谷氨酰胺转胺酶对搅拌型酸奶品质的影响
2013-02-21牟英杰林海君杨丽杰李丹凤霍贵成
牟英杰,林海君,杨丽杰,李丹凤,霍贵成
(东北农业大学,乳品科学教育部重点实验室,黑龙江哈尔滨150030)
谷氨酰胺转胺酶(TG,蛋白质-谷氨酰胺γ-谷氨酰转移酶,EC 2.3.2.13),又名转谷氨酰胺酶,是一种酰基转移酶,能催化蛋白质及多肽链中谷氨酰胺残基的γ-酰胺基(酰基供体)和伯胺(酰基受体)之间的酰胺基转移反应,在蛋白质分子内或分子间交联,形成ε-(γ-谷氨酰胺)赖氨酸异肽键[1-2],改变蛋白质的功能性质,如乳化性、流变性、溶解性等。此外,还能引入赖氨酸提高食品的营养价值。
经过热处理的乳蛋白质是TGase的良好底物。乳蛋白质经TGase作用后,蛋白质分子内或分子间交联,形成ε-(γ-glutmayl)lys二肽,虽然胃肠道中的消化液不能将其分解,但是到达肾脏后可以被γ-谷氨酰环化转移酶和γ-谷氨酰转肽酶分解为赖氨酸和谷氨酸,因此,在人体内能被消化吸收,不会降低乳蛋白的营养价值[3]。Katsuya S等[4]通过动物实验对经过TGase聚合的酪蛋白进行营养评价,发现酪蛋白聚合物中的ε-(γ-glutamyl)lys能够被吸收。
酸奶是经乳酸菌发酵后形成的酸凝乳,但是由于温度或物理条件的改变很容易出现乳清分离的问题。由于TGase作用形成共价键,提高胶体的持水力及剪切力,所以通过添加TGase可以解决这个问题,而且,它安全无毒,可以替代化学增稠剂,生产一种绿色产品。
本实验设计两种不同酶添加方式,通过对加入谷氨酰胺转胺酶生产酸奶各项指标的检测,选出能改善酸奶理化特性,并且最适宜实际生产的酶添加方法,为后期改善酸奶的生产工艺提供理论根据。
1 材料与方法
1.1 材料与仪器
谷氨酰胺转胺酶(TG-N)酶活力是100U/g 泰兴市一鸣生物制品有限公司;鲜牛奶 东北农业大学阿城牧场;蔗糖 市售食品级;氯化钠、氢氧化钠、硫酸铜、硫酸钾 分析纯,天津化学试剂三厂;氨水、盐酸、95%乙醇、乙醚、石油醚、浓硫酸、硼酸 分析纯,天津市进丰化工有限公司;甲基红、亚甲基蓝、酚酞 分析纯,天津市滨海科迪化学试剂有限公司;海砂 化学纯,上海展云化工有限公司;汉森发酵剂(嗜热链球菌、保加利亚乳杆菌);MRS培养基 青岛海博生物技术有限公司。
TA.XT Plus质构仪 英国Stable M icro System公司;MAL1038384旋转流变仪 英国MALVERN公司;SPX-150B生化培养箱 上海佳胜实验设备有限公司;Voltage 9VDC精密pH计 美国Mettler Toledo公司;AL104分析天平 美国Mettler Toledo公司;GL-20G-Ⅱ高速冷冻离心机 上海垒固仪器有限公司;HVE-50高压灭菌锅日本HIRAYAMA公司;XW-80A涡旋混匀器 上海青浦沪西仪器厂;BCD-518WS A冰箱 海尔集团。
