蚕豆-土壤系统对14CO2的吸收和积累
2012-12-26史建君浙江理工大学理学院浙江杭州310018
史建君 (浙江理工大学理学院,浙江 杭州 310018)
蚕豆-土壤系统对14CO2的吸收和积累
史建君*(浙江理工大学理学院,浙江 杭州 310018)
为了探明14CO2在环境中的行为,采用核素示踪技术研究了蚕豆-土壤系统对14CO2的吸收和积累动态.结果表明,通过蚕豆叶片光合作用从空气中吸收的14CO2会向蚕豆其他部位组织输送并形成积累趋势,被检测到的14C比活度数值比较大,表明空气中的14CO2易于通过叶片吸收而进入蚕豆各组织器官中;蚕豆各部位组织中14C比活度随时间呈线性增长,增长速率介于20.3~45.1Bq/(g⋅d),大小次序为:叶>茎>根>豆壳>豆粒.蚕豆对14CO2(14C)具有较强的富集作用,各部位的富集系数随时间呈快速增加, 其中叶片中的富集系数最高(56d时高达31.61),豆壳次之(56d时达25.57).利用蚕豆的这一富集特性可监测大气14CO2污染的情况.
蚕豆-土壤系统;14CO2;吸收;积累
核电在正常运行条件下是一种比较安全、清洁和经济的能源,但因其“三废”中含有放射性核素,人们对核电站的安全及对环境的影响问题一直非常关注.特别是切尔诺贝利核电站事故,造成大量放射性核素进入环境,许多人和动、植物因此受到辐射损伤[1-2].核电站正常运行时主要通过气态与液态排放物对环境构成影响,气态排放物主要有131I、134Cs、137Cs、58Co、60Co、90Sr、14C、144Ce等,液态排放物主要有3H、131I、134Cs、137Cs、58Co、60Co与90Sr等.14C作为主要放射性气态流出物,是最受关注的核素之一,它半衰期为 5730a,以14CO2形态存在的14C,将参与植物的光合作用而进入食物链,危害环境和人类健康.有关核电站排放的放射性核素137Cs、90Sr、60Co、95Zr、3H等在生态系统中的行为和特性研究报道较多[3-9].但对14C的研究相对较少,作者研究了樟树对14CO2的吸收和积累,孙桂莲等[11]研究了14C在水-金鱼藻系统中的消长动态,林明明等[12]研究了小麦对14CO2的吸收和积累动态,Linda等[13]研究了14C在海洋中的迁移模型.有关蚕豆-土壤系统对14C的吸收和积累研究尚未见报道.
为了探明生态环境中14CO2的行为特性,本试验应用同位素示踪技术研究了蚕豆-土壤系统对14CO2的吸收和积累动态,为科学评定蚕豆-土壤系统对14CO2的吸收、积累状况提供基础数据,为制定适合我国国情的标准、法规提供依据.
1 材料与方法
1.1 供试核素
NaH14CO3由中国原子能研究院同位素研究所提供,为透明液体,放化纯度>95%,比活度3.7×106Bq/mL,使用前按质量比 1:1.05×105配制成0.84mol/L浓度的NaH14CO3+NaHCO3混合液,混合液的14C比活度为1.85×107Bq/L.
1.2 供试土壤
供试砂壤土,取自浙江大学华家池实验农场,其理化性质参见文献[10] .
1.3 试验方法
在3只1000mm×1000mm×300mm的水泥池中各盛入砂壤土300kg(土壤含水量约18%,每池土均匀拌人90g(NH4)2SO4和120g KH2PO4作基肥),土层厚度约24cm.于当年11月20日播种蚕豆,每池播16穴,每穴3粒种子,出苗后每穴保留2株健壮蚕豆苗.在蚕豆的生长过程中适时施肥,试验水池建于露天网室中.于次年3月20日将3只与水泥池相配套的800mm×850mm×700mm污染试验箱罩(透明有机玻璃制成,内置CO2发生器和微型风扇,顶部设有4个(100×100)mm2的采样口)罩在对应的水泥池上,箱体与水泥池的接口部位用碎土填充密封,取 2.00mL NaHCO3+ NaH14CO3)混合液与5mLHClO4溶液(0.37mol/L,适度过量)在污染试验箱体内置CO2发生器中作用产生14CO2+CO2混合气体,同时开启微型风扇使其均匀扩散,初次引入后污染试验箱体内大气CO2浓度约为0.04%,14C比活度为77.7Bq/L,每隔7d引入一次,共引入8次.
