黑腹胃蝇三龄幼虫形态学和成虫行为方式研究
2012-12-24刘善辉胡德夫初红军曹玉成
刘善辉,胡德夫,初红军,曹 杰,曹玉成,李 凯
(1.北京林业大学生物科学与技术学院,北京100083;2.新疆阿勒泰市解放南路地区林业局,新疆 阿勒泰836599;3.新疆野马繁殖研究中心,新疆 乌鲁木齐830000;4.内蒙古赤峰市克什克腾旗桦木沟林场,新疆 赤峰025366)
马胃蝇属于节肢动物门(Arthropod)双翅目(Diptera)环裂亚目(Cyclorrhapha)胃蝇科(Gastrophilidae)胃蝇属(Gasterophilus)的专性寄生蝇类。其幼虫在马科动物消化道不同部位寄生,包括口腔、齿龈、舌、咽喉粘膜、胃、十二指肠和直肠,但主要寄生在胃内,寄生时间大约持续10个月[1-2],易造成胃溃烂、肠阻塞、贫血、腹泻和消化呼吸方面问题,严重的可引起死亡[3]。全世界已知的马胃蝇共有9种[1],除了南方胃蝇 (G.meridionalis)、扁腹胃蝇(G.lativentris)、三列棘胃蝇(G.ternicinctus),其他6种[黑腹胃蝇(G.pecorum)、肠胃蝇(G.intestinalis)、裸节胃蝇(G.inermis)、黑角胃蝇(G.nigricornis)、红尾胃蝇(G.haemorrhoidalis)和鼻胃蝇(G.nasalis)]在我国均有分布[4-5]。因此,对于其幼虫尤其是3龄幼虫的研究显得尤为重要,但国内外这方面研究较少,仅国外研究者[6]对鼻胃蝇的形态特征进行过研究。同时,由于成蝇生活期很短和野外观察条件限制,在自然环境中对于成虫行为报道的也较少,只有对肠胃蝇(G.intestinalis)[7-10]在这方面作了研究。
黑腹胃蝇作为新疆卡拉麦里地区感染马科动物的优势种群,其生活史的研究对于该地区疫源疫病检测和防控起着至关重要的作用。为此,本研究拟从马属动物自然或药物驱虫排出的粪便中获得黑腹胃蝇3龄幼虫对其3龄幼虫体长、体宽、体质量、化蛹期、羽化期、成蝇的存活期等进行观测,并完善成蝇野外生活习性特征的信息,以填补这一领域的空白。
1 材料与方法
1.1 材料来源 以中国新疆卡拉麦里自然保护区内放归野马及其同域生活的家马和蒙古野驴作为研究对象。以这些马科动物体内寄生虫为研究材料,材料主要从口服伊维菌素后的马科动物粪便中获取。伊维菌素由北京万丰药业有限公司生产,兽药GMP证书号为(2006)兽药GMP证字278号;生产许可证编号为(2006)兽药生产证字01021。即在一个旋转系统中,将伊维菌素以0.1g·kg-1剂量混入玉米(Zeamays)饲料中拌匀后对试验马匹进行投喂,然后将5匹野马保持稳定(被围栏圈养)5d,每天收集3次粪便,早、中、晚各一次,每次收集均使用塑料袋按标号收集,在2009年2月和2010年2月各进行一次。其余幼虫均来自马科动物自然排出的粪便和屠宰马匹尸检消化道中。
1.2 试验方法
1.2.1 3龄幼虫形态学特征及虫种鉴定 黑腹胃蝇3龄幼虫典型的特征就是伪头小齿的排布,由3组小齿组成,2组位于后面,1组位于口钩中间(图1a);体节成排的棘刺在背部第7和第8中间中断,第9体节仅仅有少量棘刺存在,第3体节棘刺单排,中间不断开。采集到的幼虫经磷酸缓冲液或者0.1%生理盐水清洗后,借助于胃蝇属3龄幼虫检索表和文献描述[1],利用最大放大倍数10~40倍的解剖镜(莱卡MZ75)对其幼虫棘刺、口钩等特征进行鉴定,鉴定完毕后使用100%乙醇保存。
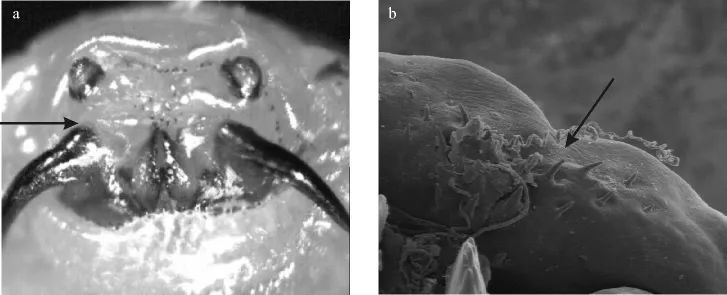
