低磷胁迫对不同油茶优良无性系酶活性的影响1)
2013-08-08陈隆升陈永忠彭邵锋王湘南杨小胡
陈隆升 陈永忠 王 瑞 彭邵锋 王湘南 杨小胡
(湖南省林业科学院,长沙,410004)
植物在长期的遗传变异过程中,体内形成了许多对养分及环境胁迫的适应性机制,其中酶系统的变化是主要的适应性反应之一。许多研究表明,作为植物体的保护酶系统,超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)相互协调,可以在一定程度上减缓活性氧对植物细胞造成的伤害[1-2]。植物在逆境条件下往往具有较高的保护酶系统,其中根系分泌到生长介质中的酸性磷酸酶活性(APase)显著升高[3-5],通过水解介质中的无效态有机磷的磷酸酯键而释放PO-43;通过增加叶片和根系组织中APase活性,重复利用植株体内有机磷来解除磷营养胁迫[6]。目前,对油茶在磷胁迫环境中研究较少,主要集中于低磷胁迫生理生化机制的探讨方面[7-8]。油茶(Camellia oleifera Abel.)广泛分布在我国南方14个省区,有上百个品种及类型[9-10],这些品种及类型在长期的系统发育过程中形成了不同的地理生态类型,具有不同的营养遗传性状,为磷高效型油茶品种的筛选提供了丰富的材料。因此,笔者通过研究不同油茶优良无性系受低磷胁迫影响后各抗氧化酶活性及膜脂过氧化的变化情况,以探讨植物耐低磷能力与保护酶系统的关系,为油茶磷高效型品种的选育和磷素利用提供理论依据。
1 材料与方法
供试材料:试验材料为1年生油茶无性系苗,5个油茶优良无性系为湘林56、湘林4、湘林81、湘林67、湘林69,其苗高基本在15 cm左右。
试验设计:采用1/2Hoagland完全营养液培养植物材料,设无磷(P0,0 mmol/L)、低磷(P1,0.05 mmol/L)、正常磷(P2,0.50 mmol/L)3 个磷浓度梯度,磷由KH2PO4提供,具体详见陈隆升的方法[8]。将油茶苗根系土壤用清水洗干净,蒸馏水清洗后,移植到装有河沙(用稀盐酸浸泡、蒸馏水反复冲洗)的塑料桶(直径20 cm,高30 cm)中培养。每桶3棵幼苗,每3 d浇1次200 mL的营养液。中间根据失水情况补充一次200 mL的蒸馏水,每个处理7盆,共21株苗。培养环境为温室塑料大棚,棚内平均温度30℃(7—9月)和22℃(10—11月)左右,相对湿度60%以上,光照时间为12~13 h。培养时间为2010年7月15日—11月22日,分别于处理后的45、75、105 d采取植株叶片,于超低温冰箱中保存。
试验指标测定方法:SOD、POD、CAT测定参考张志 良 和 邹 琦 的 方 法[11-12];脯 氨 酸 和 丙 二 醛(MDA)测定参考中国科学院上海植物生理研究所的方法[13]。叶片的酸性磷酸酶活性测定参考林启美等的叶饼法[14]。所有分析均3次重复。
数据处理与分析:应用SAS 9.0和Excel进行相关分析。
2 结果与分析
2.1 低磷胁迫对不同油茶优良无性系保护酶活性的影响