1.4 采样和制样
于首次14CO2+CO2引入后第0,7,14,21,28,42, 56d采样(采样在当天引入14CO2+CO2前进行),每池随机取蚕豆2株,3池共6株,根部用清水冲洗后用吸水纸吸去表面水,称重后将蚕豆分为根、茎、叶、豆壳和豆粒(如果有的话)5部分;用半筒式不锈钢取土器每池取土柱 1个,3池共 3个,随即将土柱按约每40mm横向均等分割成6段,所有样品(植物和土壤)分别称重后置于干燥箱中于60℃下烘干,每种样品3个重复.
将烘干后的样品磨碎混合均匀后,植物样品称取60mg,土壤样品称取100mg(每种样品3个重复)置于 OX-600型生物氧化燃烧仪上于850oC下燃烧3min,用含乙醇胺的闪烁液收集燃烧产生的14CO2,氧气通量和清洗氮气的通量约为 350mL/min,以提取植物和土壤中的14C.燃烧前接收管中先装8mL闪烁液,燃烧后将其倒入测样瓶中,并用 2mL闪烁液冲洗一次,冲洗液一并加入测样瓶中.闪烁液配方为: 0.5g POPOP+5gPPO+600mL二甲苯+150mL乙醇胺+150mL乙二醇乙醚+100mL甲醇.
1.4 14C活度测定与数据处理
所用样品均在Wallac 1400 DSA液闪仪上测定,测量误差控制在5%以内.测量结果经相应的燃烧回收效率校正和扣除本底后,换算成样品的比活度. 整株蚕豆中的浓度按其各部位中的浓度加权平均计算获得.土壤的14C比活度指全土平均.富集系数 CF[14]=生物中的14C比活度(Bq/g)/空气中14CO2比活度(Bq/L).
2 结果与讨论
2.1 蚕豆-土壤系统对14CO2吸收和积累动态
由表1可见,在蚕豆的根、茎、叶、豆壳和豆粒(如果有的话)中均检测到了14C的存在,表明通过光合作用从空气中吸收的14CO2会在蚕豆各组织中积累,被检测到的14C比活度数值比较大,表明空气中的14CO2易于通过叶片吸收而进入蚕豆各组织器官中.
蚕豆各部位组织的14C比活度均随时间呈逐渐上升的趋势,具有明显的积累特征.叶片是进行光合作用的器官,光合作用产生的同化产物首先在叶片中积累,致使叶片中的14C比活度高于其他部位;茎杆具有输送同化产物的作用,其14C比活度次之;根中的14C比活度相对较低,可能与根系生长相对上部组织比较缓慢,需要的同化产物较少有关.生长后期结出豆粒后,豆壳和豆粒中的14C比活度略低于叶片,但高于茎和根部的14C比活度,豆粒是新生组织,光合作用产生的同化产物向新生组织输送的量较多,致使豆粒中的14C比活度水平较高.
蚕豆各部位中的14C比活度均随时间而增长,对表 1数据进行线性回归分析表明: 蚕豆组织中14C比活度C的变化与时间t间呈线性正相关,拟合方程如下:
根: C=29.9t-83.6 r2=0.944;比活度随时间的增长速率为29.9Bq/(g·d)
茎: C=34.6t-77.1 r2=0.978;比活度随时间的增长速率为34.6 Bq/(g·d)
叶: C=45.1t+81.7 r2=0.979;比活度随时间的增长速率为45.1 Bq/(g·d)
豆壳: C=24.2t+783.1 r2=0.760;比活度随时间的增长速率为24.2 Bq/(g·d)
豆粒: C=20.3t+879.9 r2=0.692;比活度随时间的增长速率为20.3 Bq/(g·d)
整株: C=40.0t-48.5 r2=0.935;比活度随时间的增长速率为40.0 Bq/(g·d)
根、茎、叶和整株中14C比活度与时间的线性相关性相当好,豆壳和豆粒略差一些,其原因是豆粒在生长初期大量吸收同化产物,致使14C比活度迅速上升,在经过灌浆期后,生长逐渐趋缓,呈现出先快速上升,后逐渐走平的特性.
