水分胁迫对不同品种小麦幼芽抗氧化酶活性的影响
2013-09-15张述义刘玲玲
张述义,刘玲玲
(1.山西省农业科学院旱地农业研究中心,山西太原030006;2.山西省农业科学院小麦研究所,山西临汾041000)
植物体在水分胁迫逆境条件下,质膜膜脂的物理状态改变可能是植物感受渗透胁迫的原初响应[1]。植物在逆境下通常伴随大量活性氧自由基的产生,引起细胞中产生活性氧自由基与活性氧自由基的清除系统失衡,如果不及时清除将会造成细胞膜和一些大分子物质的破坏[2],膜结构中含有的不饱和脂肪酸的类脂易受活性氧自由基攻击,导致膜脂过氧化作用发生,脂质过氧化产物MDA含量增加,使质膜的选择透性丧失,电解质与一些小分子有机物质外渗,最终导致植物伤害发生[3]。植物体内的抗氧化系统是植物细胞对抗氧化胁迫的关键因素,其中,超氧化物歧化酶( SOD)、过氧化氢酶( CAT)、过氧化物酶( POD)是抗氧化酶系统中3种重要的保护酶,是生物体内重要的活性氧自由基清除剂,可以清除机体内多余的活性氧自由基,解除活性氧自由基对生物体的毒性[4],维持植物体内活性氧自由基产生和淬灭的动态平衡,防止膜脂过氧化。
干旱是影响小麦播种出苗的主要因素,尤其在小麦幼芽期,干旱逆境下成苗的关键不在于萌发,而主要在于萌发后的幼芽伸长[5-6]。为了解不同肥水类型小麦品种幼芽抗氧化酶系统对水分胁迫逆境的反应,本研究选用晋麦47号(冬性半冬性、旱地品种)和舜麦1718(冬性半冬性、水地品种)2个不同肥水类型的小麦品种,在发芽期进行水分胁迫处理,对幼芽中的抗氧化酶SOD,POD,CAT活性以及MDA含量和蛋白质含量进行了分析,以期为旱地栽培中选用高产小麦品种提供理论依据。
1 材料和方法
1.1 试验材料
供试材料晋麦47号、舜麦1718小麦品种,分别由山西省农业科学院小麦研究所和棉花研究所提供。
1.2 材料培养
温汤浸泡小麦种子30 min,之后沥出种子,(25±1)℃清水浸泡24 h后,再将种子沥出,挑选萌动势一致的种子,摆入培养皿(Ф10 cm)中,加入混合溶液( 0.1 mmol/LKCl,0.1 mmol/LCaCl2,0.1 mmol/L MgCl2,0.5 mmol/L NaCl,0.2 mmol/L Na20.3mmol/LMES),( 25±1)℃继续培养 5d。
1.3 试验设计
将培养5 d的小麦培养皿内水分吸净,加入0(对照),19.2%(水分胁迫处理)聚乙二醇(PEG6000)溶液进行处理,24 h后取样,测定SOD,POD,CAT酶活性和MDA及蛋白质含量,重复3次。
1.4 测定项目及方法
SOD活性测定,以反应抑制氮蓝四唑(NBT)光化还原50%的酶量为一个酶活单位,U/(g·h)[7];POD活性采用愈创木酚法测定,以每分钟内吸光度(变化0.1为一个酶活单位,U/(g·min)[8];CAT活性采用紫外分光光度法测定,以1 min内A240减少0.1的酶量为一个酶活单位,U/(g·min)[9];MDA含量采用硫代巴比妥酸法测定[7];蛋白质含量采用考马斯亮蓝染色法测定[9]。
1.5 数据分析
采用Excel对试验数据进行统计分析。
2 结果与分析
2.1 水分胁迫后不同小麦品种幼芽MDA含量的变化
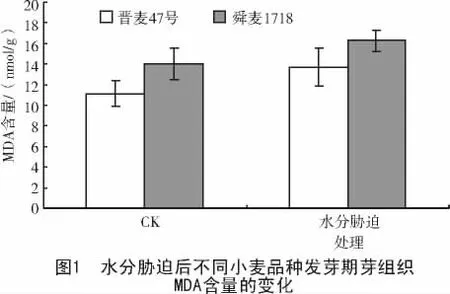
由于水分胁迫处理可以引起植物体细胞内有害自由基增多,引发膜中不饱和脂肪酸过氧化反应发生。MDA是膜脂过氧化作用的主要产物,能与酶蛋白发生链式反应聚合,使膜系统变性,造成膜脂流动性降低,影响膜结合酶的功能,膜脂过氧化产物MDA含量的增加,会造成膜伤害,则MDA含量反映膜脂质过氧化程度[10-11]。