1.2 实验方法
1.2.1 鲜牛乳的成分测定 蛋白质含量测定:参考GB 5009.5-2010《食品中蛋白质的测定》中的凯氏定氮法[5];脂肪含量测定:参考GB 5413.3-2010《婴幼儿食品和乳品中脂肪的测定》中的第一法[6];干物质含量测定:参考GB 5413.39-2010《乳和乳制品中非脂乳固体的测定》中的总固体的测定[7]。
1.2.2 酶添加方式的选择 A组为对照组:在原料乳灭菌后,冷却至43℃,加入菌种发酵。B组为酶灭活组:原料乳灭菌后冷却至50℃,添加谷氨酰胺转胺酶,添加量为0.2g/L牛乳,搅拌均匀,放入保温箱中反应20m in,再将原料乳加热到90℃灭酶10m in,立即冷却至43℃接种发酵。C组为同时加酶组:原料乳灭菌后冷却至43℃,将酶和菌种一同加入,搅拌均匀。
1.2.3 酸奶制作 酸奶的工艺流程(见图1),从鲜牛奶处理到灭菌的操作过程,在东北农业大学乳品重点实验室乳品中试车间生产线中完成。鲜牛乳过滤后,加入6%的蔗糖,经片式加热器预热至65~70℃,进入均质机,均质压力为18~20MPa,再经片式加热器90~93℃灭菌10m in。按酶灭活组和同时加酶组两种方式分别加酶,43℃接种发酵剂发酵,当对照组的pH达到4.3时终止发酵,进行搅拌,分装到150m L的无菌小烧杯中,置于4℃冰箱中贮存,每个样品3个平行。
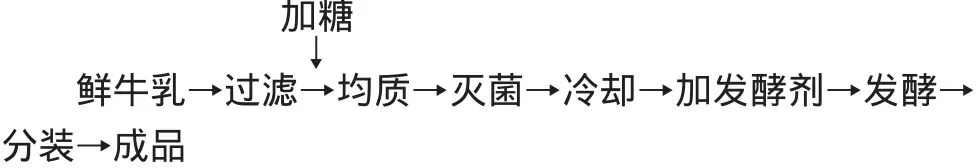
图1 酸奶工艺流程Fig.1 Processing of yogurt
1.2.4 pH和滴定酸度的测定 pH的测定:使用精密pH计直接测定;滴定酸度的测定:参考GB 5413.34-2010《乳和乳制品酸度的测定》[8]。
1.2.5 硬度的测定[9]将小烧杯置于质构仪载物台的中心位置,从4℃冰箱中取出酸奶立即测定,读取硬度的最大值。质构仪参数:探头AEC,盘径35mm,压力模式,感应力Auto-5g,测试前速度1.0mm/s,测试速度1.0mm/s,测试后速度5.0mm/s,测试距离15mm,测试时间5s。
1.2.6 流变特性的测定[10-11]用小勺轻轻搅拌酸奶20次,取约10m L置于测试台上,刮去周边多余样品,罩上铁罩防止水分蒸发并保持温度恒定。流变仪参数:夹具P/0.5/60mm,温度20℃,剪切速率在180s内从10s-1上升到50s-1,50s-1保持200s,再在180s内从50s-1下降到10s-1。