蚕豆各部位14C比活度随时间的增长速率介于 20.3~45.1Bq/(g·d)范围内, 大小次序为:叶>茎>根>豆壳>豆粒.其中叶片中14C比活度的增长速率最大,达45.1Bq/(g·d),是豆粒的增长速率的2.22倍.
与放射性金属离子(89Sr、95Zr和137Cs)相比较,14CO2在作物中的分布特性与137Cs相近[15-16],与89Sr、95Zr不同,89Sr主要积累在根和茎叶组织中,果实中的含量较低[17],95Zr主要蓄积在地下部和浸于水中的茎杆下部,上部组织几乎为0[18].
土壤中的14C比活度总体上处于较低水平,随时间呈现逐渐上升的趋势,14C进入土壤的途径主要是来源于空气中的14CO2及通过植物根系与土壤溶液发生交换而进入土壤.
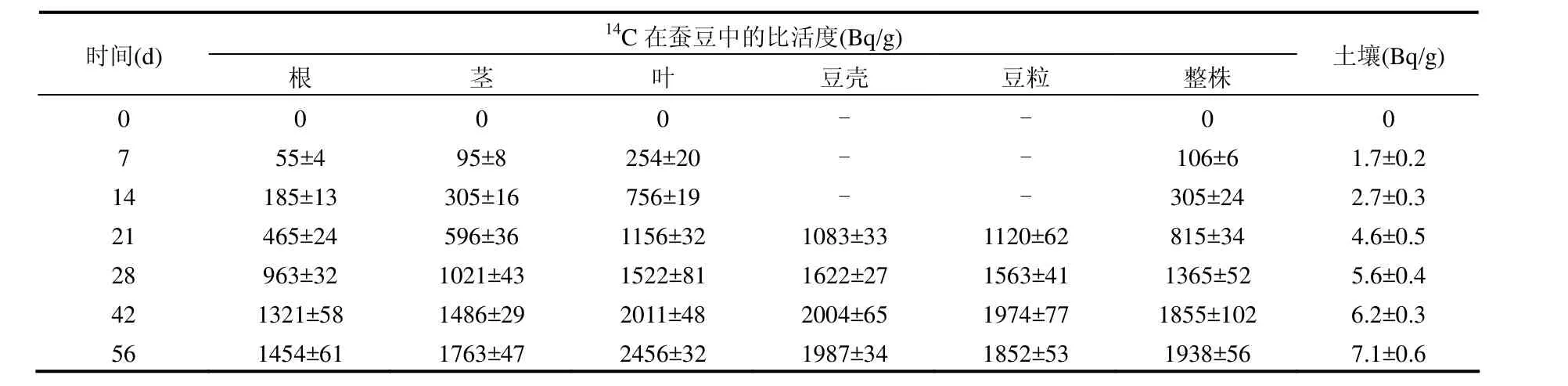
表1 14C在蚕豆-土壤系统各组分中比活度的动态变化(干重)Table 1 Dynamic change in 14C specific activity in a broadbean-soil system (dry weight)
2.2 蚕豆对14CO2的富集特性
14C的生物学性质类似于普通碳,被生物体吸收后会参于生物的各种新陈代谢过程, 蚕豆各组织对来源于空气中14CO2的14C的富集系数除 7d时的根外均大于 1(表 2),表明蚕豆对14CO2(14C)具有较强的富集作用;蚕豆各部位组织对14C的富集系数随时间呈增加趋势,具有明显的积累效应,且各部位组织的富集系数有所不同,其中叶片中的富集系数最高(56d时高达31.61),豆壳次之(56d时达 25.57),富集特性与小麦相似[12].利用蚕豆的这一富集特性可监测大气14CO2污染的情况.
