IL-10在Plasmodium yoelii 17XL和Plasmodium chabaudi AS疟原虫混合感染宿主病理损伤中的作用研究
2013-07-23曹雅明蔡连顺苏菊香
陈 光,曹雅明,刘 蕾,蔡连顺,毕 胜,苏菊香,代 月
(1.佳木斯大学基础医学院寄生虫学教研室,黑龙江佳木斯 154007;2.中国医科大学基础医学院免疫学教研室,辽宁沈阳 110001;3.佳木斯大学基础医学院病理生理学教研室,黑龙江佳木斯 154007)
疟疾是疟原虫通过媒介昆虫蚊传播的人类最为严重的寄生原虫感染性疾病。每年全世界约有655万人感染疟疾,其中将近100万人死亡,绝大多数为5岁以下的儿童[1-2]。新的研究证实,不同种属疟原虫混合感染在发展中国家是一种普遍现象。尤其在疟疾流行区,恶性疟原虫同时伴随间日疟原虫或三日疟原虫混合感染约占疟疾总感染人数的30%以上[3-5]。疟疾的流行已成为全球性最严重的三大公共卫生问题之一,给全球尤其是发展中国家带来了严重的健康危害和沉重的经济负担[6]。到目前为止,无有效疫苗能有效地预防疟疾感染[7]。因此,研制安全、价廉和有效的疟疾疫苗是人类控制乃至根除疟疾的重要途径,也是当今世界迫切需要解决的重点课题。而疟原虫感染宿主机体应答的免疫学、分子生物学等综合基础研究是其重要前提。疟疾与其他大多数感染性疾病一样,需要通过免疫效应机制对其控制和消除。同时,由于过强的炎症反应引起脑疟等免疫病理损伤也是这一疾病的突出特征。IL-10是一种抗炎性细胞因子,由巨噬细胞(MØ)、树突状细胞(DC)、B细胞以及CD4+和CD8+T细胞亚类分泌。IL-10通过抑制MØ和DCs的功能,限制了Th1/Th2型免疫应答的发生;DC或MØ分泌的IL-10能诱导自然调节性T细胞(nTreg)产生IL-10,从而阻止了宿主病理损伤的出现,但却允许病原体长期逃避免疫系统控制并引起长期(通常无症状)感染[8]。目前,IL-10在疟疾感染尤其是混合感染中作用的研究尚未深入展开。有限的研究结果显示,一种疟原虫单独感染时IL-10是由自然调节性T细胞(nTreg)、Th1/Tr1、DC和MØ分泌的,其主要作用地位体现在抑制Th1应答,减少IL-12和IFN-γ的分泌;通过抗体中和或基因敲除,发现IL-10-/-小鼠TNF-α、IFN-γ水平增高,疟原虫清除能力增强,死亡率升高,发生免疫病理损伤/脑疟的机率明显增加[8-10]。因此,本实验拟利用P.y17XL、P.cAS单独感染和P.y17XL+P.cAS混合感染鼠疟模型,明确IL-10在疟疾混合感染过程中的作用地位,这一研究无疑将为疟疾等感染性疾病的有效控制提供新的理论依据。
1 材料与方法
1.1 材料
1.1.1 实验动物、疟原虫 6~8周龄、雌性DBA/2小鼠由中国医学科学院实验动物研究所提供(许可证编号:SCXK京200420001);P.y17XL和P.cAS(日本爱媛大学分子寄生虫学教研室惠赠)。
1.1.2 主要试剂 SYBR GREEN quantitative kit(Quigen,Germany);IL-10R Ab(clone 1B1.3a;Biolegend company)。
1.2 方法
1.2.1 实验动物感染及分组 DBA/2小鼠分为正常对照组、P.y17XL单独感染组、P.y17XL和P.cAS疟原虫混合感染组和P.cAS单独感染组,9只/组,分别经腹腔感染1×106P.y17XL、P.cAS和P.y17XL+P.cAS(1∶1)寄生的红细胞,感染不同时间的小鼠经尾静脉采血,制备薄血膜,Giemsa染色,光学显微镜镜检疟原虫,计数红细胞感染率。
