不同温度下雌雄葎草营养生长期的生长特性
2013-03-14刘金平
段 婧,刘金平
(西华师范大学生命科学院 西南野生动植物资源保护教育部重点实验室,四川 南充 637002)
葎草(Humulusscandens)又称拉拉秧、拉拉藤,为桑科葎草属雌雄异株植物,一年生或多年生草质藤本。主要生长在荒山荒坡及沟边、路旁、宅旁、荒废区域,单优势种群。由于其适应范围广、生长迅速、蔓延覆盖力强,在荒坡沟壑的绿化中有极大的潜力[1]。葎草可作鲜嫩饲草、青贮和干粉饲料及饲料添加剂等,增加禽畜对环境的适应能力,增强消化吸收机能[2-3]。葎草含葡萄糖苷、胆碱、鞣质、挥发油、天门冬酰胺、草酮等化学成分,营养物质和生物活性激素类物质,促进生长因子等成分,具有清热解毒、消於利尿、抗菌消炎、健胃抑菌、抑制病毒等功能,有利于人和动物的免疫力提高、疾病防治等[4]。同时,野生葎草为农田和园林绿化的重要杂草,加强对野生葎草资源的研究,探讨其生长、生理、生态特点,对杂草有效控制及“变废为宝”式开发利用具有重要意义。
由于雌雄性别的分株表达,雌雄异株植物在环境胁迫下维持种群稳定性的能力比其他植物更加薄弱,环境胁迫对雌、雄株光合作用能力、生物量分配、水分利用效率以及凋落物降解等方面的影响,是雌雄异株植物研究的热点问题[5],其中不乏温度对雌雄植物胁迫方面的研究[6-7]。近年来,对雌雄异株草本植物的研究报道远少于木本植物。对乔、灌木的研究结果表明,环境胁迫对雌雄个体的生理、生态和生化的影响,引起性别间在生长、形态、生殖、分布以及抗逆性等方面表现出明显的差异[8],从而引起种群性比组成逐渐发生变化,种群繁殖能力下降,最终导致种群所在的生态系统的组成、结构和功能发生改变。目前,关于葎草的研究主要集中在营养成分、药物成分及饲喂效果评价方面[9-11],对野生葎草种群的雌雄株形态、性比、生殖投入、种子活性等方面也进行了较为系统的研究[12-14]。本研究采用构件分析法,设置3个温度梯度,通过测定雌雄葎草营养生长期的根、茎、叶等构件数量与性状的变化,研究雌雄株的营养生长持续期、构件性状及生物量分配对温度响应的性别差异,以期为胁迫环境下葎草种群形成、适应度及种群扩繁等研究奠定基础,为雌雄异株植物资源保护利用、生态系统可持续发展及社会经济发展提供科学依据。