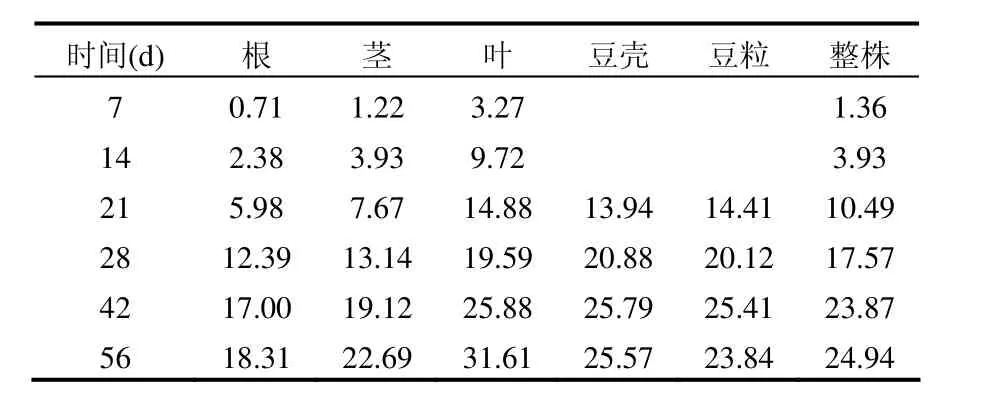
表2 蚕豆对14CO2的富集系数Table 2 Concentration factor values of 14C in broadbean
2.3 14C在土壤中的垂直分布
14C在土壤中的比活度随土壤深度及时间的变化如表3所示,总体上看处于较低水平.表层土略高一些,分析其原因,表层土中的14C主要来源于空气中的14CO2,空气中的14CO2与表土中的水分接触溶解成碳酸而进入表土,致使表土14C比活度高于中下层土壤,且随时间的增加溶解量增加,致使表土中的14C比活度呈逐渐增加的趋势.中层土壤(4~16cm)也有少量分布,其来源主要为植物根系与土壤溶液发生离子交换而致使14C进入土壤(植物根系主要分布在中层土壤).下层土壤很少,接近于本底.
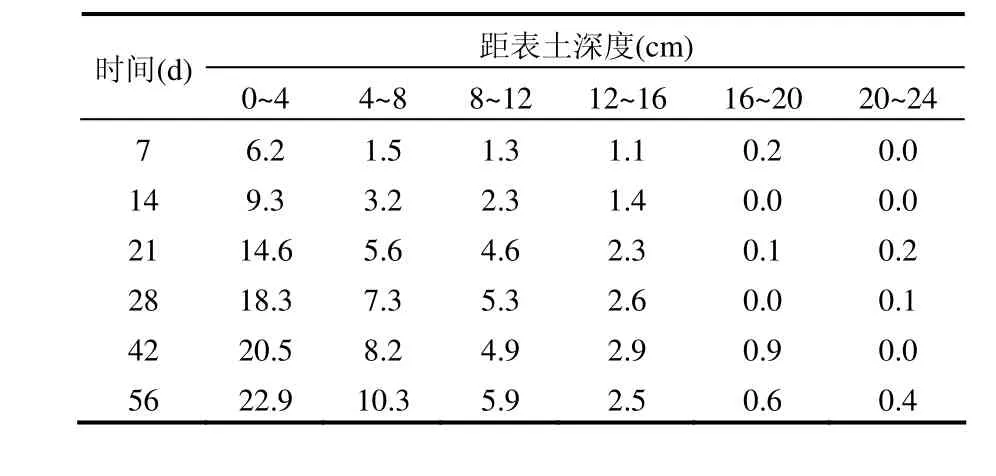
表3 14C在土壤中的垂直分布(Bq/g)Table 3 Vertical distribution of 14C specific activity in the soil (Bq/g)
2.4 14CO2引入量与14C积累量的关系
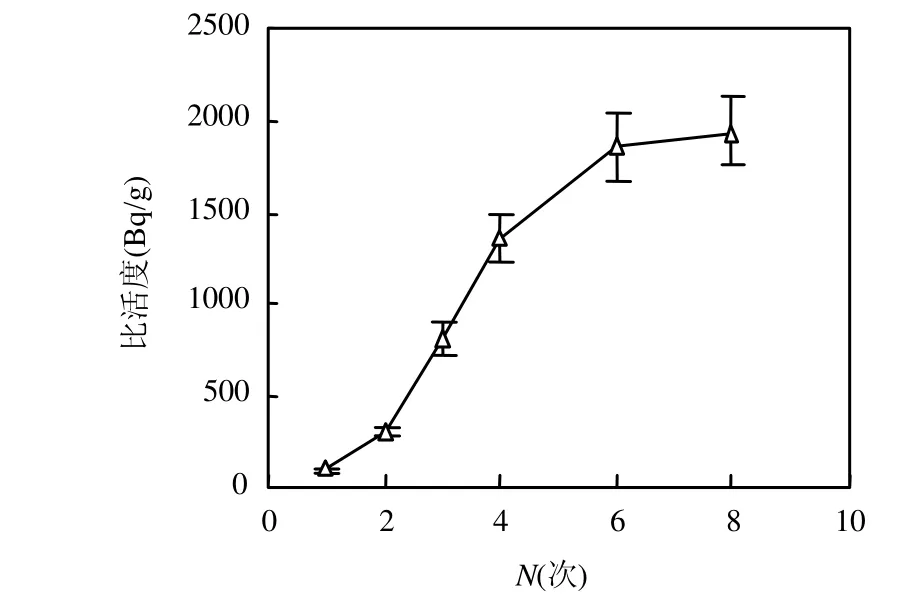
图1 蚕豆植株中14C的积累动态Fig.1 Accumulation of 14C specific activity in broadbean
由图1可见,蚕豆植株中的14C比活度随引入次数的增加而增大,至后期增长速率有所减缓,其原因是生长后期蚕豆植株已进入老熟期,光合作用和对同化产物的需求均减弱,因此表现出对14C积累效应减弱的现象.
蚕豆植株中14C比活度与14CO2引入次数的线性关系为:C=285.85N-79.41,r2=0.9151;随引入次数的增长速率为 285.85Bq/(g·次),线性关系略差一点,这主要与豆的生长周期有关.
3 结论
3.1 通过蚕豆叶片光合作用从空气中吸收的14CO2会在蚕豆各组织中积累,被检测到的14C比活度数值较大,表明空气中的14CO2易于通过叶片吸收而进入蚕豆各组织器官中.
3.2 蚕豆各部位组织的14C比活度随时间呈逐渐上升的趋势,增长速率介于 20.3~45.1Bq/(g·d)范围内,大小次序为:叶>茎>根>豆壳>豆粒.其中叶片中14C比活度的增长速率最大.蚕豆对14CO2(14C)具有较强的富集作用,富集系数也随时间呈增加趋势,其中叶片中的富集系数最高(56d时高达31.61),豆壳次之(56d时达25.57).
