性,各海区内群体间差异相对较小。
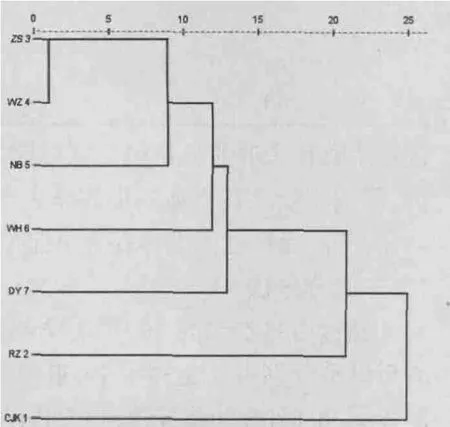
图2 7个日本蟳群体聚类分析图Fig.2 Cluster dendrogram of seven populations of C.japonica
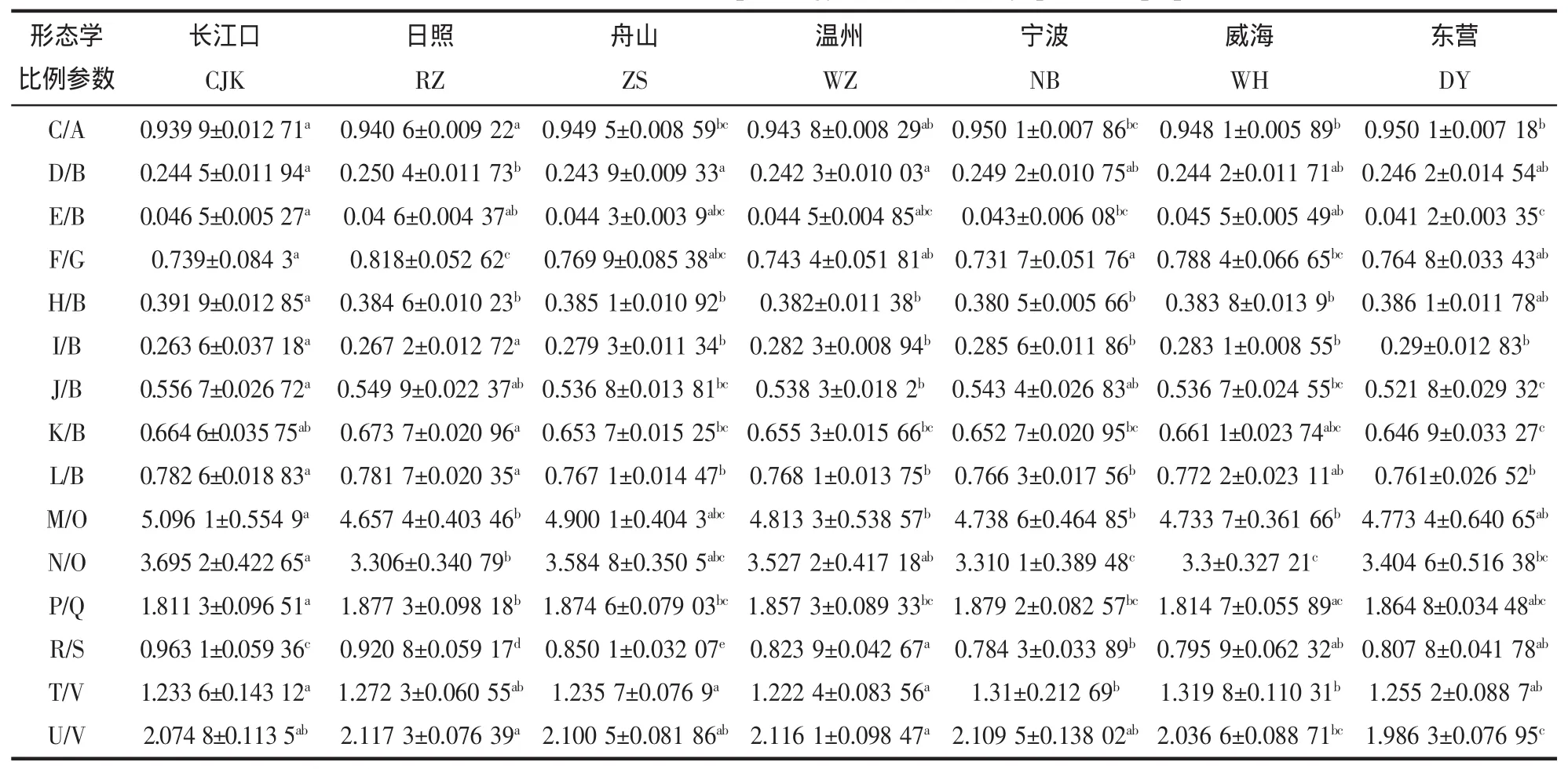
表2 日本蟳群体间15个形态学比例参数的差异Tab.2 Differences on fifteen morphology ratios of C.japonica populations
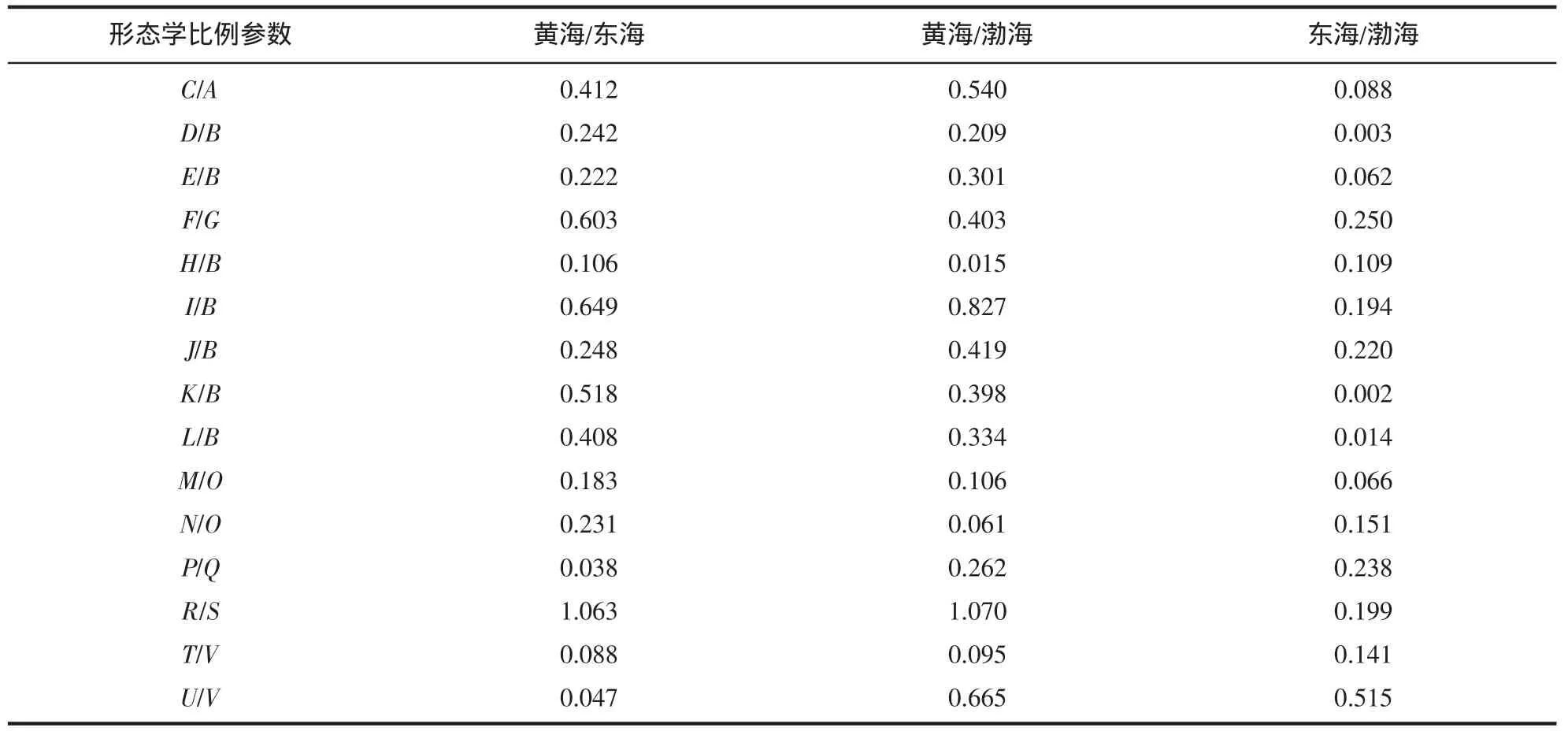
表3 日本蟳不同海区群体间形态学比例参数的差异系数(C.D.值)Tab.3 Coefficients difference of morpholoy ratios between different sea areas of C.japonica populations
2.3 主成分分析
通过对日本蟳的15项形态学比例参数进行主成分分析,我们得到了降维后的4个主成分,各主成分的贡献率分别为47.639%、19.004%、12.270%、10.178%,其累计贡献率为89.090%,表明4个主成分能解释群体间89.090%的差异性,很大程度上保留了原始数据所带有的信息。