1.2.2 实时定量PCR检测虫负荷 于感染第后3、5、8、10、12和19天分别尾静脉取血100 μL,肝素抗凝后短暂离心,采用高纯度DNA提取试剂盒抽提DNA;根据2种疟原虫的MSP-1基因序列,设计特异性引物:P.y17XL MSP-1 F5'-ACCATCAACATCTACACCAGCAC-3',R5'-CAGGCATATCCTCTTCACTTTCT-3',扩增长度为 110 bp;P.cAS MSP-1 F5'-GTAACATC ACAACCAGCATCAAC-3',R5'-CGACAAACATTTCGTCTTCACTT-3',扩 增 长度为100 bp。
1.2.3 脾细胞培养 分别于感染后第0、1、3、5、8、10、12和 15天,无菌取出小鼠脾脏,用RPMI1640培养液制备浓度为1×107个/mL脾细胞悬液,于24孔培养板中培养48 h,收集培养上清,-80℃保存。
1.2.4 细胞因子的检测 用双抗体夹心ELISA试剂盒检测脾细胞培养上清中的IL-10水平,按试剂盒说明操作,酶标仪测定450 nm处吸光度(A)值。以试剂盒提供的标准品测定结果绘制标准曲线,应用SoftMax Pro 4.3.1LS软件分析,计算细胞因子含量(pg/mL),结果以±s表示。
1.2.5 统计学处理 应用SPSS 11.5统计学分析软件,双尾t检验分析比较各组间的统计学差异。
2 结果与分析
2.1 疟疾混合感染小鼠原虫血症水平及生存率
P.cAS感染小鼠原虫血症水平迅速持续升高,感染后第9、10天达峰值约33.17±10.89%,感染小鼠全部死亡;而P.y17XL感染小鼠全部存活,感染后3 d和9 d原虫血症水平仅为1.66±0.4%和24.9±7.1%,随后迅速下降;相比P.y17XL+P.cAS混合感染小鼠的原虫血症水平也于感染后9 d达峰值约34±5.5%,小鼠生存率为100%(见图1A、1B)。
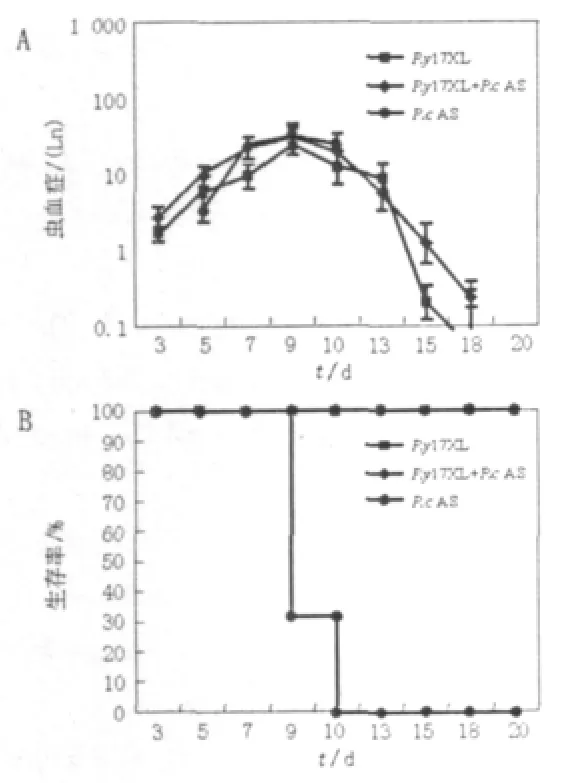
图1 疟原虫感染小鼠不同时间原虫血症水平(A)及生存率(B)Fig.1 Parasitemia(A)and survival rate(B)of P.y17XL,P.y17XL+P.c AS and P.c AS infection in DBA/2 mice in different time after infection
2.2 疟疾混合感染小鼠虫负荷水平检测
同等数量的P.y17XL和P.cAS疟原虫混合后腹腔注射DBA/2小鼠,RTQ-PCR检测感染小鼠体内2种疟原虫虫负荷水平。小鼠混合感染后3~12 d,P.y17XL 占优势;而感染后 15~19 d,P.cAS占优势(见图2)。
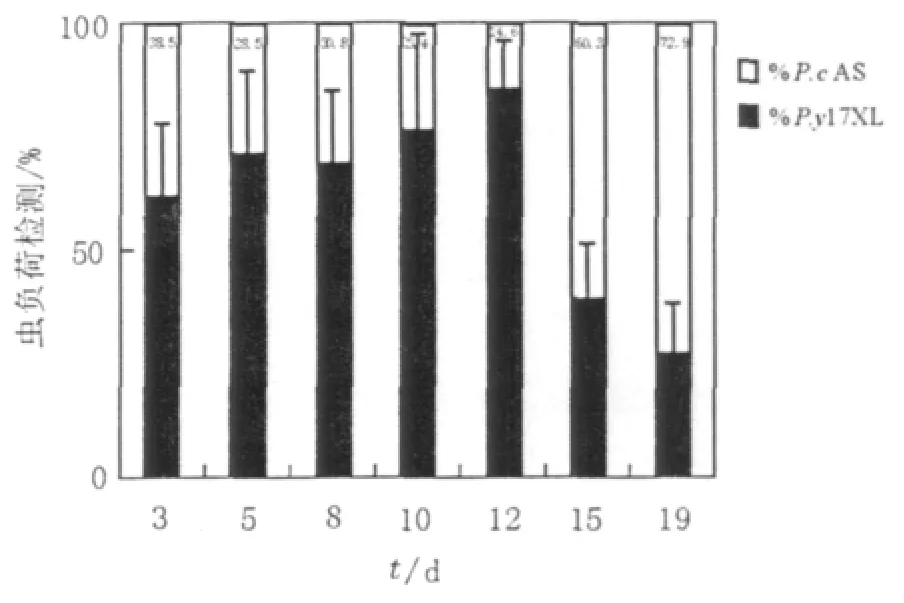
图2 疟疾混合感染小鼠不同时间疟原虫虫负荷水平Fig.2 Quantitative PCR analysis of DBA/2 miceco-infected with P.y17XL and P.cAS.Proportions of each strain measured in mixed-infections mice during the different time point after infection
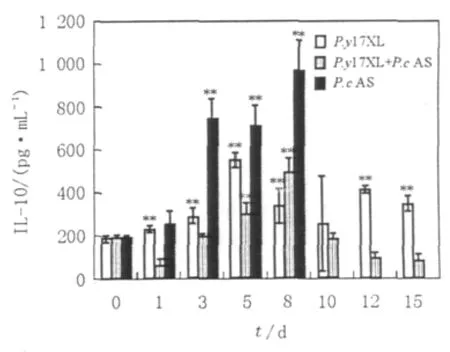
图3 混合感染小鼠感染后不同时间脾细胞上清中细胞因子IL-10水平检测Fig.3 Level of IL-10 in spleen cell supernatants during different infections in different time point
2.3 疟疾混合感染小鼠IL-10水平变化
混合感染小鼠IL-10水平于感染后第1~3天无明显变化,感染后第8天达峰值,随后迅速下降,于感染后第10天下降至正常水平;P.y17XL感染小鼠IL-10水平于感染后第1d既出现有意义的升高,第5天达峰值(P<0.01),此后虽有回落但仍维持其高水平;相比,P.cAS感染小鼠IL-10水平在感染后第3d出现迅速的有意义的升高(P<0.01),第8天达峰值,是同天混合感染小鼠IL-10水平的2倍(图3)。
3 讨论