图1 黑腹胃蝇伪头中间小齿状结构Fig.1 Central denticles on the pseudocephalon of Gasterophilus pecorum
1.2.2 试验室孵化 将从粪便中获得的黑腹胃蝇3龄幼虫体长、体宽通过精度为0.02mm的游标卡尺进行测量;体质量使用精度为0.001g的电子天秤进行称量。然后将每个个体放入透明的圆柱形塑料杯子中,杯底覆盖弄湿的沙土和粪便以及滤网,然后使用带孔纱布密封直到成蝇的出现。期间按照早、中、晚各2h对每一个杯子进行观察。对幼虫化蛹、羽化时间及成蝇性别和行为等信息都进行详细记录。成蝇一旦羽化,立即转移入长0.2m、宽0.2 m、高0.5m,事先已放置少量马毛和粪便的透明塑料圆筒内,随后使用DV机录像观察。
1.2.3 野外观察 黑腹胃蝇成蝇主要特征是缺少横翅脉边缘,略带褐色的翅膀覆盖整个雌性隔膜。雄性的胸部是黑棕色,头部和腹部是黄棕色,整个身体被黄色毛发覆盖,但中间或多或少的有少量黑毛发缝合,并且黑色或者棕色的毛发也出现在小盾板上;雌性较雄性体色更黑,特别是腹部是黑棕色,有少量稀疏的黄色毛发着生。雄性和雌性腿都是黄色,体长11~16mm(图2)。

图2 黑腹胃蝇[1]Fig.2 Gasterophilus pecorums [1]
每天在野马生活的场所采用网捕法将视野可见的蝇类进行捕捉,然后根据资料记载进行鉴定[1],以确定周围是否存在马胃蝇。对马生活环境中的杂草进行检查,以确定是否有卵粘附,一旦发现卵立即剥落下来,这有利于第2天出现的新卵能够被及时发现,以防止与前1d产的卵混淆。马胃蝇卵的外形特征如图3所示。
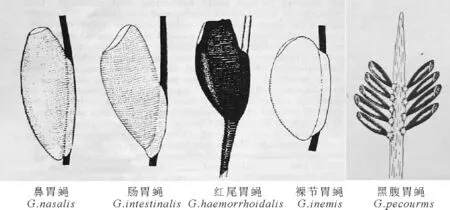
图3 马胃蝇卵形态[1]Fig.3 The eggs of Gasterophilus[1]
2 结果与讨论
2.1 3龄幼虫形态学特征 本研究共收集到1 208只马胃蝇幼虫,其中黑腹胃蝇3龄幼虫920只,2龄幼虫163只;红尾胃蝇3龄幼虫58只,2龄幼虫67只。经测定,黑腹胃蝇3龄幼虫体质量为100~420mg,98.9%个体的体质量分布在120~360mg,其中47.4%的个体体质量在200~260 mg,平均体质量为239mg。Klem等[11]从尸检所得的3龄幼虫中(包括蛹前阶段),鼻胃蝇体质量范围为30~310mg,这一数据较本研究所得到黑腹胃蝇体质量略低。本研究中黑腹胃蝇所有3龄幼虫最终成为成蝇的个体体质量为110~130mg。然而,智利研究者从自然排出的幼虫当中测得成为成蝇的个体体质量范围为130~165mg,平均体质量为145 mg[10],明显高于本研究所得数据。黑腹胃蝇成熟3龄幼虫体长范围为9~21mm,平均为13.2mm;体宽范围为2~10mm,平均为5.1mm。这与Zumpt[1]描述的黑腹胃蝇的体长为20mm的结果很吻合。
黑腹胃蝇3龄幼虫共化蛹35个,化蛹期范围为13~27d,平均为19d;在第18-20天是一个高峰期,共15个化蛹,占总化蛹数的42.86%;巅峰时期在第19天,占总共化蛹数的17.1%。阿根廷研究者观测到的鼻胃蝇化蛹期在30~40d[12],然而鼻胃蝇化蛹期较黑腹胃蝇偏长;美国研究者在室温条件下研究发现,鼻胃蝇最短化蛹期是20d[7];另一位美国研究者则观察到鼻胃蝇化蛹期为14~28d[13];南里奥格兰德州研究者观察到鼻胃蝇化蛹期为23~28d[14]。
本研究共观察到有17个(48.57%)黑腹胃蝇个体羽化出现,羽化期为15~27d,最多(29.4%)的发生在第21天。这一结果与美国研究者对鼻胃蝇观察相似,其羽化共得到56个鼻胃蝇成蝇,羽化期为15~25d,较高频率(8/56)发生在第20天,雌雄比例为3∶1[15]。
