MicroRNA-450a-3p 通过抑制Bub1 基因的表达调控小鼠细胞增殖和胚胎发育*
2012-12-23杨丹丹蒋凤兵白慧丽李宝林湛晓琴
刘 晨, 杨丹丹, 蒋凤兵, 白慧丽, 李宝林, 罗 敏, 湛晓琴, 施 琼
(重庆医科大学检验医学院临床检验诊断学教育部重点实验室,重庆400016)
自然流产的发生率约占临床妊娠的10% ~20%,其中因染色体数目异常引起的自然流产为总数的50%左右[1],而染色体数目异常胚胎细胞的发生机制至今尚不清楚。胚胎细胞的正常分裂是胚胎发育过程中的关键环节。为了确保分裂的正确性,真核生物细胞内形成一套监护网络即纺锤体组装检查点(spindle assembly checkpoint,SAC)。染色体着丝粒行使检查点功能时,关卡蛋白(checkpoint,CP)起着重要作用[2],其中Bub1 是最主要的组件蛋白之一,且Bub1 基因表达下调可导致细胞分裂染色体异常,进而影响细胞增殖和胚胎发育。最近发现,microRNA(miRNA)在胚胎干细胞的发育和分化过程中发挥重要作用,Perera 等[3]认为miRNA 可能通过调控Bub1 的mRNA 和/或蛋白水平影响胚胎发育。经过生物信息学方法预测,miR-450a-3p 可以调控靶基因Bub1 的表达。本文旨在以小鼠胚胎成纤维细胞(mouse embryonic fibroblasts,MEFs)为研究对象,通过应用miR -450a -3p 模拟体(mimic)确定miR-450a -3p 能特异性结合Bub1 的3’- 非翻译区(untranslated region,UTR),利用Western blotting、实时定量PCR (quantitative real - time PCR,qRT -PCR)等方法探究miR -450a -3p 对Bub1 表达的调控机制,以及miR-450a-3p 对MEFs 增殖、凋亡、细胞周期和染色体数目的影响,为临床治疗自然流产、防治胚胎发育异常提供理论依据。
材 料 和 方 法
1 材料
1.1 细胞、质粒和腺病毒 HEK -293 细胞由重庆医科大学检验医学院分子医学研究室提供;表达Bub1 3’-UTR 荧光报告基因载体pMIR -3’、用于转染效率校正的表达半乳糖苷酶的荧光报告基因载体pMIR-REPORT β-gal 对照质粒、包含有与miR-450a-3p mimic 高度互补的MRE(miRNA response element)荧光报告基因载体MRE -450a -3p(5’-ATGAATGCAAAGCATCCCCAAT - 3’)和分别包含Bub1 3’-UTR 两种突变的萤光素酶报告基因载体Bub1 3’- UTR mut1(5’- ATGCTTAATGGACCTTCTCCAAAA-3’)、Bub1 3’-UTR mut2(5’-AATGAATATGGACCTTCTGGTTAA -3’)由本实验室前期利用Promega 双报告萤光素酶检测试剂盒构建;过表达Bub1 的重组腺病毒Ad-Bub1 由本实验室前期构建。
1.2 主要试剂和仪器 miR -450a -3p mimic 及阴性对照mimic 购自广州锐博生物科技有限公司,miR-450a -3p mimic 序列为5’- AUUGGGGAUGCUUUGCAUUCAU-3’,阴性对照mimic 序列为5’-UUCUCCGAACGUGUCACGUTT-3’;DMEM 培养基、胎牛血清、青霉素/链霉素购自HyClone,脂质体Li-