其中对第一主成分贡献较大的形态学比例参数为:D/B(额宽/头胸甲宽)、E/B(中额齿间距/头胸甲宽)、F/G(眼柄长/眼眶宽)、J/B(第一侧齿间长/头胸甲宽)、K/B(第二侧齿间长/头胸甲宽)、L/B(第三侧齿间长/头胸甲宽)、T/V(第四步足长节长/第四步足指节宽)、U/V(第四步足指节长/第四步足指节宽);对第二主成分影响最大的为P/Q(第三颚足座节长度/第三颚足座节宽)。
利用第一、第二主成分的负荷值绘制主成分分布图,从图3中可观察采用主成分分析对不同日本蟳群体的分类结果。如图3所示,日照群体与其它6个群体相距最远,但也出现部分重叠;长江口、舟山、温州、东营群体大部分重叠在一起,宁波群体和威海群体几乎完全重叠。主成分分析的结果与聚类分析结果并不完全一致,主成分分布图没有表现出明显的形态差异与地理分化间的规律。
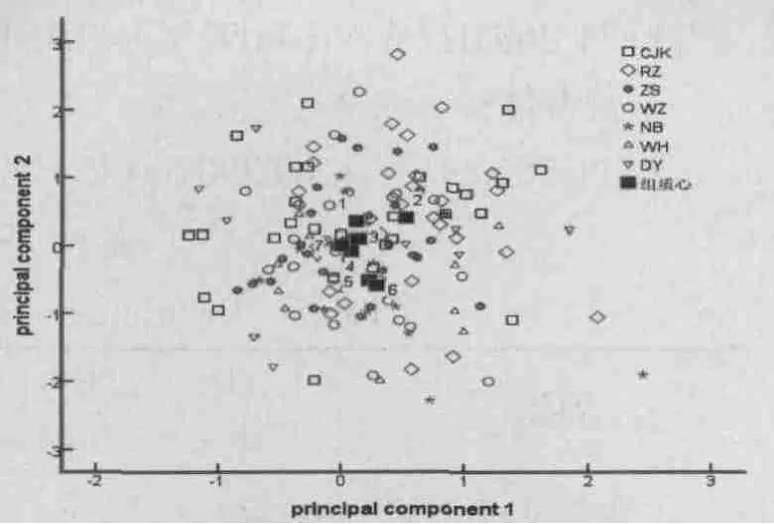
图3 日本蟳群体的主成分分布Fig.3 Scatter diagram of the principal component of C.japonica populations
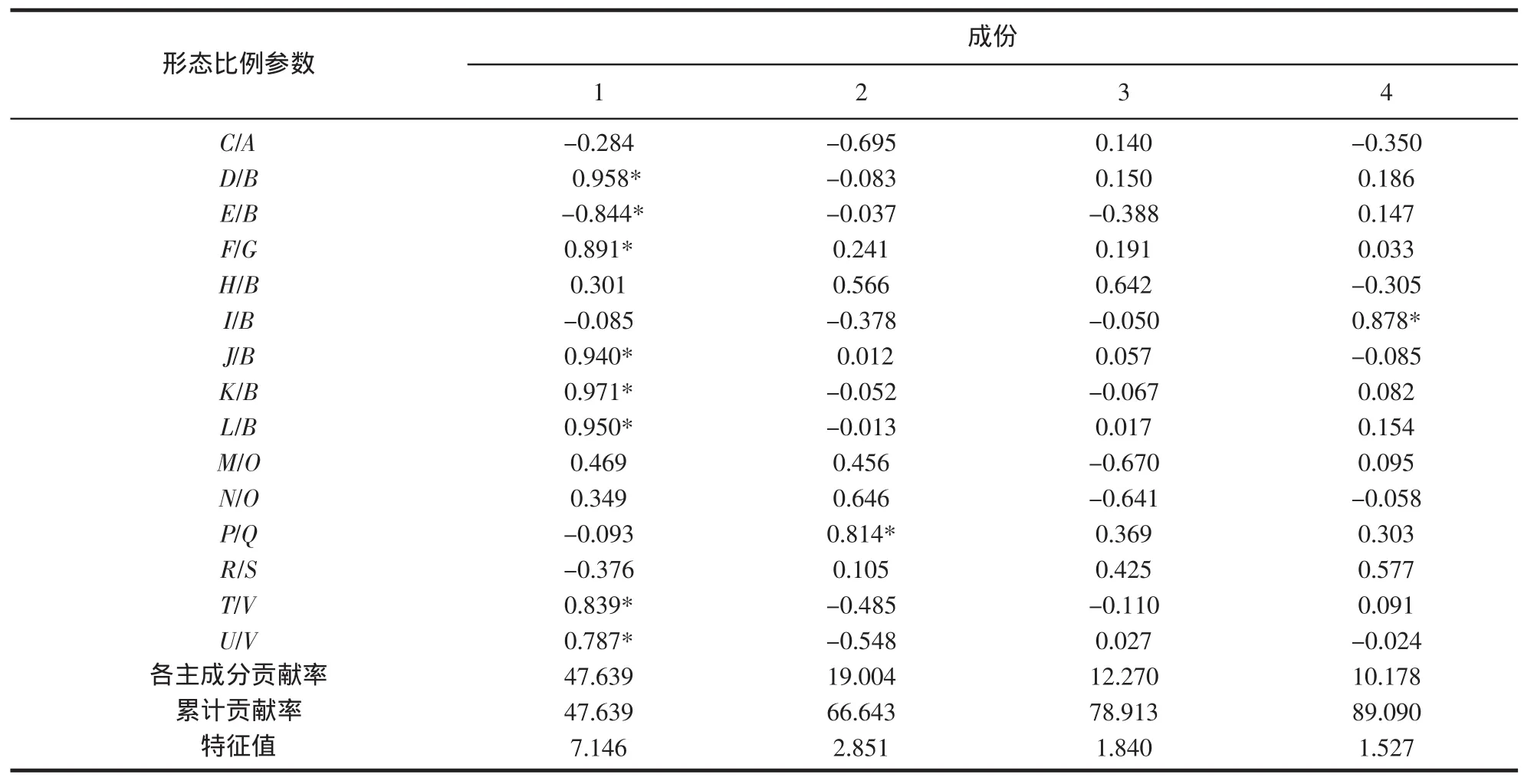
表4 7个日本蟳群体的主成分负荷值和特征值Tab.4 Eigen values and loadings of seven populations of C.japonica
2.4 判别分析
采用逐步判别法对15项形态比例参数进行判别分析,筛选出贡献率大的5个参数建立判别函数。长江口群体的判别率最高为73.3%,舟山和威海的判别率最低,均为40%,7个群体的综合判别率为56.5%。日本蟳各群体的判别结果见表5,判别公式如下:
