牛Fas相关死亡功能域蛋白(FADD)研究进展
2013-09-22张文刚杨润军
张文刚,杨润军
(吉林大学动物科学学院,吉林 长春 130062)
Fas相关死亡功能域蛋白最早是在酵母双杂交系统中发现的一种具有与Fas死亡域和TNFR-1死亡域高度同源域的蛋白,并很快被证明是Fas受体和TNFR-1受体诱导的细胞凋亡信号衔接蛋白[1]。近年来随着对FADD进一步研究发现,FADD在细胞内能与很多信号转导途径相关蛋白作用,参与细胞增殖、细胞周期进程、胚胎发育、炎症反应、肿瘤发生等生物学活动。本文从FADD基因结构、蛋白修饰、参与信号途径及其不同的生物学作用进行系统阐明和论述。
1 牛FADD基因及蛋白的结构
牛的FADD基因位于29号染色体,基因全长2406kb,编码区包含两个外显子,即长度为288bp编码死亡效应结构域(Death Effector Domain,DED)的外显子Ⅰ和长度为342bp编码死亡结构域(Death Domain,DD)的外显子Ⅱ。研究发现其上共有7个单核苷酸突变位点。
不同物种间FADD基因结构、基因序列之间存在一定差异(表1)。FADD蛋白一级结构既存在差异,又具有许多保守氨基酸位点。同源性分析结果显示,牛与野猪FADD序列同源性高达87%、与人类为82%。因此,显示出这些氨基酸位点对FADD发挥功能起重要作用。通过核磁共振等生物化学方法分析FADD结构发现,FADD的DD和DED分别由6个α螺旋组成,并成尾连尾的正交垂直结构[3]。此外,DED 包含前体caspase-8和前体caspase-10结合位点。
2 FADD蛋白质修饰
2.1 FADD的泛素化修饰
近年来研究发现,FADD存在泛素化修饰,即蛋白酶对FADD进行特异性剪切,使FADD在其参与的细胞凋亡或增殖途径中发挥相应作用。目前,已发现有两种蛋白酶对FADD具有泛素化修饰的能力,即颗粒酶 M(GzmM)和 Makorin环指蛋白1(MKRN1)。
GzmM是诱导caspase途径和细胞色素c介导的细胞凋亡途径中重要的蛋白酶,研究结果显示,GzmM在自然杀伤细胞中特异性的在196号位蛋氨酸以后剪切形成tFADD,使得自身寡聚化能力提高,进而促进前体caspase-8聚集、加工,及一系列的级联反应[4]。
实验证明,Makorin环指蛋白1(Makorin RingFinger Protein 1,MKRN1)对细胞内FADD蛋白进行泛素化修饰,使FADD失去信号衔接作用。将MKRN1基因敲除可以使FADD在细胞中稳定存在,并迅速合成大量死亡诱导信号复合体(death-inducing signaling complex,DSIC),从而激活了胞外途径的细胞凋亡。
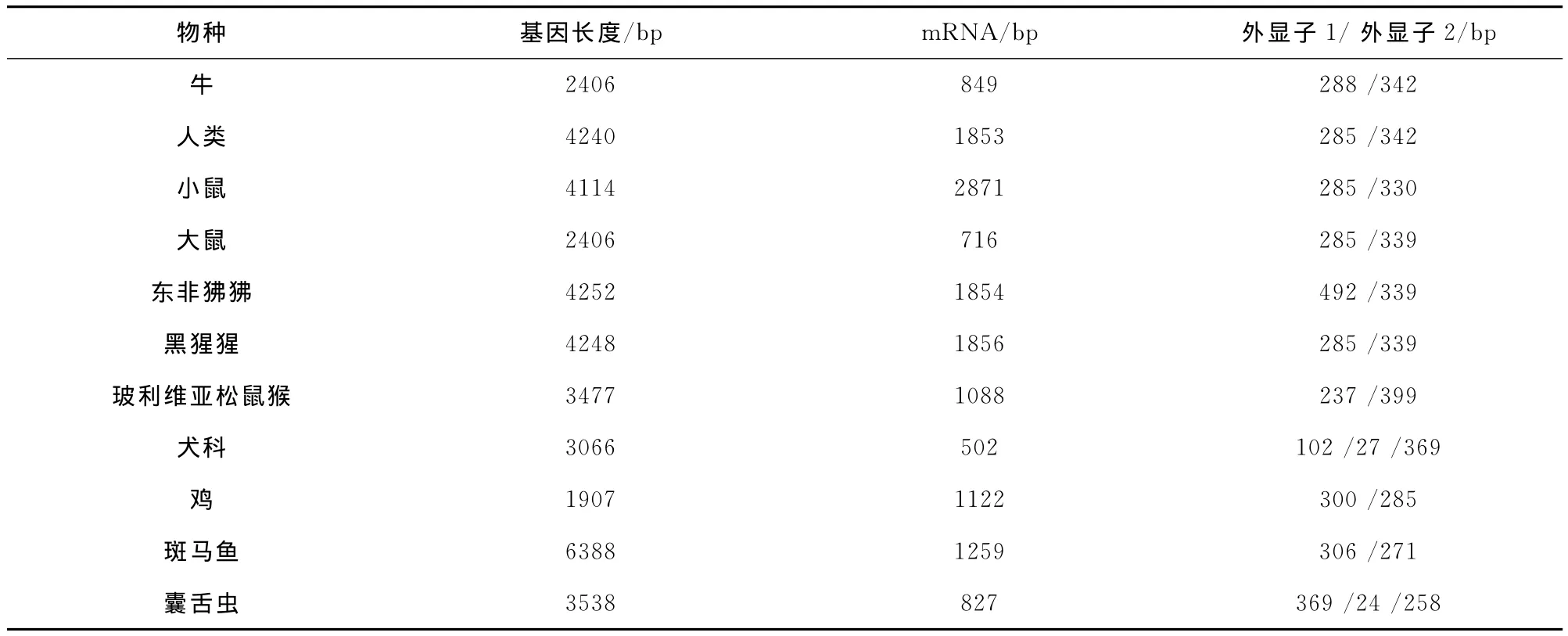
表1 不同物种FADD基因、mRNA和外显子长度
2.2 FADD的磷酸化修饰