1.2.7 持水力的测定[12]称取酸奶6~10g于离心管中,在4℃下14000g离心30m in,倾去上清液,倒置10min称量残余物质量,计算酸奶保持水分能力即持水力。

1.2.8 活菌数的测定 采用MRS培养基测定酸乳中乳酸菌总菌数,计数方法采用平板菌落计数法,选取菌落数在30~300之间的平板进行计数。取0.5m L的酸奶,加入4.5m L的生理盐水中配成1∶10的均匀稀释液,用涡旋混匀器混匀,按10倍递增,制成10-5稀释液,取0.1m L 10-5稀释液于MRS固体培养基中涂布,做3个平行。置(36±1)℃温箱中培养(48±2)h,取出,计平板内菌落数目。并按下公式进行计算:

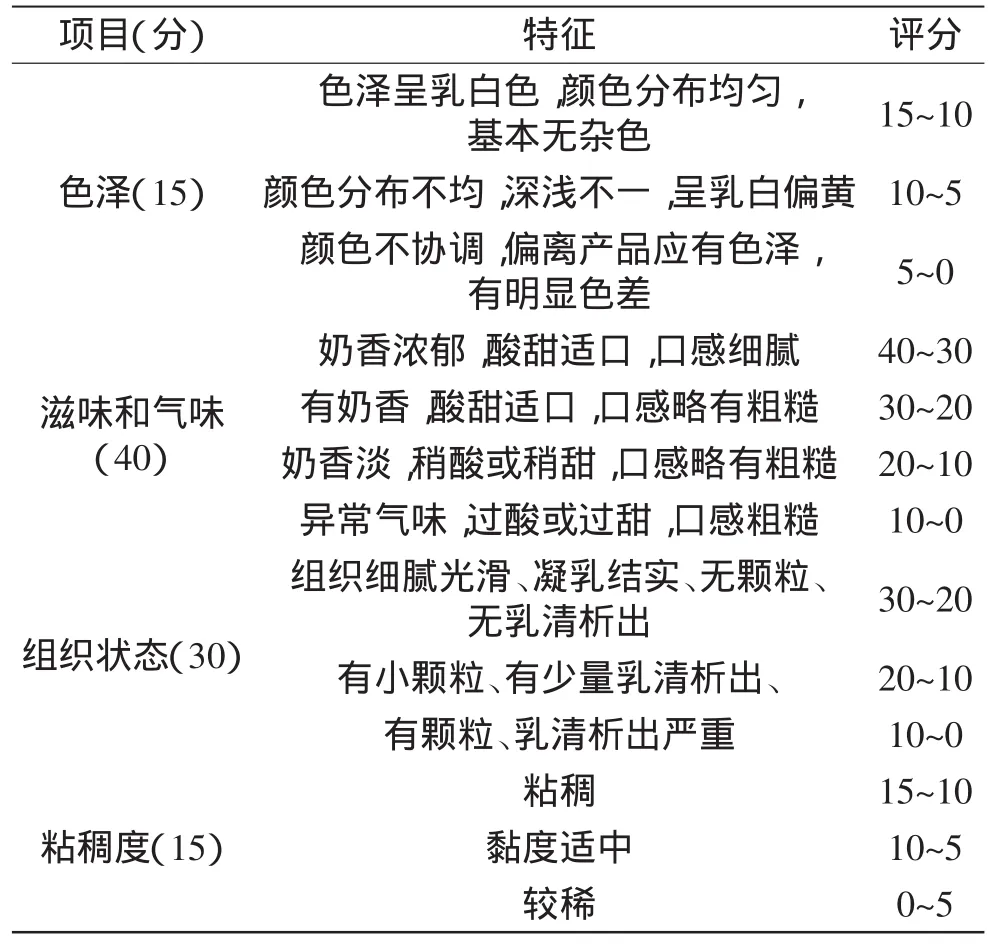
表1 酸奶感官评定标准Table1 Sensory evaluation standard of yogurts
1.2.9 感官评价 酸奶成熟后,邀请10位经过培训的食品研究人员品尝,从色泽、滋味和气味、组织状态、黏度4方面对贮存7d的酸奶样品进行感官评定,结合相关文献中酸奶的评定办法[9,13],制定评定标准,评定总分为100分,评定标准见表1。
1.3 数据处理
采用SPSS 17.0软件、Origin 6.0软件进行数据计算及分析。
2 结果与讨论
2.1 酸奶酸度和菌数的变化
2.1.1 酸度变化 从图2中可以看出,A为对照组在发酵8h(1/3d)后pH达到4.3,而B为酶灭活组pH为4.38,C为同时加酶组pH为4.33。在贮存期间A的pH最低,且滴定酸度最高,B和C的pH、滴定酸度相差不大。所有样品pH在贮存期间不断降低,滴定酸度不断升高,直到21d趋于平稳。
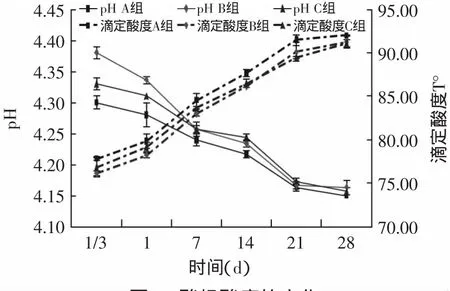
图2 酸奶酸度的变化Fig.2 Variations in the acidity of yogurts
上述结果表明加酶后的酸奶发酵时间需要延长,这可能是由于蛋白质的交联导致乳酸菌的共生关系轻微失衡[14],在贮存期间,加酶的酸奶样品后酸化较弱,可能与蛋白质交联时利用乳中的多肽或氨基酸,导致乳酸菌生长所需营养物质不足有关,菌体生长缓慢,产酸较少。根据SPSS 17.0软件进行单因素方差分析得知,样品A与B、C之间滴定酸度值差异显著(p<0.05),说明TGase对酸奶的酸度影响显著。
2.1.2 活菌数变化 由图3可知,与样品A相比,样品B和C的活菌数明显较少。所有样品的活菌数在第21d开始减少,样品A、C下降平缓,样品B下降迅速,Faergemand等[15]也曾报道在发酵前对TGase酶灭活对乳酸菌的生长有显著影响,本实验的结果与其一致。