长江口群体:
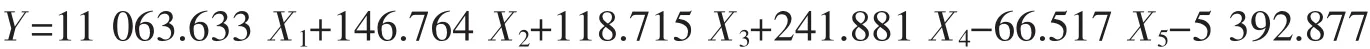
日照群体:
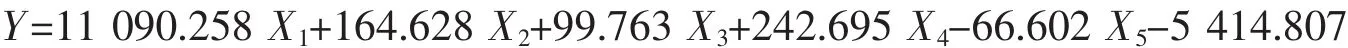
舟山群体:
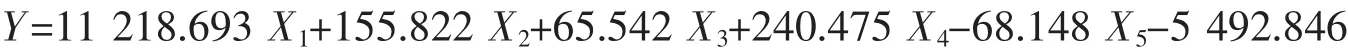
温州群体:
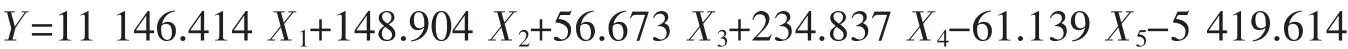
宁波群体:
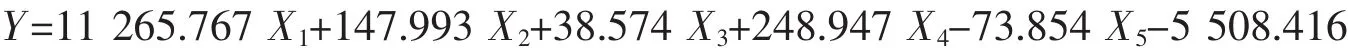
威海群体:
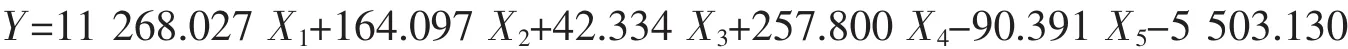
东营群体:
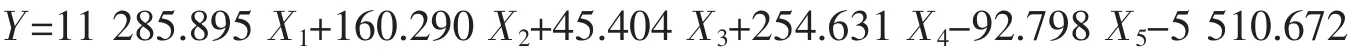
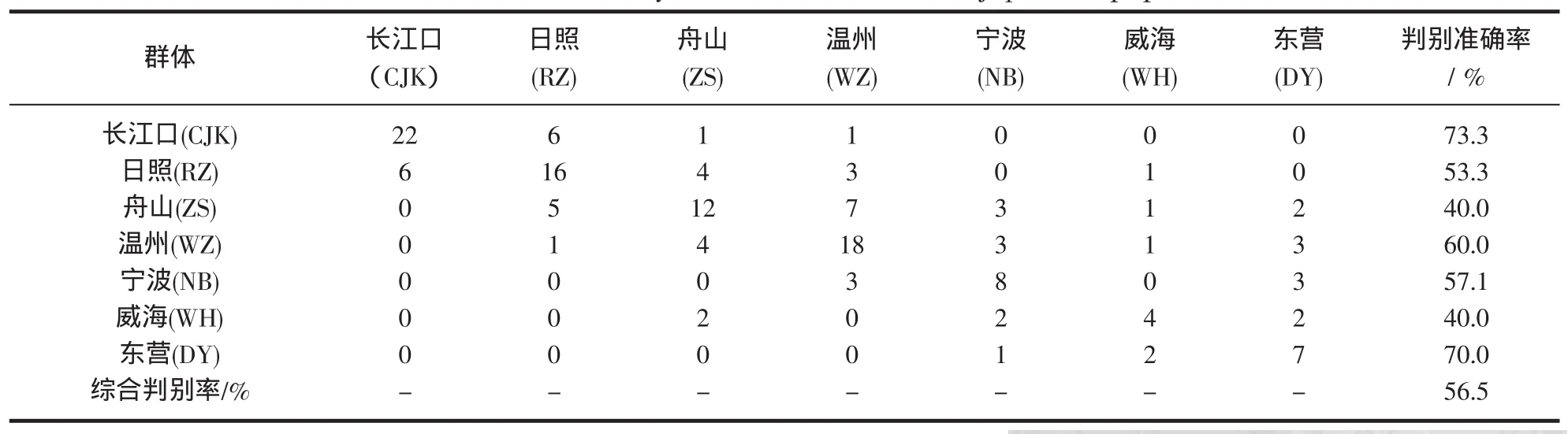
表5 日本蟳7个群体的判别分析结果Tab.5 Discriminate analysis results of seven C.japonica populations
式中X1代表内头胸甲长/头胸甲长、X2代表眼柄长/眼眶宽、X3代表第三颚足长节长度/第三颚足长节宽度、X4代表第四步足长节长/第四步足指节宽、X5代表第四步足指节长/第四步足指节宽,根据建立的判别公式可以对日本蟳进行快速简便的分类。