成蝇存活期为2~8d,最高的(58.82%)在第4-6天。在这些羽化的成蝇当中有88.24%是雌性,11.76%是雄性,雌雄性别比例为7.5∶1。考虑雌性和雄性的差异,雄蝇的生命跨度为1~7d,在第5天有很高的存活率;雌蝇的存活期为1~9d,在第6天存活率较高。
2.2 成虫行为方式研究
2.2.1 寻找寄主和交配行为 成蝇出现的时期仅在一年当中的很少几个月中,它们之间的相遇是偶然的,很大程度上是依靠机会,为了增加见到异性的机会,它们趋向于暂时集中于某一空间[1]。本研究发现,在成蝇出现之后,以一种狂热的方式寻找同伴,当一个成蝇和另一个成蝇出现在非常接近的距离的时候,它们能够很容易地发现彼此,从而以摔跤式方式进行碰撞。交配不仅仅只发生在马的周围,在马存在的牧场周围环境中也可能存在[16-17]。原因是成蝇为了减少自身能量供应,交配行为可以在粪堆上迅速发生,否则其必须飞到另外一个地点进行交配[18]。在本研究的几个场合(马匹周围、马粪便周围、水源地旁边、杂草堆、标本盒),仅在马匹周围捕获了少量黑角胃蝇成蝇,但并没有发现其他种马胃蝇成蝇及其交配和产卵行为,其主要原因可能与当地物种多样性和复杂的生态系统及气候变暖有关[19-20]。成蝇在交配以后开始寻找寄主,没有交配的雌蝇没有产卵的迹象,一旦交配就立即开始准备在马身上产卵[7]。当食物、交配等一些生理上的要求因素诱导时成蝇行为会变得活跃,可能表现得很有目的性[8]。
2.2.2 卵的调查 在放置成蝇的标本盒中,本研究观察到了黑腹胃蝇雌蝇有疑似交配的行为,并于次日产下黑色的卵。卵端部有类似植物根茎的穗状结构,通过这一结构牢牢地固定在附着物上(图4a)。同时在同域生活的家马体表发现了鼻胃蝇和肠胃蝇的卵,鼻胃蝇的卵呈椭圆形,腹面存在一种长条状结构,几乎整个卵的腹面都依靠此结构粘附在马毛上(图4b);肠胃蝇的卵呈楔形,只有一半腹面粘附在马毛上(图4c)。

图4 黑腹胃蝇的卵(a)、鼻胃蝇的卵(b)和肠胃蝇的卵(c)Fig.4 The eggs of Gasterophilus pecorum (a),G .nasalis (b)and G .intestinalis (c)
胃蝇属昆虫对于马的颜色没有明显的偏爱和吸引[21]。然而,本研究发现深颜色的马较浅颜色的更容易受到胃蝇的偏爱,因为观测到的肠胃蝇和鼻胃蝇的卵均是在颜色较深的马匹上发现的。这一观点也得到了研究者的证实[22]。
雌性肠胃蝇在马体表产卵部位多于胃蝇属其他种类,计算产卵数量已经被用来确定哪些部位是马胃蝇最易产卵区域的方法之一[8,22-23]。在早期的工作中,前腿内侧一直是马胃蝇产卵的偏爱场所[24-25]。本研究过程中也证实了这一观点。
肠胃蝇间歇性的从一匹马飞向另一匹马的行为被认为需要分散产卵,其产卵量高于同属的其他物种,并且是独一无二的产卵在植物上的物种[4],这与其他研究者[1,26]观点不同,他们认为产卵在植物上的物种是黑腹胃蝇,肠胃蝇产卵地点是在马体表面。俄国研究者[27]报道了黑腹胃蝇仅仅需要1 300~2 400个卵就能够感染整个牧场,从而进一步证明了黑腹胃蝇产卵地点是在植物上,而不是像肠胃蝇、鼻胃蝇等产在宿主体表。本研究在马科动物活动区域前后选取30匹家马和200个样方(10m×10m)数量的植被作为研究对象,对其是否着生黑腹胃蝇卵进行了详细检查,结果均未发现黑腹胃蝇卵的存在。究其原因可能与当地地理环境和寄主不同有关。马胃蝇这种分散产卵行为,实际是一种协同进化的适应性行为,可以为幼虫取得一个合适的寄主提供更好的机会,减少子孙后代因为失去一个寄主就面临最终死亡的几率。