磷酸化作为FADD的主要修饰方式,是保证FADD发挥多种生物学功能的重要前提。研究发现,磷酸化FADD在细胞增殖和细胞周期进行等过程中起着重要作用[5,6]。一般哺乳动物FADD磷酸化氨基酸位点为Ser-194,除此之外还存在Ser-187和Ser-203位点(见图1)。实验证明,酪蛋白激酶1α(casein kinase 1α,CK1α)能使FADD Ser-194磷酸化,并能形成FADD-CK1α复合体,在细胞有丝分裂中期将磷酸化FADD定位于纺锤丝两极,参与有丝分裂的进行。当小鼠脾细胞缺乏CK1α,磷酸化FADD数量明显减低,并且细胞有丝分裂受到抑制作用[7]。
Polo样激酶1(polo-like kinase 1,PLK1)是细胞在有丝分裂G2期和M期形成中心体、纺锤体两极的关键蛋白。实验证明,利用紫杉醇处理过的细胞,只有在FADD Ser-203位点被 Aur-A 磷酸化前提下,PLK1才能将 FADD Ser-187磷酸化[8],磷酸化FADD蛋白又负反馈作用于PLK1,并且具有抑制肿瘤增殖、提供细胞死亡潜在可能等作用[9]。
(3)钾化:为矿区最早的蚀变,以形成大面积的面状钾长石为特征,蚀变后岩石整体呈肉红色,主要矿物为钾长石,少量石英和绢云母及条带状黄铁矿,钾长石呈细脉或粒(斑)状分布于矿体和蚀变围岩中,因受挤压而破碎,被绢云母化交代、叠加而呈港湾状,可见硅质细脉及黄铁矿细脉穿插其中。
随着更多FADD磷酸化激酶被发现和磷酸化FADD作用机制被阐明,FADD生物学功能被深刻的了解。有人指出在一些粘附细胞株中,FADD聚集于细胞核,FADD Ser-194磷酸化后才能从胞核转移至胞浆。胞质内FADD的定位,依赖于磷酸化FADD与核浆穿梭蛋白exportin-5的相互作用。所以说磷酸化FADD是调控细胞增殖与凋亡平衡的关键蛋白[6],磷酸化和非磷酸化FADD之间的平衡,是细胞保持内稳定的重要前提。磷酸化FADD在细胞中作用的分子机制,为肿瘤疾病的治疗提供了新的思路。但磷酸化FADD亚细胞定位[6]以及磷酸化和非磷酸化FADD平衡调控机制有待进一步研究。
3 FADD生物学功能
3.1 FADD参与细胞凋亡
程序性死亡(PCD)即细胞凋亡,对于多细胞生物的正常发育和内环境稳定至关重要,在组织分化、器官发育、机体稳态的维持中具有重要意义。凋亡是指细胞的形态学发生改变,伴随细胞收缩、细胞膜破裂、染色体凝缩等形态学改变。与之对应是坏死,通常指细胞在受毒素刺激后细胞逐渐破裂溶解的过程。目前已知的三种主要凋亡途径包括:细胞膜上死亡受体(DRs)途径,内质网应激途径、线粒体凋亡途径。
肿瘤坏死因子受体家族成员在细胞增殖、分化和凋亡过程中各自起着不同作用。目前已知19个死亡受体中主要有:肿瘤坏死因子(TNFR-1)、CD40L、CD27L、Fas等。FADD主要参与Fas/Fa sL途径和TNFR-1途径细胞凋亡,是信号传递中的衔接蛋白。但有研究发现FADD也是TRAIL途径细胞凋亡的衔接蛋白[10]。

图1 人类、小鼠、大鼠、黑猩猩、松鼠猴、绿狒狒、鸡、牛、斑马鱼FADD核酸保守序列分析
3.1.1 FADD参与Fas/FasL途径细胞凋亡 Fas基因位于牛26号染色体长臂(26q13),基因编码产物为45KD I型跨膜蛋白,主要分布于胸腺细胞、外周活化T、B淋巴细胞、NK细胞和部分组织细胞。Fas配体(Fas Ligand,FasL)是40KD II型跨膜蛋白,主要表达并存在于T、B淋巴细胞表面,亦可被释放至胞外形成可溶性功能活性分子[11,12]。Fas/FasL途径诱导免疫细胞、生殖组织细胞等多数细胞凋亡,在维持组织正常发育、控制免疫反应、调节机体生理平衡中起重要作用。