疟疾与其他大多数感染性疾病一样,需要通过免疫效应机制对其控制和消除。同时,由于过强的炎症反应引起脑疟等免疫病理损伤也是这一疾病的突出特征。不断积累的研究数据表明:红内期阶段宿主通过强烈的促炎性免疫反应控制疟原虫增殖并促进感染红细胞清除,同时也严格调控免疫介导病理损伤(严重合并症)的程度[9,11],在疟疾感染过程中IL-10对促炎性应答具有重要的调节作用[12]。
本实验结果发现:P.y17XL单独感染和混合感染小鼠IL-10水平在感染后第5和8天分别达峰值,随后开始下降至正常水平,小鼠虫血症均达中等水平,存活率100%;相比P.cAS感染小鼠IL-10在感染后第3天突然出现高水平升高并且维持时间较长;于感染后第8天达峰值,是同天P.y17XL单独感染和混合感染小鼠IL-10水平的2倍,虫血症水平较高,小鼠全部死亡。由此提示,决定疟疾混合感染病理损伤严重性的应答效应明显受控于宿主免疫网络的调节,而IL-10可能正是在疟疾混合感染的发生和发展过程中处于举足轻重的作用地位。相关研究发现,细胞因子IL-10具有双向免疫调节活性,即负向调节作用及其免疫刺激作用。IL-10发挥不同的免疫学功效主要与IL-10蛋白的浓度、不同的微环境和病理条件及其不同的微环境、不同细胞和病理条件中IL-10受体表达格局的不同导致了IL-10不同的免疫学活性[13]。
同时发现,混合感染小鼠于感染后18 d自愈。实时定量PCR结果发现,混合感染小鼠,于感染后3~12 dP.y17XL增殖占优势,而感染后15~19 d则P.cAS增殖处于优势状态。相关的实验研究证明:与其它疟原虫虫株相比,P.cAS生长相对缓慢,针对生长速度不同的疟原虫,宿主免疫系统产生的免疫应答强度具有明显差异性[14];啮齿类动物混合感染模型证明:感染早期,P.y17XL能抑制P.cAS的生长[15];同时人们发现针对一种寄生虫感染造成的特有的致命性病理损伤能被同时感染的另一种寄生虫减轻,不同疟原虫虫株之间存在着拮抗作用[16]。
综上所述,本实验证实以IL-10为核心的免疫调节网络与疟疾感染过程中病理损伤密切相关。同时提示混合感染小鼠应答模式与P.y17XL感染小鼠的应答模式相同。上述问题的阐明不仅将进一步明晰疟疾免疫应答相关机制的本质,而且研制诱导物种间交叉免疫为靶向的疟疾疫苗新策略提供积极的指导意义,同时也为疟疾的免疫治疗手段开拓新的思路。
[1]World Health Organization[R].World Malaria Report,2012.
[2]Murray CJ,Rosenfeld LC,Lim SS,et al.Global malaria mortality between 1980 and 2010:a systematic analysis[J].Lancet,2012,379(9814):413-431.
[3]Imwong,M.,S.Pukrittayakamee,A.C.Grüner,et al.Practical PCR genotyping protocols forPlasmodium vivaxusingPvcsandPvmspl[J].Malar.J,2005,4(1):20.
[4]Mayxay M,Pukrittayakamee S,Newton PN,et al.Mixe-species malaria infections in humans[J].Trends Parssitol,2004,20(5):233-40.
[5]Zimmerman PA,Melotra RK,Kasehagen LJ,et al.Why we need to know more about mixedPlasmodiumspecies infections in humans[J]?Trends Parasitol,2004,20(9):440-447.