对SOD活性的影响:从表1中可以看出,随培养介质磷水平的下降,油茶无性系叶片SOD活性显著增加,磷水平间差异达到极显著水平(P<0.01),其活性增幅为7.40% ~30.87%(P0与P2比较)和2.15% ~18.61%(P1与 P2比较),具有明显的趋势性。进一步地统计分析表明,不同磷水平下各参试油茶无性叶片SOD活性差异显著(P<0.05),在无磷(P0)和低磷(P1)下,均以湘林67最高;与正常磷(P2)相比,湘林69的SOD活性增幅最大,分别增加了30.87%和18.61%,各无性系的增幅大小依次为湘林69、湘林81、湘林67、湘林4、湘林56。
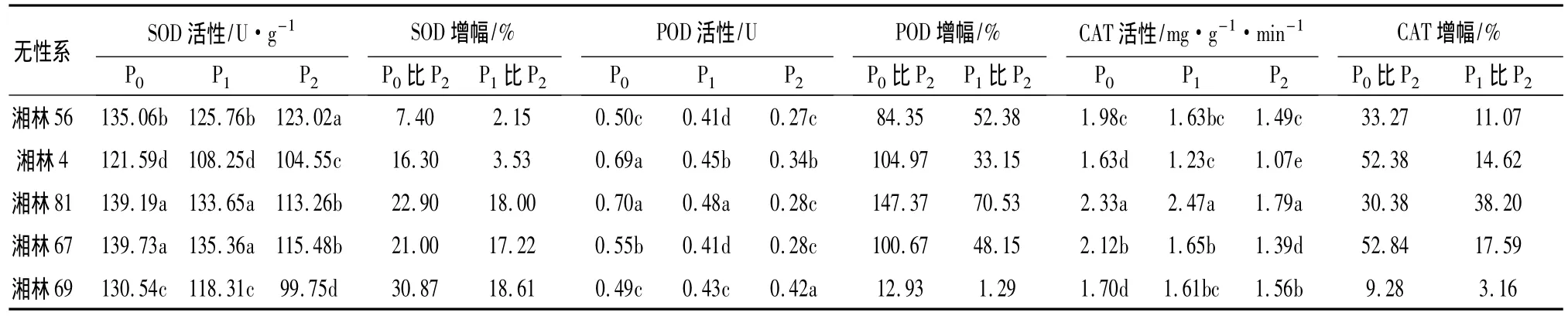
表1 低磷胁迫对不同油茶无性系叶片保护酶活性的影响
对POD活性的影响:从表1中可以看出,随培养介质磷水平的下降,油茶无性系叶片POD活性显著增加,磷水平间差异达到极显著水平(P<0.01),其活性增幅为12.93% ~147.37%(P0与P2比较)和1.29% ~70.53%(P1与P2比较)。方差分析表明,不同磷水平下各参试油茶无性系叶片POD活性有显著异差(P<0.05),在无磷(P0)和低磷(P1)下,均以湘林81最高;与正常磷(P2)相比,湘林81的POD活性增幅最大,分别增加了147.37%和 70.53% 。
对CAT活性的影响:从表1中可以看出,随培养介质磷水平的下降,油茶无性系叶片CAT活性显著增加,磷水平间差异达到极显著水平(P<0.01),其活性增幅为9.28% ~52.84%(P0与P2比较)和3.16% ~38.2%(P1与 P2比较)。方差分析表明,不同磷水平下各参试油茶无性叶片CAT活性有显著异差(P<0.05),在无磷(P0)和低磷(P1)下,均以湘林81最高;与正常磷(P2)相比,无磷胁迫下(P0)湘林67的CAT活性增幅最大,为52.84%,低磷胁迫下(P1)则以湘林81的幅最大,为38.2%,其次是湘林 67,为 17.59%。
2.2 低磷胁迫对不同油茶优良无性系 MDA和APase活性的影响
从表2中可以看出,在无磷(P0)和低磷(P1)下,各无性系的MDA质量摩尔浓度均显著(P<0.05)高于正常磷(P2),其活性增幅为2.62% ~26.15%(P0与P2比较)和1.33% ~16.20%(P1与 P2比较)。方差分析表明,不同磷水平下各参试油茶无性系叶片丙二醛质量摩尔浓度有显著异差(P<0.05),在无磷(P0)和低磷(P1)下,均以湘林56最高,湘林81最低;无磷胁迫下(P0)湘林69的MDA质量摩尔浓度增幅最小,为2.62%(P0与 P2比较),低磷胁迫下(P1)则以湘林67的增幅最小,为1.33%,其次是湘林69,为3.95%(P1与P2比较)。在缺磷处理(P0、P1)叶片APase活性均极显著(P<0.01)高于正常供磷处理(P2),叶片APase活性增幅为23.44% ~97.30%(P0与 P2)和 15.06% ~47.48%(P1与 P2比较)。随胁迫程度的加重,酶活性增强趋势明显。这表明低磷胁迫下,油茶无性系植株体内的APase活性也会发生变化,以使植株体内仅有的磷得以多次重复利用。方差分析结果表明,无磷(P0)、低磷(P1)胁迫下,不同油茶无性系叶片APase活性均达到显著差异(P<0.05)。在无磷胁迫下,叶片APase活性以湘林67最强,湘林69次之,低磷胁迫水平下,叶片APase活性以湘林67最强。