FADD参与Fas/FasL途径细胞凋亡过程是:表面存在Fas蛋白受体的细胞与FasL结合,形成三聚体并在细胞内开始募集FADD,即Fas细胞内的DD与FADD的DD高度同源并反向平行连接。一旦FADD被激活,FADD的DED会与细胞内游离前体caspase-8或少数前体caspase-10的DED结合,至此以上蛋白(Fas的 DD、FADD、procaspase-8,-10)形成的三聚体被称为DISC(见图2)。
前体caspase-8和前体caspase-10经过修饰形成caspase-8和caspase-10,会在细胞内发生一系列的caspase级联反应[13,14],进而细胞发生凋亡。但胞内游离的c-FLIP(cellular FLICE-inhibitor protein)的DED也与FADD的DED存在高度同源性,因此会和前体caspase-8竞争结合,抑制细胞凋亡[15,16](见图3)。
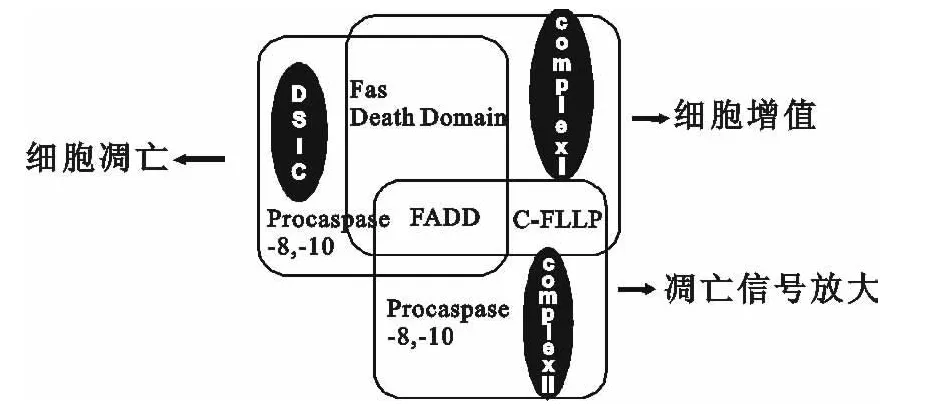
图2 Fas/FasL诱导细胞凋亡中,三种复合体组成及功能
研究发现,胞质内脱离Fas后的FADD与前体caspase-8、c-FLIP共同形成的复合体具有放大通路凋亡信号作用[13],作用机制有待进一步的研究。
3.1.2 FADD参与 TNF-R1途径细胞凋亡 牛TNF基因位于22号染色体长臂(22q23),编码蛋白为234个氨基酸单位II型跨膜蛋白。表达于包括淋巴细胞、肥大细胞、内皮细胞、心肌细胞等多数细胞。当细胞受到脂多糖、细菌毒素或白介素的刺激时,会大量表达。皮肤受到损伤出现炎症时,肥大细胞大量分泌前体TNF。TNF-R1途径细胞凋亡在先天免疫、炎症发生、抗肿瘤细胞等中起重要作用。
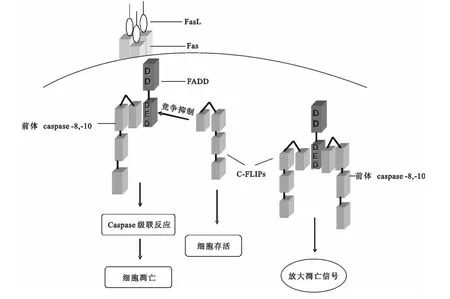
图3 FADD参与Fas/Fasl诱导细胞凋亡
TNF-R1诱导细胞凋亡分为两个步骤:首先,细胞膜上受体蛋白TNF-R1在外源因子TNFα的刺激,细胞膜内侧会募集形成TNF-R1-TRADDTRAF2-RIP复合体,激活 NF-κB细胞存活信号。在细胞执行完生命活动后,进行第二个步奏,上述复合体从细胞膜内侧脱落,在胞质内形成TRADDTRAF2-RIP-FADD-Procaspase-8,-10复合体[13,17]。前体caspase-8和前体caspase-10经过修饰形成caspase-8和caspase-10之后会在细胞内发生一系列的caspase级联反应,引起细胞发生凋亡。