通过判别分析可得两个典型判别函数,其贡献率分别为76.4%和14.2%,累计贡献率为90.7%。基于前两个典型判别函数值绘制散点图(图4),从图4中明显可以看出威海和东营群体几乎完全重合在一起,说明两群体间差异性最小;长江口群体与日照群体大部分重叠在一起,表明两群体的相似度很高;舟山、温州和宁波群体间也有很大部分的交叠,彼此间差异也较小。判别分析的结果显示7个日本蟳大致分不同的海区聚在一起,但各海区群体间也存在部分重叠,并没有完全分离。如属于黄海的威海群体和渤海的东营群体聚为一类,属于黄海的日照群体与东海边缘的长江口群体聚为一类,属于东海的宁波、舟山和温州为一类。
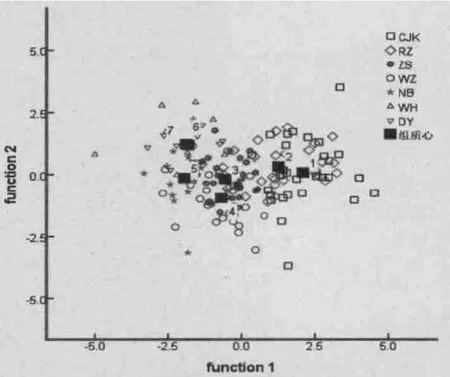
图4 典型判别函数散点图Fig.4 Scatter plots of canonical discriminant functions
3 讨论
形态学分析方法是传统的分类、鉴别方法,由于形态学指标简单、易获得,这种生物学测定方法被广泛应用于海洋生物种群鉴定研究,如孙成波等[14]对凡纳滨对虾Litopenaeus vannamei4个群体的形态差异研究、陈蓉等[15]对5个海区不同群体的毛蚶Scapharca subcrenata形态差异研究、董志国等[16]对三疣梭子蟹Portunus trituberculatus5个群体的形态差异分析等。