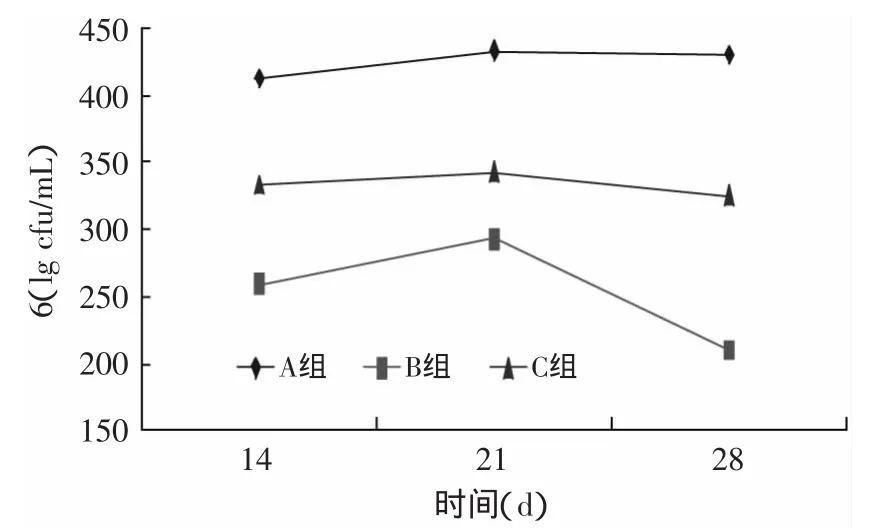
图3 贮存期间菌数的变化Fig.3 Variations in the counts of lactic acid bacteria colonies during storage of yogurts
从上述结果可以看出,TGase酶对菌种的生长有显著影响。TGase是安全无毒的,所以对乳酸菌影响的唯一可能是乳酸菌生长所需的小分子质量的多肽或氨基酸被TGase酶交联,导致部分不可被乳酸菌利用,生长受到影响[14]。
2.2 硬度和流变特性的变化
2.2.1 硬度变化 从图4可以看出,硬度总体呈先上升后下降的趋势,经TGase酶处理的样品硬度都高于对照组,与样品A相比,样品B的硬度在储存期间比较稳定,上升和下降的趋势平缓,在14d达最大值39.55g,而样品C的硬度提高幅度较大,且变化趋势明显,在21d硬度达到最大值45.05g后开始下降。样品A与样品B和C的硬度差异极显著(p<0.01)。
TGase酶对酸奶硬度的影响极显著,这是由于TGase催化交联乳蛋白,主要在分子间形成酪蛋白低聚物,增强了酸奶的凝胶强度[16]。在酶的用量相同,对酶的处理方式不同时,TGase不灭活可以较大程度的提高酸奶的硬度,而且在后期TGase酶的活力也会减弱,对酸奶的结构状态不会产生负面影响。在实际生产中,不灭活酶的方法更实用。
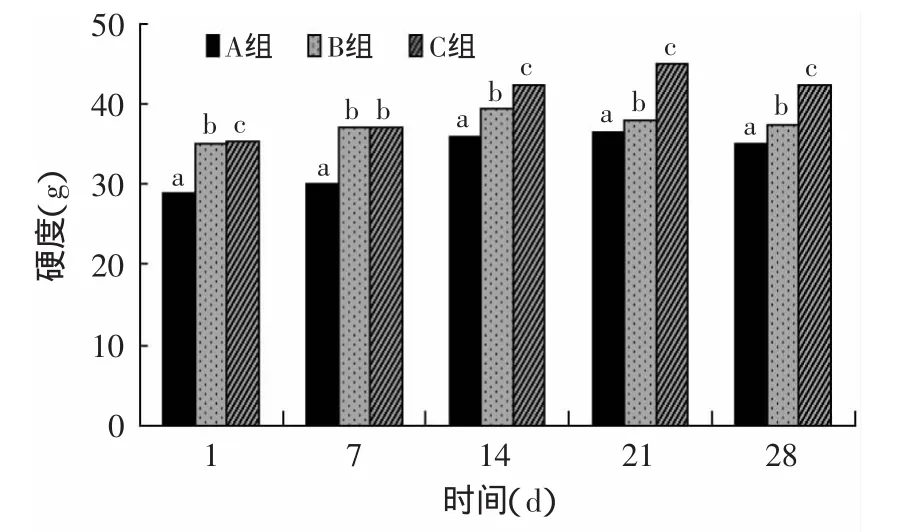
图4 贮存期间硬度变化Fig.4 Variations in the hardness during storage of yogurts
2.2.2 流变特性的变化 搅拌型酸奶是具有非牛顿剪切稀化特性的流体,随着剪切速率的增大,剪切应力呈先快后慢增加趋势,当剪切速率降低时,剪切应力呈降低趋势,但两条曲线不会重合,趋势降低的曲线应力小于趋势增加的曲线的应力,两条曲线之间的环面积(滞回曲线面积)反映破坏流体结构所需能量[9]。
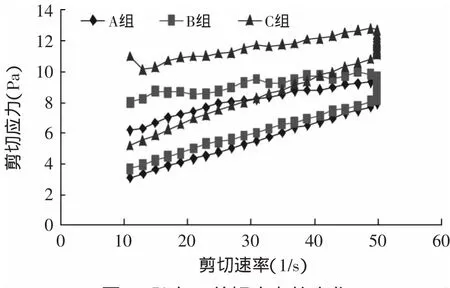
图5 贮存7d剪切应力的变化Fig.5 Variations in the shearing stress after 7d storage