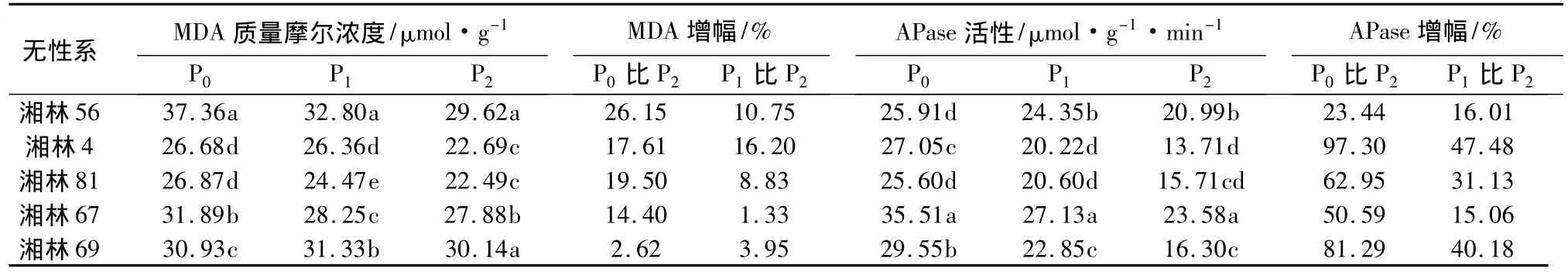
表4 低磷胁迫对不同油茶无性系叶片MDA和APase的影响
3 结论与讨论
POD、SOD、CAT等酶活性的变化已广泛应用于植物抗逆性研究[15-17],作为植物细胞的保护酶系统,SOD、CAT和POD在清除活性氧过程中发挥着巨大的作用。本研究发现,与正常供磷(P2)相比,在无磷(P0)和低磷(P1)胁迫下各油茶无性系的保护酶活性均显著提高。在无磷处理(P0)下,SOD活性增幅为7.40% ~30.87%,POD 活性增幅为12.93% ~147.37%,CAT 活性增幅为9.28% ~52.84%;低磷处理(P1)下,SOD 活性增幅为2.15% ~18.61%,POD活性增幅为1.29% ~70.53%,CAT 活性增幅3.16% ~38.20%。研究进一步证实了保护酶系统活性的提高是油茶适应低磷胁迫的重要生理反应机制之一。
保护酶系统的变化及其对活性氧的清除可以认为是植物耐低磷胁迫的生理机制之一[16-17]。本研究发现在无磷(P0)和低磷(P1)胁迫下5个油茶优良无性系在保护酶活性变化方面存在显著差异,与正常磷(P2)相比,无磷(P0)和低磷(P1)胁迫下湘林69的SOD活性增幅最大,分别增加了30.87%和18.61%;湘林81的POD活性增幅最大,分别增加了147.37%和70.53%;无磷胁迫下(P0)湘林67的CAT活性增幅最大,为52.84%,低磷胁迫下(P1)则以湘林81的增幅最大,为38.20%,表明湘林81、湘林67、湘林69这3个无性系能快速调整保护酶系统活性从而增强了植株的抗低磷能力。
MDA是反映及植物受害后自我修复能力的重要指标之一,利用MDA质量摩尔浓度和(或)其它生理生化指标进行作物抗性的评价与鉴定,这在水稻、甘蔗等作物的抗旱性鉴定中已有成功的应用[18-19]。本研究发现,与正常供磷(P2)相比,在无磷(P0)和低磷(P1)下,MDA质量摩尔浓度均以湘林56最高,湘林81最低,无磷胁迫下(P0)湘林69的MDA质量摩尔浓度增幅最小,为2.62%,低磷胁迫下(P1)则以湘林67的增幅最小,为1.33%,其次是湘林69,为3.95%,表明湘林56在受到低磷胁迫时其膜的过氧化水平和膜结构的受害程度大,而湘林81、湘林67、湘林69受害程度相对较小,这是由于这3个无性系保护酶活性的大幅度提高,及时清除自由基,这也进一步验证了这3个无性系在受到低磷胁迫时具备较强的自我修复能力。
APase分泌量的提高是忍耐缺磷胁迫的适应性反应,也是作为耐低磷品种筛选的一个生化指标,本研究发现在无磷和低磷胁迫下,湘林67和湘林69具有较高的叶片APase活性。同时在前期的研究中,笔者发现在无磷和低磷胁迫下,湘林67、湘林81和湘林69根系内部和外部的APase活性同样较高,同时湘林67和湘林69在磷缺乏的条件下仍然具有较高的光合速率[8],因此初步认定这2个无性系为较耐低磷的无性系,但其耐低磷的能力、稳定性及与产量的关系等均有待进一步的研究。
[1] 张玉霞,李志刚,张玉玲,等.缺磷胁迫对大豆膜脂过氧化及保护酶活性的影响[J].中国农学通报,2004,20(4):196-197,208.