3.1.3 FADD参与内质网应激途径细胞凋亡 内质网应激(endoplasmic reticulum stress,ERS)是细胞抵抗不良外界刺激的重要机制,也是应激损伤细胞的重要机制[18]。内质网应激能激活细胞保护机制,目前发现同型半胱氨酸、氧化应激、钙离子代谢紊乱等都能引起内质网应激级联反应[19]。但应激过度时,可导致细胞凋亡。实验证明,FADD参与内质网应激途径细胞凋亡。在衣霉素作用下FADD被募集至内质网与RTN3相互作用,激活FADD依赖型caspase-8、-9级联反应,引起细胞凋亡。与此同时,Bid被切割引起细胞色素C从线粒体中释放,也能引起细胞凋亡[20]。由此可以看出FADD参与内质网应激途径的细胞凋亡,与线粒体途径细胞凋亡也存在联系。
3.2 FADD与T淋巴细胞增殖
除了介导细胞凋亡的生理功能外,FADD在T淋巴细胞增殖中起着重要作用[21]。实验证明,FADD隐性突变小鼠T淋巴细胞由于凋亡而活性受到抑制[22]。又有实验证明5周大FADD基因敲除小鼠与野生型小鼠相比,胸腺细胞CD4+CD8+量显著减少,并且T淋巴细胞有丝分裂过程受到抑制[13]。
FADD S-191被天冬氨酸取代后得到的转基因小鼠FADD(S191D),其FADD蛋白处于一种模拟的永久磷酸化状态。这样的小鼠其发育过程不正常,表现出了和免疫系统FADD缺失的小鼠相似的表型,小鼠能存活但表现出贫血和脾大,只有很少的CD4+CD8+胸腺细胞,而其免疫系统发育方面的问题也暗示了增殖的缺陷。
有研究证明,在T细胞中条件性敲除caspase-8显示出与FADD相似的T细胞增殖缺陷。T细胞条件性敲除c-FLIP也能导致相似的T细胞增殖缺陷。因此,FADD、caspase-8和c-FLIP不仅协同参与细胞凋亡,同时也参与促进T淋巴细胞的增殖过程。所以,有待于进一步的实验证明依赖FADD途径T细胞增殖的分子机制,才能全面的了解FADD的生理功能。
3.3 FADD与组织融合及胚胎发育
在胚胎发育过程中,组织融合对个体生物器官的形成,如:神经管、上颚、眼裂等起着重要作用。组织融合相关调控基因如果发生缺失或突变,就会引起先天畸形等症状。
研究表明,在FADD缺失的发育胚胎中,细胞表现为增殖能力增强、RIP1/RIP3调控的程序性坏死途径激活、发育过程中出现眼裂无法闭合的现象。进一步研究发现,FADD基因的表达受细胞内Pax2因子和Vax2因子的调控。通过对大鼠FADD基因编码区上游序列分析,发现了两个Pax2因子结合位点(位于-381、-158)和一个Vax2蛋白因子的结合位点(位于-246)。野生型发育胚胎在发育过程中,Pax2因子与Vax2因子会结合到FADD基因调控区的相关位点,使FADD基因进行正常转录与表达,保证了胚胎组织眼部发育融合的正常发生。如果胚胎细胞中Pax2或Vax2缺失,会导致FADD表达量不足,引起RIP1/RIP3介导的程序性坏死途径激活,眼裂无法正常闭合。因此以上事实说明FADD是Pax2与Vax2的下游目的基因,是调控眼部组织融合的分子开关[23]。又已知FADD-caspase-cFLIP复合体在胚胎发育过程中,阻止RIP3介导的程序性细胞凋亡[24],故推断此为FADD阻止眼裂非正常融合的分子机制。
上述事实说明FADD参与胚胎发育过程中的组织融合,又有实验证实FADD基因缺失的小鼠胚胎在形成后11.5d 就会死亡[25],且该过程与caspase-8,-10无关,由此推测在胚胎发育过程中FADD不仅参与凋亡信号转导,同样参与非凋亡信号转导。