从图5~图8可知,C组的剪切应力最大,在贮存期间呈上升趋势,样品A的剪切应力最小。与样品A相比,样品B的剪切应力提高不明显。搅拌型酸奶的剪切应力是蛋白质、乳酸菌体和胞外多糖相互结合的结果,蛋白质是酸奶中最主要的成分,也是应力产生的主要原因[9],加入TGase酶后,乳蛋白发生交联,形成了稳定的凝胶结构,增大了酸奶的剪切应力。经Origin 6.0软件计算,样品C的环面积最大,样品B次之,样品A最小。滞回曲线区域差异说明,经TGase酶交联的酸奶对剪切力形成了一个更稳定的凝胶结构。
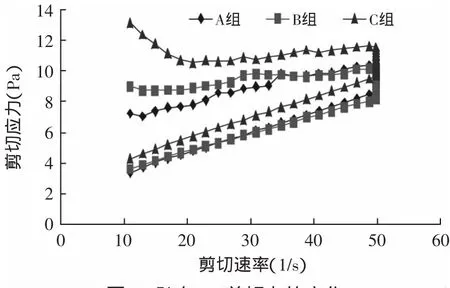
图6 贮存14d剪切力的变化Fig.6 Variations in the shearing stress after 14d storage of yogurts
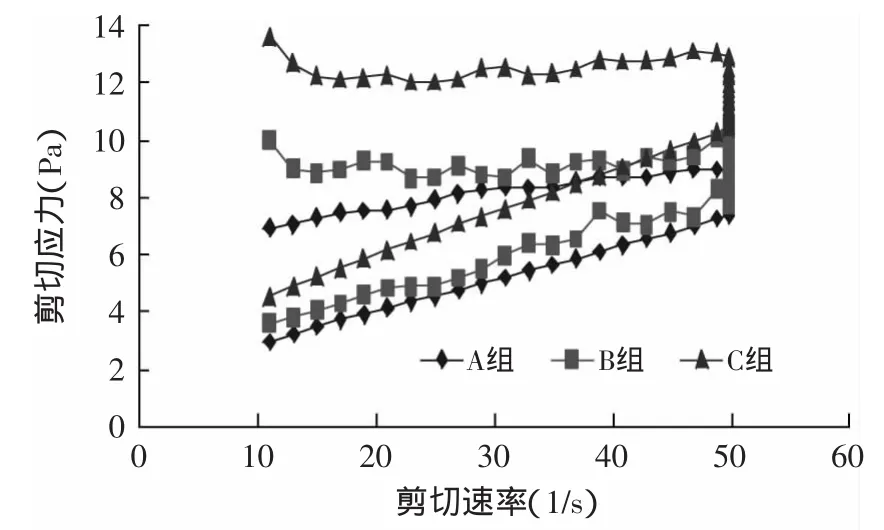
图7 贮存21d剪切力的变化Fig.7 Variations in the shearing stress after 21d storage of yogurts
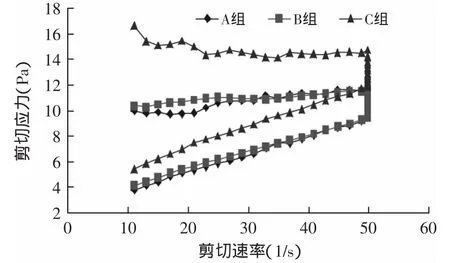
图8 贮存28d剪切力的变化Fig.8 Variations in the shearing stress after 28d storage of yogurts
2.3 持水力的变化
持水力(WHC)是表示酸奶离心后所能保留的水分,大多是与凝胶中大分子结合较牢固的水分子,凝胶立体网络中的毛细管束缚的自由水大多被除去。WHC越高说明此凝胶体系中的大分子可以通过较强的作用力结合较多的水分子,产品中结合水的含量也越高,乳清析出越少[17]。
样品A在第1d的持水力为30.01%,在第28d达到33.01%,增幅为3%,样品B和样品C第1d的持水力分别为29.87%、30.93%,增幅分别为5.13%、5.41%,样品C的增幅最大。由图9也可以看出,随贮存时间的延长,酸奶持水力不断增加,两种酶处理方法都可以改善酸奶的结构,样品C能显著提高酸奶的持水力,而且不需要对酶灭活,更符合实际生产。
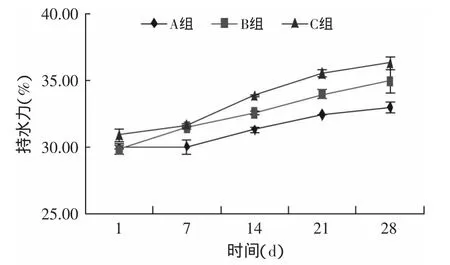
图9 贮存期间持水力的变化Fig.9 Variations in theWater holding capacity during storage of yogurts
2.4 感官评价
感官评定结果如表2所示。
酸奶样品呈乳白色,颜色分布均匀,奶香纯正,酸甜适口,无不良气味,无乳清析出。虽然三种样品在色泽、滋气味、组织状态的差别不大,总分也比较接近,但还是有一定的差异性。在色泽上,样品A、B,样品B、C两两差异不显著,样品A、C差异显著;在滋气味方面三种样品差异不显著;在组织状态上,样品A、B差异不显著,但都与样品C差异显著;在黏度上,三种样品之间的差异均显著,样品C最粘稠。经综合评价,样品C品质最优。