pofectamine 2000、Trizol 购自Invitrogen,逆转录PCR试剂盒、qRT - PCR 试剂盒购自TaKaRa,MTT、DMSO、ECL 试剂盒和秋水仙素购自Sigma,氯化钾、甲醇、冰醋酸购自重庆川东试剂公司,Hoechst 染色试剂盒购自北京晶美公司,Giemsa 染色试剂盒购自南京建成生物公司。Bub1 抗体和β - actin 抗体均购自Santa Cruz,HRP 标记的鼠抗羊IgG(用于Bub1 蛋白检测)和羊抗鼠IgG(用于β -actin 检测)均购自北京中杉金桥生物技术公司。qRT - PCR 仪购自Corbett,垂直电泳槽购自北京君意东方电泳设备有限公司,Western blotting 显影仪购自Bio -Rad,浓度测定仪购自Thermo,MTT 检测用酶标仪购自Sunrise,荧光素酶报告分析仪购自Promega。细胞培养皿、细胞培养板均购自Corning。
2 方法
2.1 MEFs 的制备、培养和分组 无菌条件下取14.5 胚龄(days postcoitum,dpc)的小鼠胚胎,移去胚胎的四肢和内脏,并弃之。留下头的下部,移去脑部或含有脑的头上部。剪碎后经胰酶消化后在37 ℃、5%CO2细胞培养箱中培养,培养基为DMEM 高糖培养基(加入10%胎牛血清、1 ×105U/L 青霉素、100 mg/L 链霉素)。取5 代以内对数生长期、生长状态良好的Mefs 细胞,随机分为3 组:(1)空白对照组:未做任何处理;(2)阴性对照组:转染control mimic组;(3)实验组:转染miR -450a -3p mimic 组。相同条件重复3 次。
2.2 qRT-PCR 检测miR -450a -3p mimic 转染前后miR-450a -3p 水平 Trizol 法分别提取3 组细胞总RNA,低温送至广州锐博生物科技有限公司进行qRT-PCR 检测成熟miR -450a -3p 的表达,每组实验重复3 次。
2.3 萤光素酶报告基因检测实验 将MEFs 以30%密度接种于6 孔板,按LipofectamineTM2000 说明转染实验干预因素,实验分组同前,同时共转染pMIR-3’,设3 个复孔。转染36 h 后弃去培养基,PBS 洗3 次,收获细胞,每孔加入200 μL 细胞裂解液,室温轻微振荡15 min。在荧光素酶报告分析仪专用检测板每孔加入30 μL 荧光素酶催化底物,再加入20 μL 细胞裂解产物后混匀,测定荧光素酶活性,每组实验重复3 次。所有数据都经过Promega 双报告系统pMIR -REPORT β -gal 对照质粒为内参校正后进行统计学分析。
2.4 Western blotting 检测miR-450a-3p mimic 转染前后Bub1 蛋白水平 接种MEFs 以60%密度接种于直径为10 cm 的细胞培养皿中,待细胞达到对数生长期,按LipofectamineTM2000 的说明转染实验干预因素,实验分组同前。继续常规培养,48 h 后收集细胞,提取细胞总蛋白,采用BCA 法进行蛋白定量。各取200 μL 样品上样进行SDS -PAGE,Ⅰ抗分别为Bub1抗体(1∶500)和β -actin 抗体(1∶1 000),Ⅱ抗分别为鼠抗羊IgG(1∶5 000)和羊抗鼠IgG(1∶5 000),最后按ECL 试剂盒说明进行电化学发光检测,并用Bio -Rad显影仪采集图像。
2.5 qRT-PCR 检测miR -450a -3p mimic 转染前后Bub1 mRNA 水平 Trizol 法分别提取3 组细胞总RNA,浓度仪测定样品纯度和含量,按逆转录试剂盒说明书提供的方法逆转录,逆转后的cDNA 作为PCR 的模板进行PCR 扩增。Bub1 上游引物5’-TGGTTGAACAAGTCCACAGC - 3’,下游引物5’-CTGACCCAGGTCAATCAATG - 3’。内参照选用小鼠的GAPDH,上游引物5’-GGCTGCCCAGAACATCAT-3’,下游引物5’-CGGACACATTGGGGGTAG-3’。反应体系为:cDNA 2 μL,SyberGreen Mix 10 μL,上、下游引物各0.5 μL,ddH2O 12 μL。反应条件为:95 ℃变性60 s;95 ℃30 s,59 ℃20 s,72 ℃20 s,共35 个循环。以Bub1 mRNA 拷贝数与内参照GAPDH mRNA 拷贝数的比值为Bub1 的相对表达量。