3.4 FADD与炎症发生
炎症是血管组织对有害刺激如:病原体、受损细胞、刺激物等的一种防御性反应,并通过除去有害刺激和启动愈合程序,使生物体受到保护的一种机制。通常情况下,炎症是有益的,是人体的自动的防御反应,但是有的时候,炎症也是有害的,例如对人体自身组织的攻击、发生在透明组织的炎症等等。
炎症的发生主要是通过三个不同信号通路:JAK-STAT、MAPK、NF-κB[26]。其中,FADD 对NF-κB途径的炎症发生具有抑制和促进两种相反作用。风湿性关节炎中,NF-κB通路的形成需要细胞膜表面的受体激活,这些受体包括:Toll样受体4(Toll-like receptor 4,TLR4)、白介素1受体(interleukin 1receptor,IL-1R)。在热休克蛋白60(HSP60)、纤连蛋白 A(Fibonectin A)、透明质酸(Hyaluronic acid)等作用下,激活 TLR4受体;在白介素1β作用下,激活IL-1R受体。随着上述受体的激活,细胞内MyD88蛋白与IRAK蛋白结合,标志着NF-κB途径炎症发生,分泌白介素1、白介素6、白介素8、TNFα等。当细胞中存在FADD蛋白时会与IRAK竞争结合MyD88,从而抑制炎症的发生。
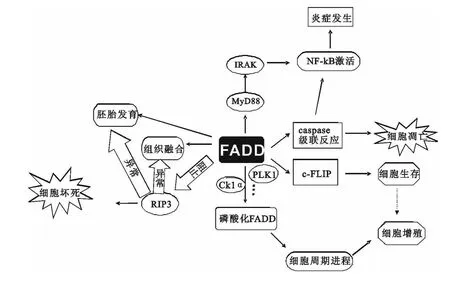
图4 FADD已知生物学功能
但FADD蛋白在TNF-R1和Fas介导的NF-κB途径中同样起着关键作用[13,27]。实验证明,TNF-α是重要的促炎因子,在缺失FADD蛋白细胞中TNFR1和Fas介导的NF-κB途径受到彻底抑制。并且Fas-FADD复合体过表达会引起细胞调亡和炎症的发生。
FADD除了参与上述两种炎症发生途径外,在RIP3介导的细胞坏死途径而引发的慢性肠炎中起抑制作用[28,29]。以上事实证明FADD蛋白在炎症发生过程中存在于多种途径,并扮演不同的角色。现已明确部分细胞只存在某一种通路,不同通路间存在相互作用,但机体如何协调FADD在各通路中的作用机制目前尚不明确。所以,亟待实验进一步证明。
4 展望
在科技水平日益发达的今天,各种新技术手段对FADD蛋白的深入研究起到了积极作用。通过10年左右的研究,不断地发现FADD如何作为一个多功能蛋白在基因层面、蛋白质层面对机体生理功能的影响。图4描述的是以FADD为中心的相关生物学功能。从图中可以看出FADD有关的生物学功能并非孤立存在,而是具有内在联系。不同种类细胞、不同发育阶段细胞、不同外界刺激等这些变量是FADD在细胞中具有不同生物学功能、介导不同信号通路的根本原因。尽管国内外研究者对FADD的了解日渐加深,但关于牛FADD基因的相关报道不多见。能否将候选基因FADD作为肉牛肉质和胴体性状遗传标记还是一个亟待解决的问题。
焦裴等曾提出FADD上游蛋白FAS基因型可能与肉牛肉质和胴体相关,因此在今后的研究中根据FADD基因的多态性来判断其基因型,找出各种基因型与肉牛肉质和胴体之间的相关性从而将分子生物学技术应用到育种和生产中去。