3 结论

表2 酸奶感官评定结果(n=10)Table2 Sensory evaluation resultof yogurts(n=10)
通过TGase交联乳蛋白来提高酸奶的理化特性是一种替代酸奶中加入稳定剂的好方法。TGase酶可以提高酸奶的硬度、持水力、剪切应力及感官特性,然而对乳酸菌的生长有不利影响,导致发酵时间的稍微延长。
通过对酸奶各项理化指标的测定和比较,发现在巴氏杀菌后将TGase酶与发酵剂同时加入,可以更显著提高酸奶的理化特性,在贮存后期TGase酶活力减弱,对酸奶的质构无不良影响,而且不需要额外的酶反应时间,热处理工序及设备,符合实际生产要求。
[1]K Yokoyama,N Nio,Y Kikuchi.Properties and applications of microbial transglutaminase[J].Appl Microbiol Biotechnol,2004,64:447-454.
[2]Hulda Chambi,Carlos Grosso.Edible films produced with gelatin and casein cross-linked with transglutaminase[J].Food Research International,2006,39:458-466.
[3]付颖.微生物谷氨酰胺转胺酶对酸奶品质的影响[D].上海:华东师范大学,2005.
[4]Katsuya S,Yoshiyuki K,Chiya K,et al.Theε-(γ-Glutamyl)lysine Moiety in Crosslinked Casein Is an Available Source of Lysine for Rats[J].The Journal of Nutrition,1996,126(10):2557-2562.
[5]GB 5009.5—2010.食品中蛋白质的测定[S].北京:中国标准出版社,2010.
[6]GB 5413.3—2010.婴幼儿食品和乳品中脂肪的测定[S].北京:中国标准出版社,2010.
[7]GB 5413.39—2010.乳和乳制品中非脂乳固体的测定[S].北京:中国标准出版社,2010.
[8]GB 5413.34-2010.乳和乳制品酸度的测定[S].北京:中国标准出版社,2010.
[9]田辉,牟英杰,杨丽杰,等.半工业化生产酸奶与直投式发酵剂的性能评定[J].食品科技,2012,37(2):52-57.