1 材料与方法
1.1试验材料与设计 试验在四川南充市进行,该区属典型的中亚热带湿润季风气候区,年均温17.4 ℃,最高温40.1 ℃,最低温-2.8 ℃,年日照时数1 266.7 h,平均有霜期仅13.7 d,年降水量1 020.8 mm。
以当地采集的野生葎草种子为材料,该植物种子难以区分性别,假定雌雄性比为1∶1。于2011年3月,采用高34 cm、口径50 cm的塑料花盆,将河沙∶腐殖土以1∶1均匀混合作为基质,按每盆10粒种子繁殖,共30盆。在25 ℃左右的室内培养,期间进行正常水分管理。3月下旬出苗后每盆按3株均匀定苗;1~2对真叶时,随机10盆分为1组,分别移入15、20、25 ℃的人工气候培养箱中,12 h光照/12 h黑暗进行培养,光照强度为1 300 lx,相对湿度为32%。
1.2测定指标与方法 营养生长期:从出苗到花序出现的持续时间。
叶的性状及数量: 相同温度处理组全部出现花序后,每组随机选取雌、雄各3株(雌株花序为穗状花序,雄株为圆锥花序),测定叶片叶长、叶宽、单叶面积(坐标低法)、总叶片数,3次重复。
茎的性状及数量:测叶性状的同时,测定茎的节间长、茎直径、最长茎长(直尺法)和分枝数,3次重复。
根的性状:测定茎叶的同时,出盆冲洗泥土后测定根长。
鲜质量与干质量:清洗根、茎、叶,用吸水纸吸干,分别称鲜质量后,装入信封,在105 ℃下烘至质量恒定,测定干质量。
生物量比例:各构件生物量(干质量)占单株总生物量(根、茎、叶的干质量之和)的百分比。
1.3数据分析 用SPSS 16.0软件进行双因素方差分析。
2 结果与分析