2.2.3 梳理行为 在被粪便和草环绕而且有部分沙土的塑料透明杯子中,48.57%的蛹孵化成功。黑腹胃蝇破蛹而出后,需要5~10min充分舒展前翅,然后进行振翅发出类似飞机起飞前发动机的声响。肠胃蝇也会出现类似的情况[17]。梳理行为是昆虫维持体表洁净与外界进行信息交流,并完成运动、取食、呼吸等生理机能的必要前提。普遍存在于昆虫、鸟类和哺乳类等各种动物[28],具有重要的生理意义以及社会功能[29]。本研究中也观察到黑腹胃蝇具有明显的梳理行为,表现在使用前足和后足作为梳理的执行器官,梳理翅膀、头部、口器、腹部,原因是在这些部位上分布着重要的感觉器官。
致谢:感谢新疆阿勒泰恰库尔图保护站在采样过程中给予的大力支持与配合。
[1]Zumpt F.Myiasis in Man and Animals in the Old World[M].London:Butterworths,1965:267.
[2]Otranto D,Milillo P,Capelli G,etal.Species composition ofGasterophilusspp.(Diptera,Oestridae)causing equine gastric myiasis in southern Italy:Parasite biodiversity and risks for extinction[J].Veterinary Parasitology,2005,133:111-118.
[3]Sequeira J L,Tostes R A,Sequeira T C.Prevalence and macro and microscopic lesions produced byGasterophilus nasalis(Diptera:Oestridae)in the Botucatu Region[J].Veterinary Parasitology,2001,102:261-266.
[4]范滋德.中国常见蝇类检索表[M].第2版.北京:科学出版社,1992:890-895.
[5]薛万琦,赵建铭.中国蝇类(下册)[M].沈阳:辽宁科学技术出版社,1996:2207-2217.
[6]Cogley T P,Cogley M C.Field observations of the hostparasite relationship associated with the common horse bot fly,Gasterophilusintestinalis[J].Veterinary Parasitology,2000,88:93-105.
[7]Hadwen S,Cameron A E.A Contribution to the Knowledge of the Bot-flies,Gastrophilusintestinalis,Deg.,G.haemorrhoidalis,L.andG.nasalis,L.[M].H.M.Stationery Office,1918:91-106.
[8]Wells R W,Knipling E F.A report of some recent studies on species ofGasterophilusoccuring in horses in the United States[J].Iowa State College Journal Science,1937,12:181-210.