[10]Amatayakul T,Halmos AL,Sherkat F,et al.Physical characteristics of yoghurts made using exopolysaccharide producing starter cultures and varying casein to whey protein ratios[J].International Dairy Journal,2006,16:40-51.
[11]Martin PBönisch,Manfred Huss,Sabine Lauber,etal.Yoghurt gel formation bymeans of enzymatic protein cross-linkingduring microbial fermentation[J].Food Hydrocolloids,2007,21:585-595.
[12]Gosta Bylund.Dairy Processing Handbook[M].Sweden Tetra Pak Processing Systems AB,1995:250.
[13]刘靖竟,乔发东.糯米酒制作凝固型酸奶的工艺研究[J].中国酿造,2011,228(3):173-178.
[14]Barbaros Ozer,Huseyin Avni Kirmaci,Sebnem Oztekin,et al.Incorporation of microbial transglutaminase into non-fat yogurt production[J].International Dairy Journal,2007(17):199-207.
[15]Faergemand M,Sorensen M V,Jorgensen U,et al.Transglutaminase:Effect on instrumental and sensory texture of set style yoghurt[J].Milchwissenschaft,1999,54(10),563-566.
[16]CGauche,TTomazi,PLM Barreto,etal.Physical properties of yoghurtmanufactured with milk whey and transglutaminase[J].LWT-Food Science and Technology,2009(42):239-243.
[17]孙慢慢.变性淀粉的性质及在酸奶中的应用研究[D].无锡:江南大学,2012.