2.6 MTT 检测miR -450a -3p 对MEFs 增殖的影响 用0.25%胰蛋白酶消化细胞,以每孔500 个接种于96 孔板中,加入200 μL DMEM 完全培养基,常规培养待细胞贴壁后加入实验干预因素,另设一组同时经miR-450a-3p mimic 和Ad -Bub1 处理,共4 组。每组设置10 个复孔。于0 h、12 h、24 h、36 h、48 h 和60 h 分别加入20 μL MTT,37 ℃继续培养4 h,小心弃上清。再加入150 μL DMSO,振荡摇匀,酶标仪测定各孔490 nm A 值,重复3 次。
2.7 Hoechst 染色检测miR -450a -3p 对MEFs 凋亡的影响 用0. 25% 胰蛋白酶消化细胞,以每孔500 个接种于96 孔板中,加入200 μL DMEM 完全培养基,常规培养待细胞贴壁后加入实验干预因素,另设一组同时经miR -450a -3p mimic 和Ad - Bub1处理,共4 组。每组设置3 个复孔。48 h 后,每孔依次加入细胞染色缓冲液100 μL,Hoechst 染色液0.5 μL,冰浴30 min,PBS 洗涤1 次,荧光显微镜下计数300 个细胞。凋亡率(%)=凋亡细胞数/细胞总数×100%。
2.8 流式细胞术检测miR-450a-3p 对MEFs 细胞周期的影响 用0.25%、不含EDTA 的胰蛋白酶消化收集加入实验干预因素0 h、12 h、24 h、36 h、48 h和60 h 的3 组细胞,PBS 洗涤2 次后,重悬于预冷的70%乙醇。4 ℃保存,上流式细胞仪分析各组细胞的细胞周期(由重庆医科大学儿童医院儿研所完成检测)。每组设3 个平行样本,实验重复3 次。
2.9 染色体核型分析检测miR -450a -3p 对Mefs细胞染色体数目的影响 细胞传代24 h 后,经观察有较多分裂期细胞,加入终浓度为0.3 mg/L 的秋水仙素处理3 h,使分裂的细胞停止于有丝分裂中期。随后胰酶消化收集细胞,再经0.075 mol/L KCl 低渗膨胀细胞,减少染色体间的相互缠绕和重叠,最后用甲醇和冰醋酸将细胞固定于载玻片上,经Giemsa 染色后在显微镜下计数300 个细胞,观察染色体的结构和数量。
3 统计学处理
结 果
1 miR-450a-3p mimic 能够上调MEFs 中miR-450a-3p 的表达
经qRT-PCR 检测结果显示,加入外源性miR-450a-3p mimic,与阴性对照组相比,miR-450a -3p的表达明显增加4.4 倍(P <0.01),见图1。

Figure 1. The expression of miR-450a-3p in MEFs quantified by qRT - PCR. MEFs were transfected with miR -450a-3p mimic for 48 h. ±s.n =3. **P <0.01 vs control mimic group.图1 qRT-PCR 检测miR-450a-3p mimic 转染前后miR-450a-3p 水平
2 miR-450a-3p 通过靶作用于Bub1 3’-UTR调控Bub1 表达
萤光素酶报告基因检测结果显示,经pMIR -3’和miR-450a -3p mimic 共转染的MEFs 的萤光素酶活性比阴性对照组下降57% (P <0. 01),见图2A。经序列分析,我们发现Bub1 基因转录产物的3’-UTR 在终止密码子后与miR -450a -3p 有14个碱基互补,见图2C。
为了验证以上结果,将pMIR-3’与MRE-450a-3p 进行共转染,与阴性对照组相比,MEFs 的萤光素酶活性被抑制了90%左右(P <0.01),见图2B。同时,把Bub1 3’-UTR 与miR-450a-3p 结合的位点做了突变,见图2C,并进行转染,可以看到突变mut1 和mut2 均能够使萤光素酶活性明显升高,也就是抑制了miR-450a-3p 与Bub1 3’-UTR 的结合,见图2B。