2.1温度对营养生长持续期和叶性状的影响 温度对葎草营养生长持续期出现显著影响(P<0.05)(表1)。在20 ℃处理下营养期最短,约为28 d,仅为25 ℃的营养生长持续期的一半左右。雌、雄株性别间差异显著,在20 ℃时雌株先于雄株完成营养生长,而15、25 ℃时雄株先于雌株完成营养生长。方差分析表明,在性别间、温度间及互作间,营养生长持续期均有显著差异。
葎草雌雄株叶宽和单叶面积受温度影响差异显著(P<0.05),性别间差异不显著(表1)。叶长在性别间、温度间差异均显著。25、20 ℃下,雄株的总叶数均略高于雌株。25 ℃时雌株的单叶面积大于雄株,在15、20 ℃时雄株单叶面积大于雌株;且雌株的单叶面积随温度升高先减小再增大,雄株无规律。叶长、叶宽、单叶面积在温度与性别互作间差异均显著。
2.2温度对茎、根系性状的影响 雌、雄株茎的分枝数、茎长度、茎直径及节间长在温度间差异显著(P<0.05),性别间除分枝数外,均差异不显著(P>0.05)。分枝数和茎直径在温度和性别间的互作效应显著(表2)。所有温度下,雌株分枝数均大于雄株。20 ℃时雌雄株分枝数均最少。随温度升高,雄株茎长度和节间长性状显著升高;雌株则没有明显的规律;雄株茎直径性状显著下降,除15 ℃外且始终小于雌株,雌株无明显的规律,仅20 ℃时茎直径达到最大值。
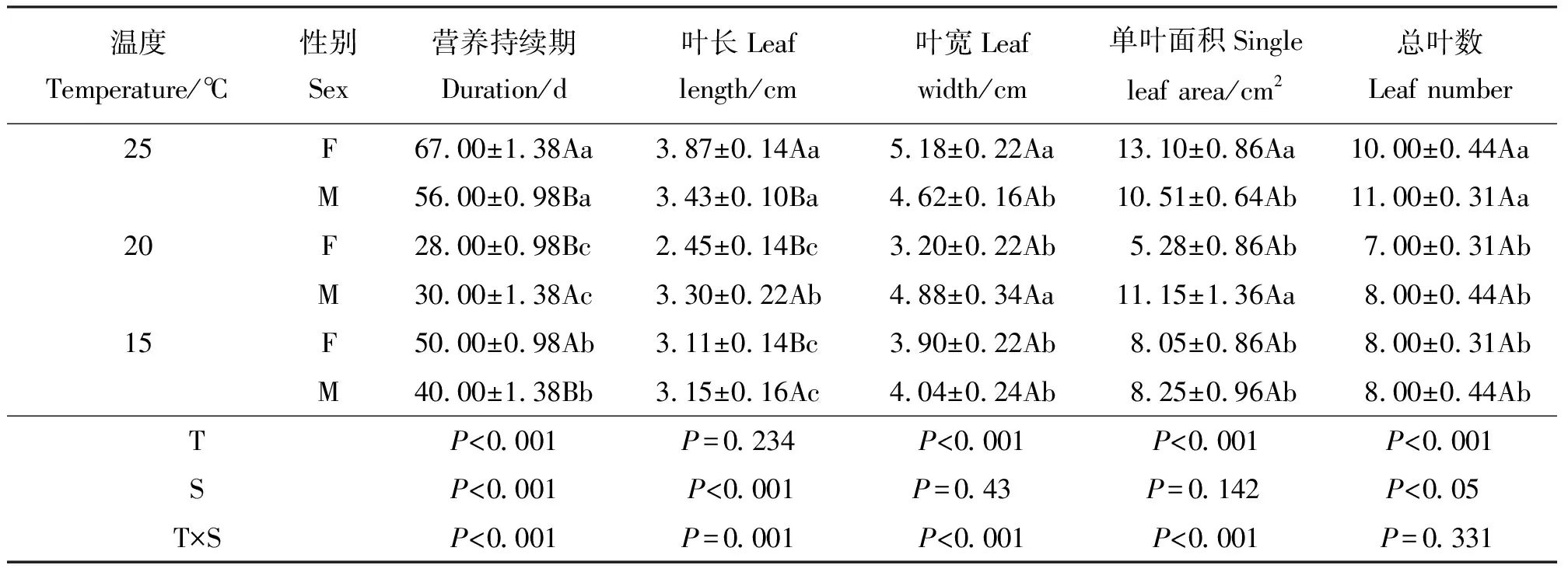
表1 营养生长持续期及叶性状对温度响应的差异Table 1 Duration of vegetative growth and leaf traits in response to temperature
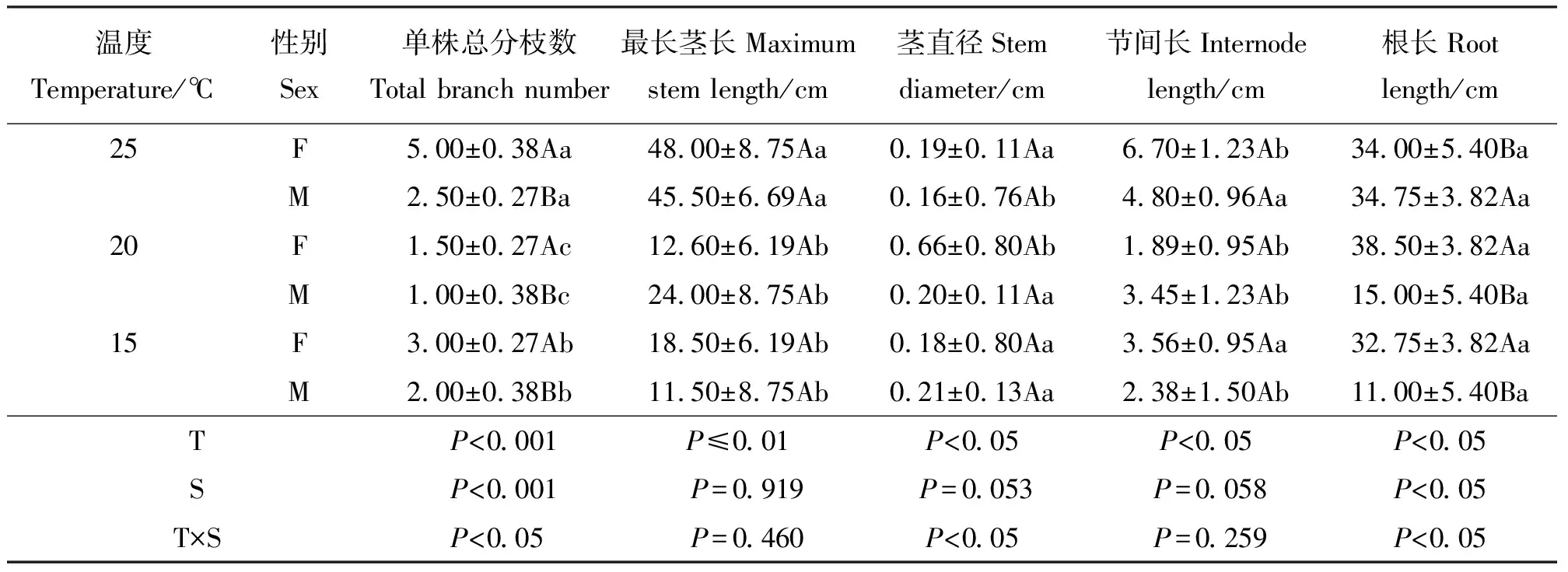
表2 茎性状及根性状对温度响应的差异Table 2 Traits of stem and root in response to temperature