[9]Cope S E,Catts E P.Parahost behavior of adultGasterophilusintestinalis(Diptera:Gasterophilidae)in Delaware[J].Journal of Medical Entomology,1991,28:67-73.
[10]Hall M J R.Trapping the flies that cause myiasis:Their responses to host stimuli[J].Annals of Tropical Medicine and Parasitology,1995,89:333-357.
[11]Klem M A P,Rodrigues A C,Rezender A M L.Gasterofilose em equídeos:infecção natural porGasterophilusnasalisL.(Diptera:Gasterophilidae)no Estado do Rio de Janeiro [J].Revista Brasileira de Parasitologia Veterinária,1997,6(1):61-67.
[12]Alcaíno H,Gormen T,Cornejo S,etal.Estudio epizootiológico del parasitis moestomacal del equino de la Zona Centro Sur de Chile[J].Archivio Medicina Veterinaria,1980,12(1):10-29.
[13]Lahille F.Los Gastrofilos en la Republica Argentina Boletim do Ministerio da Agricultura da[J].Republica da Argentina,1911,13(12):836-856.
[14]Panitz E.Occurrence of second and third instar ofGasterophilusintestinalisandGasterophilusnasalisin horses in the Mid Atlantic United States[J].Veterinary Parasitology,1978,4:161-166.
[15]Brum J G.Periodo de pupa deG.nasalis(Diptera,Gasterophilidae)[M].Gramado,RS,Anais:Congresso Estadual Medicina veterinaria,1992:60.
[16]Rocha U F.Observagoes sobre gasterofilose emequideos da regiao de Aragatuba,Estado de Sao Paulo,Brash[J].Revista de Medicina Veterinaria de Saoaulo,1954,5(1):17-25.
[17]Catts E P.Sex and the bachelor bot(Diptera:Oestridae)[J].American Entomologist,1994,40(3):153-160.
[18]Humphreys W F,Reynolds S E.Sound production and endothermy in the horse bot-flyGasterophilusintestinalis[J].Physiological Entomology,1980,5:235-242.
[19]李晓东,傅华,李凤霞,等.气候变化对西北地区生态环境影响的若干进展[J].草业科学,2011,28(2):286-295.
[20]岳静,郝敏,张灵菲,等.物种多样性-生态系统功能关系的影响因素[J].草业科学,2011,28(4):532-535.
[21]Sutcliffe J F.Distance orientation of biting flies to their hosts[J].Insect Science and its Application,1987(8):611-616.
[22]Pandey V S,Ouhelli H,Elkhalfane A.Observations on the epizootiology ofGasterophilusintestinalisandG.nasalisin horses in Morocco[J].Veterinary Parasitology ,1980,7:347-356.
[23]Brocard P,Pfister K.Approche de l'Épidémiologie de la Gastérophilous du cheval en Suisse[J].Schweiz Arch Tierheilk,1991,133:409-416.
[24]Egri B.Megfigyelesek aGasterophilusintestinalis(De Geer,1776)peterakasanak dinamikajarol eslokalizaciojarol Magyarorszagon[J].Agrártudományi Egyetem Közleményei,Hungary,1985,27:83-94.
[25]Clark,Bracy.An Essay on the Bots of Horses and Other Animals[M].London:Old Bailey,1815:72.
[26]Suter I,Brown G,Hansman D.Horse bot-fly(Gasterophilusintestinalis)eggs laid on human hair[J].Medical Journal of Australia,1972,2:609-611.
[27]Chereshnev N A.Biological peculiarities of the botflyGasterophiluspecorumFabr.(Diptera:Gastrophilidae)[J].Doklady Akademii Nauk SSSR,1951,77:765-768.
[28]Spruijt B M,Van Hooff J A,Gispen W H.Ethology and neurobiology of grooming behavior[J].Physiological Reviews,1992,72(3):825-852.
[29]Phillis R W,Bramlage A T,Wotus C,etal.Isolation of mutations affecting neural circuitry required for grooming behavior in Drosophila melanogaster[J].Genetics,1992,133(3):581-592.