本文采用单因子方差分析、聚类分析、主成分分析和判别分析对日本蟳日照、长江口、舟山、温州、宁波、东营和威海群体进行形态学差异分析。这四种方法分析结果相似,但各自从不同的角度分析了群体间的形态学差异。本文的单因子方差分析结果表明日本蟳东海群体、渤海群体和黄海群体间存在一定程度的形态学差异,但属于不同地理群体差异水平;聚类分析的结果表明,各群体大致按分布海区的不同而聚集,长江口群体由于其地理位置的特殊性而单独聚为一支,不同海区之间群体存在一定差异性,各海区内群体间差异相对较小;主成分分析结果显示各日本蟳群体的主成分没有按照海区分布,主成分分析的结果与聚类分析结果并不完全一致,主成分分布图没有表现出明显的形态差异与地理分化间的规律;判别分析的结果表明日本蟳7个群体的综合判别率为56.5%,典型判别函数的累计贡献率为90.7%,说明判别效果比较理想。判别分析的结果与聚类分析的结果基本一致,进一步论证了日本蟳不同海区群体之间存在一定差异性。主成分分析的结果与聚类分析和判别分析有出入,导致这种结果的可能原因是威海、东营和宁波群体所采的样品个数过少,影响了主成分的准确性。
综合上述四种多元分析的结果,我们认为日本蟳渤海群体、黄海群体和东海群体间存在一定的形态学差异,但并没有形成地理隔离;受地理环境因素的影响,各海区群体间存在一定的交叉,如威海群体与长江口群体;各海区内群体差异性很小,无明显的地理分化现象。本文研究结果与丁金强等[12]对日本蟳4个群体的形态学分析结果基本一致。
为了对上述现象进行合理的解释,我们从日本蟳的生物学特性以及地理环境因素进行分析。日本蟳作为一种沿岸定居性蟹类,其成体的扩散能力可能有限,且蟹类均有抱卵行为,这就导致日本蟳受精卵不具备自由扩散的能力,普遍认为蟹类群体间的交流是通过处于浮游期的幼体所完成的。