试验结果(图1)表明,在正常供水的对照中,晋麦47号幼芽中MDA含量低于舜麦1718;水分胁迫处理24 h后,晋麦47号和舜麦1718的MDA含量均升高,与对照相比,晋麦47号的MDA含量增加了23.26%,舜麦1718增加了15.9%。说明舜麦1718幼芽中的过氧化代谢产物积累相对少,对膜损伤程度较小。
2.2 水分胁迫后不同小麦品种幼芽SOD酶活性的变化
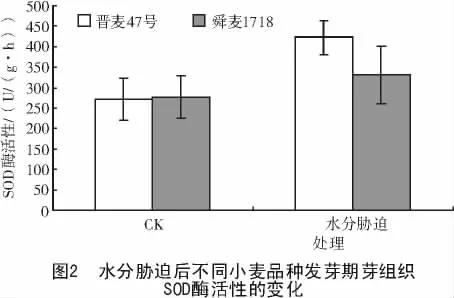
SOD是植物体内一个关键的抗氧化酶,可通过Haber-weiss反应清除植物体内多余的超氧根阴离子,将2个超氧自由基发生歧化反应形成水和氧。从图2可以看出,对照的幼芽中晋麦47号与舜麦1718的SOD酶活性相近,分别为271.25,277.16 U/(g·h)。水分胁迫处理24 h后,2个小麦品种的SOD酶活性均上升,晋麦47号上升幅度大于舜麦1718,二者的SOD酶活性分别增加了55.83%,19.58%,可见,在水分胁迫下,晋麦47号幼芽以提高SOD酶活性淬灭超氧阴离子的能力大于舜麦1718。
2.3 水分胁迫后不同小麦品种幼芽CAT酶活性的变化
过氧化氢(H2O2)是植物生长过程中的代谢产物,逆境会导致过量的过氧化氢产生,对机体造成损害。CAT主要功能是分解植物组织中氧代谢中产生的H2O2以及有机过氧化物 ROOH,快速地转化为其他无害较小的物质,可以改善质膜氧化损害程度,CAT酶活力越高,H2O2以及ROOH增加的比例越小。从图3可以看出,对照的舜麦1718幼芽中CAT酶活性高于晋麦47号。经过24 h水分胁迫处理后,2个小麦品种的CAT酶活性均提高。晋麦47号的CAT酶活性比对照增加了163.84%,舜麦1718的CAT酶活性比对照增加了11.69%,可见,晋麦47号幼芽中的过氧化氢酶活性对水分胁迫反应非常敏感。

2.4 水分胁迫后不同小麦品种幼芽POD酶活性的变化
POD在植物体内的主要作用也是清除H2O2,并将其分解为O2和H2O,其活性升高可以减轻活性氧对细胞膜的伤害。由图4可知,晋麦47号幼芽POD酶活性高于舜麦1718。水分胁迫处理24 h后,晋麦47号的POD酶活性比对照增加92.52%,舜麦1718比对照增加59.25%。

2.5 水分胁迫后不同小麦品种幼芽蛋白质含量的变化
可溶性蛋白质是植物渗透调节物质之一,在维持生物膜结构功能稳定性、调节离子运输、抑制蛋白酶活性等方面起重要作用。很多与植物耐旱性相关的逆境诱导蛋白可能在不同小麦品种之间存在较一致的调节机制。由图5可知,舜麦1718幼芽中蛋白质含量高于晋麦47号,水分胁迫处理24 h后,晋麦47号蛋白质含量与对照相比增加118.84%,舜麦1718增加了23.39%,与酶活性增加趋势一致。

2.6 水分胁迫后不同小麦品种幼芽抗氧化酶活性总体的变化
从图6可以看出,小麦发芽期,水分胁迫处理24 h后,晋麦47号幼芽中3个抗氧化酶活性增加百分率的总和大于舜麦1718,晋麦47号的抗氧化酶活性对水分胁迫敏感。晋麦47号以蛋白质为主要渗透调节物质,以增强抗氧化酶活性,来抵御水分胁迫的危害。

3 结论与讨论
晋麦47号是一个抗旱、高产的优良旱地冬小麦品种,舜麦1718是一个高产优良的水地冬小麦品种[12-15],晋麦47号幼芽在水分胁迫处理24 h后,3个抗氧化酶活性增强,表现为清除体内活性氧自由基能力加强。SOD在清除超氧化物阴离子自由基的过程中,会产生H2O2,对细胞造成伤害。过氧化氢酶(CAT)具有清除过量H2O2的作用,与SOD和POD一起保护膜系统免受自由基的伤害。舜麦1718幼芽中的3个抗氧化酶对水分胁迫反应没有晋麦47号那么敏感,其抗氧化酶活性增强水平较低,活性氧代谢活动相对平稳,可能是由于自身具有其他抑制或清除过氧化物积累的能力,但这还有待于进一步研究。