[6]Njau JD,Goodman CA,Kachur SP,et al.The costs of introducing artemisinin-based combination therapy:evidence from district-wide implementation in rural Tanzania[J].Malar J,2008,7(1):4.
[7]Pierce Susan K,Louis H.Miller.World Malaria Day 2009:What Malaria Knows about the Immune System That Immunologists Still Do Not[J].The Journal of Immunology,2009,182:5171-5177.
[8]Joss,A.,M.Akdis,A.Faith,et al.IL-10 directly acts on T cells by specifically altering the CD28 co-stimulation pathway[J].Eur.J.Immunol,2000,30(6):1683-1690.
[9]Artavanis-Tsakonas K,Tongren JE,Riley EM.The war between the malaria parasite and the immune system:immunity,immunoregulation and immunopathology[J].Clin Exp Immunol,2003,133(2):145-152.
[10]Couper KN,Blount DG,Riley EM.IL-10:the master regulator of immunity to infection[J].J Immunol,2008,180(9):5771-5777.Review.
[11]Black,J.Mixed infections withPlasmodium falciparumandP.malariaeand fever in malaria[J].Lancet,1994,343(8905):1095.
[12]Engwerda C,Belnoue E,Gruner AC,et al.Experimental models of cerebral malaria[J].Curr Top Microbiol Immunol,2005,297:103-143.
[13]周琳,周光炎,路丽明.IL-10的双向免疫调节作用[J].细胞与分子免疫学杂志,2012,28(10):1100-1102,1106.
[14]Cheesman S,Raza A,Carter R.Mixed strain infections and strain-specific protective immunity in the rodent malaria parasitePlasmodium chabaudi chabaudiin mice[J].Infection and Immunity,2006,74(5):2996-3001.
[15]Snounou,G.,T.Bourne,W.Jarra,et al.Assessment of parasite population dynamics in mixed infections of rodent plasmodia[J].Parasitology,1992,105(3):363-374.
[16]Tatiana Voza,Ana M.Vigário,Elodie Belnoue,et al.Species-Specific Inhibition of Cerebral Malaria in Mice Coinfected withPlasmodiumspp[J].Infection and Immunity,2005,73(8):4777-4786.