LIU等[17]和HEWITT等[18]的研究也表明,具有高度扩散能力的海洋生物不同地理群体间遗传差异很小。位于黄海的威海群体与渤海的东营群体之间的交流受到自西向东的鲁北沿岸流的影响,导致两群体间的形态差异性很小;位于黄海的日照群体受到黄海沿岸流的冲击作用,与位于东海的长江口群体发生了交流,因此两群体间的形态差异性很小;东海的温州、舟山和宁波群体则是因为东海沿岸流的影响而导致群体间差异不明显。自北向南的黄海沿岸流在经过长江口附近时,受到自西向东扩展的长江冲淡水影响,影响了黄海群体与东海群体之间的交流,造成了两群体间的形态学差异性。长江冲淡水的阻隔作用也在其它海洋生物的研究中发现,DONG等[19]对花笠螺Cellana toreuma的研究显示,由于受到长江冲淡水的影响,导致了花笠螺的黄海群体与东海和南海群体间的遗传结构差异性。鲁北沿岸流在绕过成山头顺鲁南沿岸和苏北沿岸南下时,遇到了自南向北的黄海暖流,这在一定程度上影响了渤海群体与黄海群体间的交流,造成群体间的形态学差异性,董志国等[21]对三疣梭子蟹5个群体的研究也出现了相似的结果。
形态学差异除了受到基因交流的影响,还受到环境因素的影响。渤海、黄海和东海的气候、水温、盐度等生活环境差异也可能导致日本蟳群体形态的差异,这些环境因素与形态学差异的相关性还有待进一步的研究。
[1]俞存根,宋海棠,姚光展.东海日本蟳的数量分布和生物学特性[J].上海水产大学学报,2005,14(1):40-45.