[1] Zhang Jianke and Astar Winoto.A Mouse Fas-Associated Protein with Homology to the Human Mort1/FADD Ptotein Is Essential for Fas-Induced Apoptosis[J].Molechlat and Cellular Ciology,1996JUNE,16(6):2756-2763.
[2] Huang B H,Matthias E,Edward TO,et al.NMR structure and mutagenesis of the Fas(APO-1/CD95)death domain[J].Nature,1996,384(19/26):638-641.
[3] Paul E C,Cristinel S,Wei YF,et al.The Structure of FADD and Its Mode of Interaction with Procaspase-8[J].Molecular Cell,2006,22(5):599-610.
[4] Wang S,Xia P,Shi L,et al.FADD cleavage by NK cell granzyme M enhances its self-association to facilitate procaspase-8recruitment for auto-processing leading to caspase cascade[J].CELL DEATH AND DIFFERENTATION,2012,19(4):605-615.
[5] Hua Z C,Sue J S,Chulho K,et al.A Function of Fas-Associated Death Domain Protein in Cell Cycle Progression Localized to a Single Amino Acid at Its C-Terminal Region[J].Immunity,2003,18(4):513-521.
[6] Zhang J,Zhang D P,Hua Z C.FADD and its Phosphorylation[J].IUBMB Life,2004JULY,56(7):395-401.
[7] Elizabeth C A,Christine F,Benjamin B,et al.Molecular Cell Phosphorylation of FADD at Serine 194by CKIαRegulates Its Nonapoptotic Activities[J].2005,19(3):321-332.
[8] Jang Moon-Sun,Su-Jin Lee,Nam Sook Kang,et al.Cooperative Phosphorylation of FADD by Aur-A and Plk1in Response to Taxol Triggers Both Apoptotic and Necrotic Cell Death[J].CANCER RESEARCH,2011,71(23):7207-7215.