在水分胁迫下,不同小麦品种的蛋白质含量都有所增加,这可能是由于幼芽蛋白质组分发生了变化,有新蛋白亚基产生,这些蛋白可能是一些结构蛋白,也可能是一些渗透调节物质或酶,会在植物受到伤害时起保护作用[16]。在水分胁迫下,晋麦47号蛋白质含量增长最大,在逆境诱导蛋白的表达方面优于舜麦1718。
不同小麦品种幼芽,在水分胁迫下通过不同的生理代谢过程和代谢水平,维持体内活性氧产生和淬灭的动态平衡。旱地品种晋麦47号主要是依靠提高SOD,POD,CAT酶活性和蛋白质含量来抵御干旱的危害;水地品种舜麦1718虽然抗氧化酶活性变化幅度小,但从膜脂过氧化状态分析,有研究显示,抗旱性较强的品种MDA含量增加幅度低于抗旱性弱的品种[17],水分胁迫下舜麦1718的MDA含量增加小于晋麦47号,由于MDA含量变化率小,质膜的损伤程度较轻,表明水地小麦品种舜麦1718可能具有适合旱地栽培的潜力,植物受到干旱胁迫后,可通过启动体内的一系列反应来抵御干旱[18],2个品种抵御水分胁迫危害可以通过不同途径来实现,其生理代谢特点还需要从多方面进一步深入探讨。
[1]邱全胜.渗透胁迫对小麦根质膜膜脂物理状态的影响[J].植物学报,1999,41( 2):161-165.
[2]高银.植物抗逆机制与基因工程研究进展[J].内蒙古农业科技,2007( 5):75-78.
[3]陈少裕.膜脂过氧化与植物逆境胁迫 [J].植物通报,1989,6( 4):211-217.
[4]贺杰,王伟,胡海燕.小麦种子活力与其保护酶活性关系的研究[J].天津农业科学,2012,18( 2):17-19.
[5]山仑,郭礼坤.逆境成苗生态生理研究Ⅰ.春播谷类作物成苗期间的抗旱性及其需水条件 [J].作物学报,1984,10(4):257-263.
[6]杨美红,贾俊仙,李国锋.水分胁迫对不同耐旱性品种小麦幼芽、幼苗的保护酶和淀粉酶活性的影响 [J].山西农业科学,2001,29( 1):45-48.
[7]中国科学院上海植物生理研究所.现代植物生理学实验指南[M].北京:科学出版社,1999.
[8]龚富生,张嘉宝.植物生理学实验[M].北京:气象出版社,1995.
[9]邹琦.植物生理学实验指导[M].北京:中国农业出版社,2000.
[10]山仑,陈培元.旱地农业生理生态基础[M].北京:科学出版社,1998:1-18.
[11]王宝山.生物自由基与植物膜伤害 [J].植物生理学通讯,1988( 2) :2-16.
[12]孙来虎,李秀绒,柴永峰,等.晋麦47号产量结构特点与高产栽培技术[J].耕作与栽培,2003( 5):48-49.
[13]董孟雄,李秀绒,柴永峰,等.旱地小麦新品种:晋麦47号[J].麦类作物学报,2001,21( 1):98.
[14]张运校,樊立强,刘彦军,等.国审优质小麦新品种舜麦1718高产栽培技术[J].种子科技,2012( 3):28-29.
[15]潘幸来,史引红,王永杰,等.舜麦1718小麦新品种选育报告[J].小麦研究,2007,28( 4):26-36.
[16]戴明,邓西平,杨淑慎,等.水分胁迫对不同基因型小麦幼芽蛋白质表达和某些生理特性的影响 [J].应用生态学报,2009,20( 9):2149-2156.
[17]张文英,智慧,柳斌辉.谷子孕穗期一些生理性状与品种抗旱性的关系[J].华北农学报,2011,26( 3):128-133.
[18]荣少英,郭蜀光,张彤.干旱胁迫对甜高粱幼苗渗透调节物质的影响[J].河南农业科学,2011,40( 4):56-59.