[2]俞存根,宋海棠,姚光展.东海大陆架海域蟹类资源的评估[J].水产学报,2001,28(1):41-46.
[3]叶孙忠,张壮丽,叶泉土.福建南部沿海日本蟳的生物学特性[J].福建水产,2002(4):18-21.
[4]吴常文,王志铮,王伟洪,等.舟山近海日本蟳生物学、资源分布以及开发利用[J].浙江水产学院学报,1998,17(1):13-18.
[5]黄培民.闽东北外海蟹类种类组成[J].福建水产,2005(2):24-27.
[6]王春琳,陈建青,叶晓圆,等.日本蟳的营养成分组成分析[J].营养学报,2005,27(1):81-83.
[7]王春琳,薛良义,刘凤燕,等.日本蟳繁殖生物学的初步研究I雌性繁殖习性[J].浙江水产学报,1996,15(4):261-266.
[8]刘洪军,戴玉蓉,张富君,等.日本蟳人工育苗及养殖技术研究[J].海洋科学,2000,24(8):23-27.
[9]王春琳,薛良义,刘凤燕,等.日本蟳繁殖生物学的初步研究[J].齐鲁渔业,1998,15(3):18-20.
[10]王春琳,蒋霞敏,陈惠群,等.日本蟳繁殖生物学的初步研究II雄性繁殖习性及胚胎发育[J].东海海洋,2000,18(1):43-49.
[11]闫 愚,孙颖民,段 钰.日本蟳胚胎发育的初步观察[J].齐鲁渔业,1998,15(1):18-20.
[12]丁金强,刘 萍,李 建,等.中国沿海日本蟳4个地理群体的形态差异比较分析[J].中国水产科学,2012,19(4):604-610.
[13]高天翔,张秀梅,柳广东,等.10个日本绒螯蟹群体与中华绒螯蟹形态的主成分分析[J].大连水产学院学报,2003,18(1):273-277.
[14]孙成波,陈国良,董汉荣,等.美国4个凡纳滨对虾种群形态差异与判别分析[J].海洋与湖沼,2009,40(1):27-32.
[15]陈 蓉,刘建勇,唐连俊,等.中国5个海区不同群体毛蚶形态差异分析[J].海洋科学,2009,33(7):64-69.
[16]董志国,李晓英,阎斌伦,等.中国海区五种群三疣梭子蟹的形态差异分析[J].海洋通报,2010,29(4):421-426.
[17]LIU J X,GAO T X,WU S F,et al.Pleistocene isolation in the Northwestern Pacific marginal seas and limited dispersal in a marine fish,Chelon haematocheilus(Temminck and Schlegel,1845)[J].Mol Ecol,2007,16:275-288.
[18]HEWITT G M.The genetic legacy of the Quaternary ice ages[J].Nature,2000,405:907-913.
[19]DONG Y W,WANG H S,HAN G D,et al.The Impact of Yangtze River Discharge,Ocean Currents and Historical Events on the Biogeographic Pattern of Cellana toreuma along the China Coast[J].PLoS ONE,2012,7(4):e36178.