[2] 余叔文,汤章城.植物生理与分子生物学[M].上海:科学出版社,1999:375-380.
[3] Song J Y,Shawn M K.Induction of maize acid phosphatase activities under phosphorus starvation[J].Plant and Soil,2001,237(1):109-115.
[4] Tian Jiang,Liao Hong,Wang Xiurong,et al.Phosphorus starvation-induced expression of leaf acid phosphatase isoforms in soybean[J].Acta Botanica Sinica,2003,45(9):1037-1042.
[5] Baldwin J C,Karthikeyan A S,Raghothama K G.Aphosphorus starvation induced novel acid phosphatase from tomato[J].Plant Physiol,2001,125:728-737.
[6] 梁霞,刘爱琴,马祥庆,等.磷胁迫对不同杉木无性系酸性磷酸酶活性的影响[J].植物生态学报2005,29(1)54-59.
[7] 陈隆升,陈永忠,彭邵锋,等.油茶对低磷胁迫的生理生化效应研究[J].林业科学研究,2010,23(5):782-786.
[8] 陈隆升,陈永忠,王瑞,等.磷胁迫对不同油茶优良无性系Apase活性的影响[J].中国农学通报,2011,27(31):58-63.
[9] 陈永忠.油茶栽培技术[M].长沙:湖南科学技术出版社,2008:28-30.
[10] 庄瑞林.中国油茶[M].2版.北京:中国林业出版社,2008:56-89.
[11] 张志良.植物生理学实验指导[M].北京:高等教育出版社,1990:120-121.
[12] 邹琦.植物生理生化实验指导[M].北京:中国农业出版社,1995:68-70.
[13] 中国科学院上海植物生理研究所.现代植物生理学实验指南[M].上海:上海科学技术出版社,1999:305-306.
[14] 林启美,黄德明.应用酸性磷酸酶进行番茄磷素诊断[J].华北农学报,1991,6(2):78-83.
[15] 王伟,曹敏建,王晓光,等.低钾胁迫对不同钾营养效应型大豆保护酶系统的影响[J].大豆科学,2005,24(2):101-105.
[16] 刘厚诚,邝炎华,陈日远.缺磷胁迫下长豇豆幼苗膜脂过氧化及保护酶活性的变化[J].园艺学报,2003,30(2):215-217.
[17] 潘晓华,刘水英,李锋,等.低磷胁迫对不同水稻品种叶片膜脂过氧化及保护酶活性的影响[J].中国水稻科学,2003,17(1):57-60.
[18] 程建峰,潘晓云,刘宜柏,等.水稻抗旱性鉴定的形态指标[J].生态学报,2005,25(11):3117-3125.
[19] 高三基,罗俊,张华,等.甘蔗抗旱性生理生化鉴定指标[J].应用生态学报,2006,17(6):1051-1054.