雌、雄株根长在温度间、性别间及互作间均差异显著(P<0.05)。随温度升高,雌株根长先增后降,但雄株根系则越来越长。可见,雌株的根系对温度胁迫的抗性显著高于雄株,这或许就是野生种群中雌雄性比失衡的主要原因。
2.3温度对各构件鲜质量和干质量的影响 葎草雌雄植株根、茎、叶的鲜质量与干质量在温度和性别间均表现出显著差异(P<0.05)(表3)。雌株根、叶的鲜质量和干质量在不同温度水平下均显著大于雄株,茎的鲜质量和干质量仅在20 ℃时小于雄株。雌、雄株根的鲜、干质量均随着温度上升逐渐增大。雌雄株茎、叶则没有类似的规律。各构件的鲜质量的温度与性别互作效应显著。
2.4温度对单株生物量及其构成的影响 温度对葎草雌、雄单株生物总量的影响差异显著(P<0.05)。性别间生物总量差异亦显著,雌株的生物总量大于雄株(表4)。随着温度升高,根的生物量比例逐渐增大,且雌株均大于雄株,增长幅度基本一致。茎、叶的生物量比例在性别间差异显著。随着温度升高,雄株茎生物量比例逐渐增大;雌株茎生物量比例先降低再升高。随着温度升高,雌株叶的生物量比例先增后降,20 ℃时达到最大值;雄株叶生物量比例则逐渐降低,在25 ℃时与雌株基本持平。由此可知,雌雄株应对不同温度时,根茎叶生物量分配方式各不相同。随着温度上升雄株倾向于减少叶片的有机物含量,雌株则通过根的延伸以吸取更多的营养物质为接下来的生殖生长提供物质基础。根、茎、叶的生物量分配的温度与性别互作效应显著。
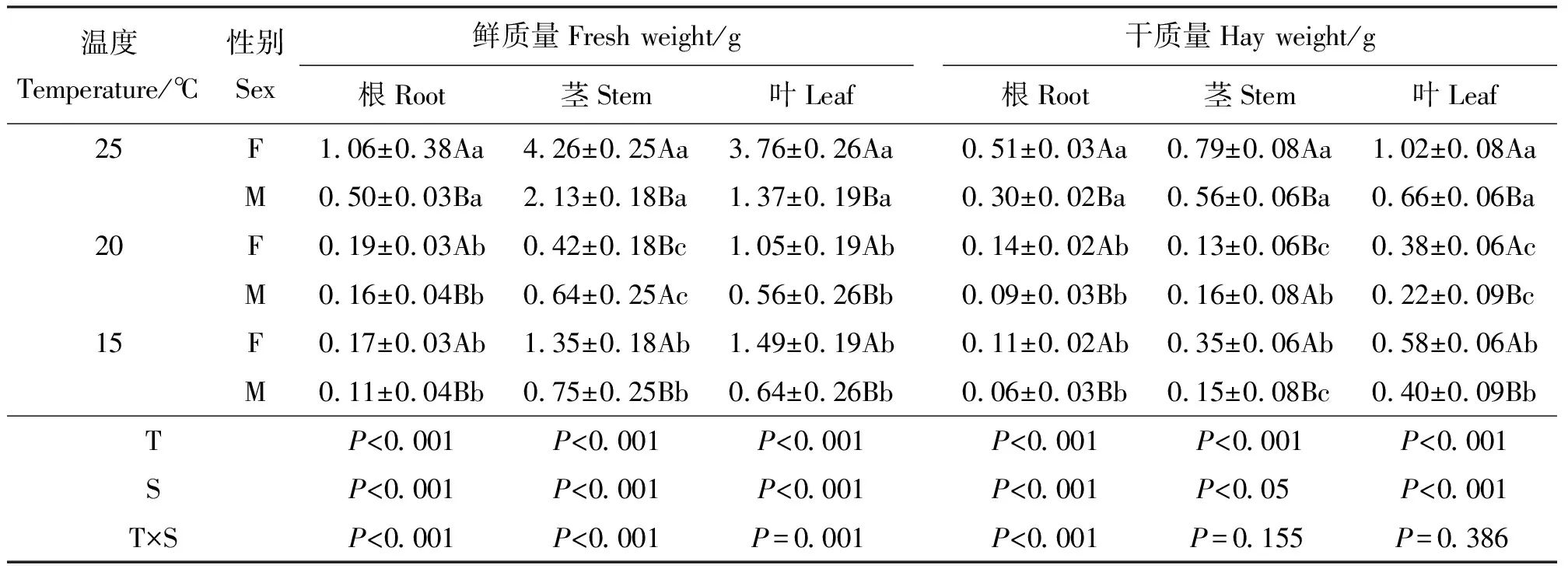
表3 构件生物量对温度响应的差异Table 3 Component biomass in response to temperature
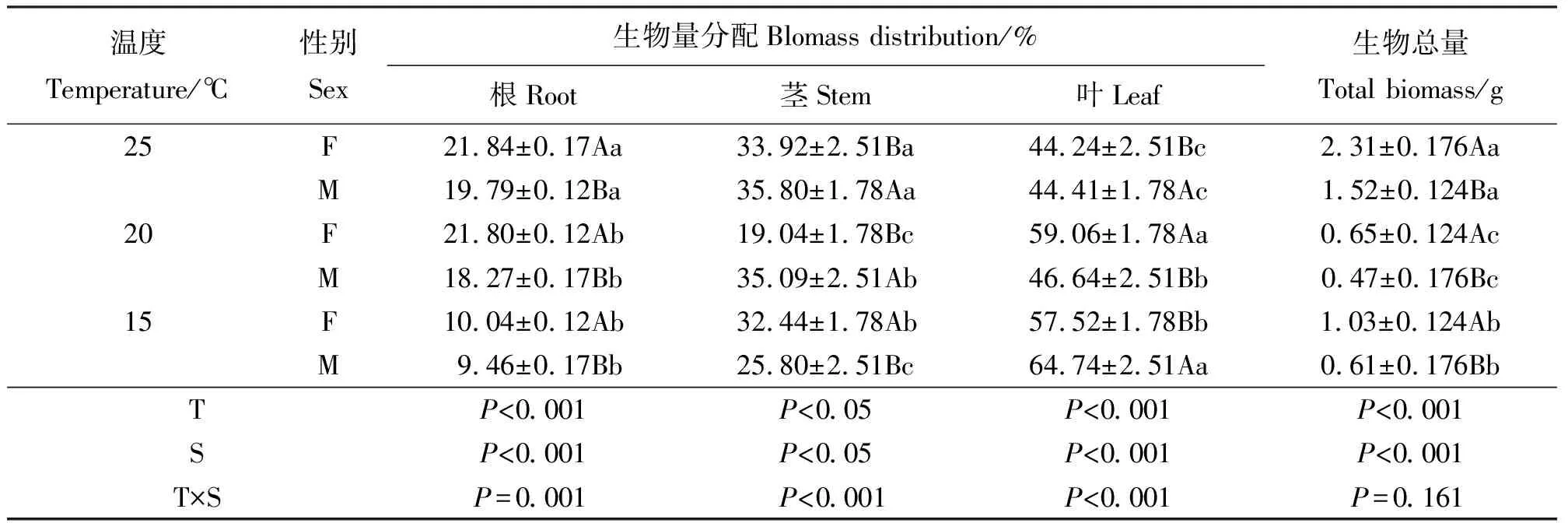
表4 生物量分配对温度响应的差异Table 4 Biomass distribution in response to temperature
3 讨论与结论