[9] Jang M S,Lee S J,Kim C J,et al.Phosphorylation by polo-like kinase 1induces the tumor-suppressing activity of FADD[J].Oncogene,2011,30(4):471-481.
[10] Pascal S,Margot T,Kim B,et al.TRAIL Receptors 1(DR4)and 2(DR5)Signal FADD-Dependent Apoptosis and Activate NF-kB[J].Immunity,1997,7:831-836.
[11] 袁 红.Fas/FasL 与凋亡研究进展[J].Foreign Medical Sciences,2000,20(1):26-28.
[12] 朱锡华.Fas系统研究进展[J].中华微生物和免疫学杂志,1996,16(2):77-82.
[13] Marta M,Arul M C,Frank C K,et al.FLICE,A Novel FADD-Homologous ICE/CED-3-like Protease,Is Recruited to the CD95 (Fas/APO-1)Death-Inducing Signaling Complex[J].Cell,1996,(85):817-827.
[14] Medema J P,Scaffidi C,Frank C K,et al.FLICE is activated by association with the CD95death-inducing signaling complex(DISC)[J].The EMBO Journal,1997,16(10):2794-2804.
[15] Lea Tourneur and Gilles Chiocchia.FADD:a regulator of life and death.Cell.2010,31(7):260-269.
[16] Safa A R.c-FLIP,A MASTER ANTI-APOPTOTIC REGULATOR[J].Experimental oncology,2012,34(3):176-184.
[17] Olivier Micheau and Jürg Tschopp.Induction of TNF Receptor I-Mediated Apoptosis via Two Sequential Signaling Complexes[J].Cell,2003,(114):181-190.
[18] 项 荣,谭志平,林云婷,等.RTN3与细胞凋亡[J].生命科学研究,2010,14(1):80-84.
[19] 李载权,周爱儒,唐朝枢.内质网应激反应分子机理研究进展[J].中国生物化学与分子生物学报,2004,20(3):283-288.
[20] Rong X,Liu Y L,Zhu L,et al.Adaptor FADD is recruited by RTN3/HAP in ER-bound signaling complexes[J].Apoptosis,2006,(11):1923-1932.
[21] Zhang J K,Cado D,Chen A,et al.Fas-mediated apoptosis and activation-induced T-cell proliferation are defective in mice lacking FADD/Mort1[J].Nature,1998,392(19):296-300.
[22] Kim N,Alan W H,Mary L B,et al.A dominant interfering mutant of FADD/MORT1enhances deletion of auto reactive thymocytes and inhibits proliferation of mature T lymphocytes[J].The EMBO Journal,1998,17(3):706-718.
[23] Ishaq A V,Todd F,Shannon D,et al.Pax2regulates a FADD-dependent molecular switch that drives tissue fusion during eye development[J].Human Molecular Genetics,2012,21(10):2357-2369.
[24] Christopher P D,Andrew O,Ricardo W,et al.Survival Func-tion of the FADD-CASPASE-8-cFLIPL Complex[J].Cell,2012,1(5):401-407.
[25] Yeh W C,Pompa J L,McCurrach M E,et al.FADD:essential for embryo development and signaling from some,but not all,inducers of apoptosis[J].Science,1998,279(5358):1954-8.
[26] 刘 辉,姚咏明.细胞内炎症信号通路交汇作用研究进展[J].中国病理生理杂志,2005,21(8):1607-1613.
[27] Bruno S,Pedro R,Serge L.Toll-like receptors'two-edged sword:when immunity meets apoptosis[J].2007,37(12):3311-3318.
[28] Welz P S,Wullaert A,Vlantis K,et al.FADD prevents RIP3-mediated epithelial cell necrosis and chronic intestinal inflammation[J].Nature,2011,477(15):330-335.
[29] Marion C B,Daniela P,Welz PS,et al.The Adaptor Protein FADD Protects Epidermal Keratinocytes from Necroptosis In Vivo and Prevents Skin Inflammation[J].Immunity,2011,35(4):572-582.