3.3 来源于空气中的14CO2也会轻微污染土壤,其主要途径有二:其一为空气中的14CO2与表土中的水分接触溶解成碳酸而进入表土,其二为通过植物根系与土壤溶液发生离子交换而致使14C进入土壤.
[1] Moore D H, Tucker J D, Jones I M, et al. A study of the effects of exposure on cleanup workers at the chernobyl nuclear reactor accident using multiple and points [J]. Radiation Research, 1997,148(5):463-475.
[2] Hutchins D A, Stupakoff I, Hook S, et al. Effects of Arctic temperatures on distribution and retention of the nuclear waste radionuclides241Am,57Co and137Cs in the bioindicator bivalve macoma balthica [J]. Marine Environmental Research, 1998, 45(1):17-28.
[3] Liu L L, Shi J J, Zhao X Y, et al. Dynamics of transfer and distribution of95Zr in the broadbean-soil ecosystem [J]. Journal of Environmental Radioactivity, 2005,80:217-233.
[4] Golikov V, Logacheva I, Bruk G, et al. Modelling of long-term behaviour of cesium and strontium radionuclides in the Arctic environment and human exposure [J]. Journal of Environmental Radioactivity, 2004, 74 (1-3):159-169.
[5] Chao J H, Chiu C Y, Lee H P. Distribution and uptake of137Cs in relation to alkali metals in a perhumid montane forest ecosystem [J]. Applied Radiation and Isotopes, 2008,66(10):1287-1294.