雌雄异株植物生理和生态存在着性别差异,在生长、存活、生殖格局、空间分布等方面表现出明显不同[15-17],通常认为雌株比雄株有更高的繁殖成本[18-19]。雌、雄株在营养生长期,对不同环境的应对方式直接关系到种群的形成、性比、空间格局和种群的发展趋势。据长期野外观察发现,不同生境下葎草种群的营养期差异很大,在生境良好时,第1年只进行营养生长,而不良生境条件下的种群,营养生长期很短,仅为50 d左右就转入生殖生长[13]。本研究仅通过控制温度单个生态因子,发现雌雄葎草对温度的响应存在着性别差异。温度是控制植物由营养生长向生殖生长转变的临界因子。不同植物的成花各自有其一定范围的温度需求,温度过低或过高都会抑制花的分化发育。依据本研究结果,可推断出20 ℃是最接近葎草花器官发育的温度。
植物的春季物候,如发芽、展叶、开花的时期主要取决于气温的高低[20],果实或种子的成熟等主要取决于果实生长期的积温[21]。本研究表明,温度对葎草构件数量、生物量结构及繁殖策略均有显著影响,并且雌雄个体表现出明显的性别差异,雌株对温度的响应更敏感,特别在20 ℃时,雌株先于雄株进入生殖生长阶段为花序分化及种子发育奠定物质基础。繁殖投资的不同也导致了雌、雄株的生长策略不同,雌性较于雄性由于繁殖方面的高资源投资降低了其营养生长[8]。本研究中,雌株在低温条件下,由多分枝逐渐转变为少分枝或者无分枝,茎矮壮、节间缩短,单叶面积小,根的生长加快变长,同时生物量分配上根的比例增大。但雄株的分枝数和单叶面积的变化较雌株小,尤其根系明显小于雌株。可见,雄株与雌株在营养、水分、空间等资源的竞争处于劣势,雌株的有效利用资源率明显高于雄株。营养生长是植物生殖生长的基础,温度对营养期葎草雌雄株形态的影响必然会影响生殖生长以及葎草种群性比和种群空间分布。生殖生长与营养生长密不可分,本研究仅在形态水平上研究葎草雌雄株的差别,关于花、果实以及生理生化方面还有待进一步研究。
[1] 刘生荣,张海.葎草在残塬沟壑区荒坡治理的生态效应研究[J].中国生态农业学报,2004,12(2):155-157.
[2] 孙跃春,徐彤宝,王艳玲.葎草的综合开发利用[J].中国林副特产,2003(3):57.
[3] 王桃云,马红昌,刘江颖.葎草叶蛋白提取工艺的优化[J].苏州科技学院学报(自然科学版),2005,22(1):59-64.
[4] 高政权,孟春晓.葎草研究进展[J].安徽农业科学,2007,35(31):9982-9984.
[5] 胥晓,杨帆,尹春英,等.雌雄异株植物对环境胁迫响应的性别差异研究进展[J].应用生态学报,2007,18(11):2626-2631.
[6] Li C,Ren J,Lu R.Sex-specific physiological and growth responses to water stress inHippophaerhamnoidesL.populations[J].Acta Physiol Plant,2004,26:123-129.
[7] Jones M H,Macdonald S E,Henry G H R.Sex-and habitat-specific responses of a high arcticwillow,Salixarctica,to experimental climate change[J].Oikos,1999,87:129-138.
[8] 尹春英,李春阳.雌雄异株植物与性别比例相关的性别差异研究现状与展望[J].应用与环境生物学报,2007, 13(3):419-425.
[9] 张劫,张鲲,刘湘桂,等.葎草的形态组织鉴定[J].中药材,1998,21(12):605-609.
[10] 李守阳,任相泉.葎草饲喂畜禽[J].中国畜牧兽医,2003,30(6):48-49.
[11] 雷青娟,刁治民,张正国,等.葎草的开发利用及防治对策的研究[J].安徽农学通报,2005,11(7):68-69.
[12] 刘金平,王艳.南充地区野生葎草构件性状多样性分析[J].草业科学,2009,26(8):68-71.
[13] 刘金平,康军利.野生葎草种群有性生殖投入与收益差异性分析[J].西南农业学报,2010,23(3):872-875.
[14] 刘金平,欧阳梅.成熟度与贮藏期对野生葎草种子发芽力的影响[J].安徽农业科学,2009,38(4):1817-1818.
[15] Cipollini M L,Whigham D F.Sexual dimorphism and cost of reproduction in the dioecious shrubLinderabenzoin(Laumceae) [J].American Joumal of Botany,1994,81(1):65-75.
[16] Jin J,Jiang H,Yu S Q,etal.Sex-linked photosynthesis research and the evolutionary analysis in relict plant,GinkgobilobaL[J].Acta Ecologica Snica,2008,28(3):1128-1136.
[17] Thomas S C,Lafrankie J V.Sex,size and interyearvariation in flowering among dioecioustrees of themalayan rain forest [J].Ecology,1993,74(5):1529-1537.
[18] Dawson T E,Ehleringer J R.Gender-sepecific physiology,carbon isotope discrimination, and habitat distribution in boxelder[J].Acer Negunda Ecology,1993,74:798-815.
[19] Dawson T E,Geber M E.Sexual dimorphism in physiology and morphology[A].In:Geber M A,Dawson T E,Delph L F.Gender and Sexual Dimorphism in Flowering Plant [M].New York,USA:Springer,1999:175-215.
[20] Frank M,Chmielewski,Thomas R.Response of tree phenology to climate change across Europe[J].Agricultural and Forest Meteorology,2001,108(2):101-112.
[21] 张福春.气候变化对中国木本植物物候的可能影响[J].地理学报,1995,50(5):403-408.