[6] Choi Y H, Lim K M, Lee W Y, et al. Tissue free water and organically bound tritium in the rice plant acutely exposed to atmospheric HTO vapor under semi-outdoor conditions [J]. Journal of Environmental Radioactivity, 2002,58(1):67-85.
[7] Wei G X, Wang Y B, Wang Y L. Using137Cs to quantify the redistribution of soil organic carbon and total N affected by intensive soil erosion in the headwaters of the Yangtze River, China [J]. Applied Radiation and Isotopes, 2008,66(12):2007-2012.
[8] Shi J J, Guo J F. Uptake from soil and distribution of95Zr in Chinese cabbage [J]. Journal of Agricultural Science, 2002,139: 431-435.
[9] Zhao X Y, Cai Z Q, Gong F H, et a1.Transference kinetics of60Co in an aquatic-terrestrial ecosystem [J]. Nuclear Science and Techniques, 2008,19(4):213-217.
[10] 史建君.樟树对14CO2的吸收和积累 [J]. 中国环境科学, 2010, 30(12):733-737.
[11] 孙桂莲,史建君.14C在水-金鱼藻系统中的消长动态 [J]. 中国环境科学, 2006,26(2):219-223.
[12] 林明明,史建君,陈 晖.小麦对14CO2的吸收和积累动态 [J].中国环境科学, 2007,27(6):841-844.
[13] Linda K, Michael G, Ulrik K, et al. An ecosystem model of the environmental transport and fate of carbon-14 in a bay of the Baltic Sea, Sweden [J]. Ecological Modelling, 2003,166(3): 193-210.
[14] 陈传群,王寿祥,徐寅良,等.放射生态学论文集 [C]. 北京:原子能出版社, 1995.201-219.
[15] 徐寅良,陈传群, 陈 斌,等.水稻对134Cs的吸收和134Cs在水稻-土壤中的分配 [J]. 核农学报, 1991,5(1):19-24.
[16] 商照荣,于凤义,路子显.秦山、大亚湾核电站周围地区土壤盆栽水稻对137Cs的吸收规律 [J]. 辐射防护, 1995,19(3):183-188.
[17] 史建君,王寿祥.水稻对放射性锶的吸收及撒洒硅藻土对其行为的影响 [J]. 核农学报, 2003,17(3):203-206.
[18] Shi J J, Guo J F, Chen H. Dynamics of95Zr in the Rice/Water/Soil System [J]. Applied Radiation and Isotopes, 2002,56:735-740.
Uptake14CO2from air and accumulation of14C in broadbean-soil system.
SHI Jian-jun*(Institute of Science, Zhejiang Sci-Tech University, Hangzhou 310018, China). China Environmental Science, 2012,32(1):120~124
Uptake14CO2from air and accumulation of14C in broadbean-soil system were studied by using the isotope-tracer techniques, which was used to get a better understanding of the environmental behavior of14CO2. The14CO2, which was absorbed from the air by the leaf photosynthesis of broadbean, would be sent to the other parts of the broadbean and form a accumulating trend.14C specific activity was detected high, which demonstrated14CO2in the air easily got into every tissue of broadbean through photosynthesis. The specific activity of14C in every tissue increased linearly with time. And the increasing rate was within the range from 20.3 to 45.1 Bq/(g·d). The order of the rate in tissues was as below: leaf blade >stalk> root >husk>seed. The broadbean had an intensive concentration of14CO2. The concentration factor values increased rapidly with time, and those in the leaf blade reach the highest level(31.61 in 56 days) and followed by the husk (25.54 in 56days). Owing to its high accumulation of14CO2, the broadbean could be used as the indicator plant in the supervisor of14CO2air pollution.
broadbean-soil system;14CO2;uptake;accumulation
2011-03-30
国家自然科学基金资助项目(20477040);浙江省自然科学基金资助项目(Y505076)
* 责任作者, 教授, jjshi61@sohu.com
X591
A
1000-6923(2012)01-0120-05
史建君(1961-)男,浙江杭州人,教授,主要从事环境科学和放射生态学研究.发表论文80余篇.