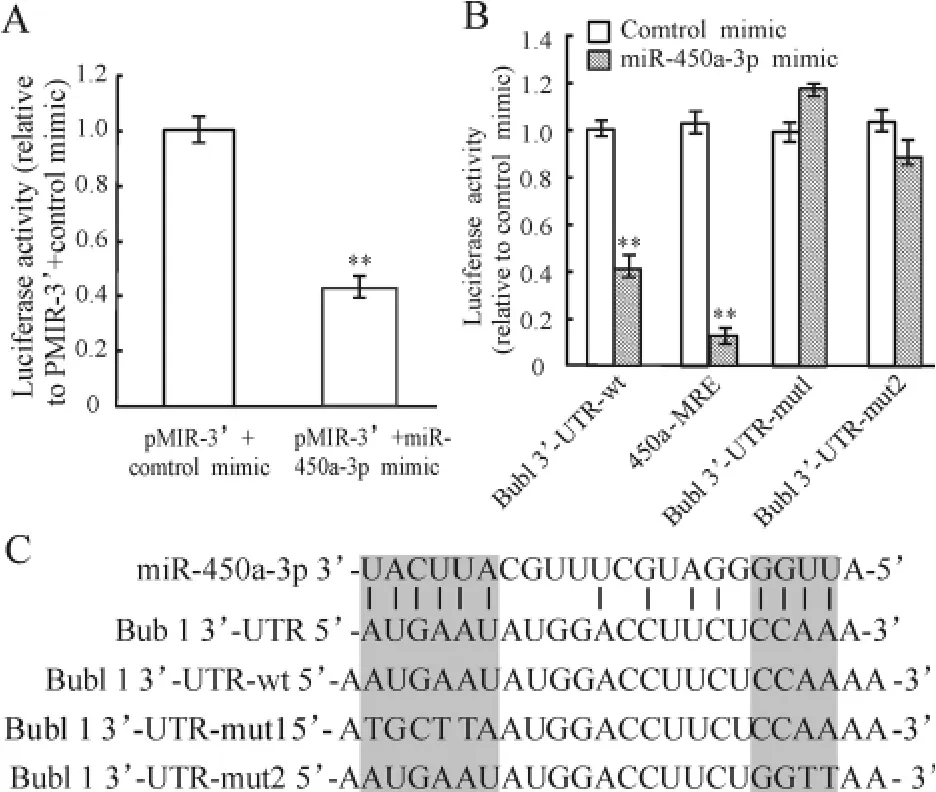
Figure 2. miR-450a-3p binds to the 3'-UTR of mouse Bub1 mRNA. A:luciferase activity of MEFs after cotransfected with pMIR - 3' vector and miR - 450a - 3p mimic. ±s.n=3. **P <0.01 vs pMIR -3'+control mimic group. B:luciferase activity of MEFs after transfected with Bub1 3' - UTR - wt,450a - MRE,Bub1 3'-UTR-mut1 or Bub1 3'-UTR-mut2 in the presence of miR-450a -3p mimic. ±s. n =3. **P<0.01 vs control mimic group. C:sequences of the miR-450a-3p binding sites within mouse Bub1 mRNA (Bub1 3'-UTR),wild -type 3'-UTR of Bub1(Bub1 3'-UTR -wt)and mutant 3'-UTR nucleotides of the miR-450a-3p binding sites (Bub1 3'-UTR-mut1 and Bub1 3'-UTR-mut2).图2 萤光素酶报告基因实验检测miR-450a-3p 与Bub1 3’-UTR 的结合
3 miR-450a-3p 干扰MEFs 中Bub1 蛋白水平的表达
Western blotting 检测MEFs 中Bub1 蛋白表达水平,结果显示,与空白对照相比,实验组细胞Bub1 蛋白明显减少(P <0.01),而阴性对照组与空白对照组之间差别不大,表明MEFs 转染miR-450a-3p 会使Bub1 蛋白水平明显减少,见图3。
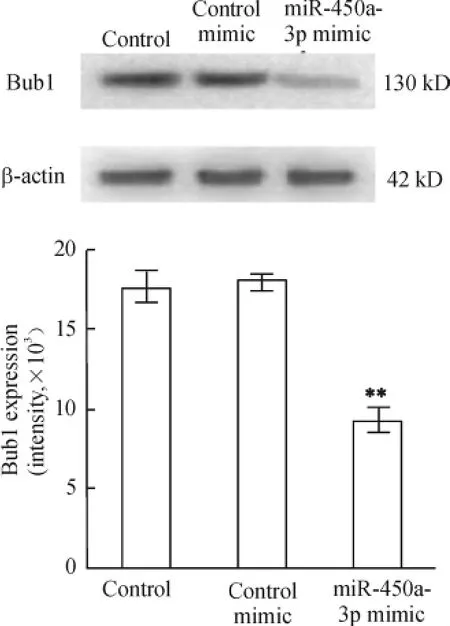
Figure 3. Bub1 protein expression in MEFs 48 h after transfected with miR-450a -3p mimic was detected by Western blotting. ± s. n = 3. **P <0. 01 vs control mimic group.图3 Western blotting 检测miR-450a-3p mimic 转染前后Bub1 蛋白的表达
4 miR-450a-3p 干扰MEFs 中Bub1 mRNA 水平的表达
qRT - PCR 检测MEFs 中Bub1 mRNA 表达水平,结果显示,与空白对照相比,实验组细胞和阴性对照组细胞Bub1 mRNA 表达水平的变化均不明显,无显著差异,表明MEFs 转染miR-450a-3p 并不会使Bub1 转录水平改变,见图4。
5 miR-450a-3p 对MEFs 增殖和凋亡的影响
MTT 法检测细胞增殖,结果显示,经mimic 处理后36 h 内,3 组细胞增殖率间没有明显差异,而60 h时与空白对照组相比,实验组细胞增殖率明显减低(P <0.01),而阴性对照组与空白对照组之间无显著差异。但在加入miR - 450a - 3p 的同时过表达Bub1,细胞增殖率相比较对照组不会出现明显的下降。这表明miR-450a-3p 可以抑制MEFs 的增殖,且加入外源性的Bub1 能干扰这种抑制,见图5A。
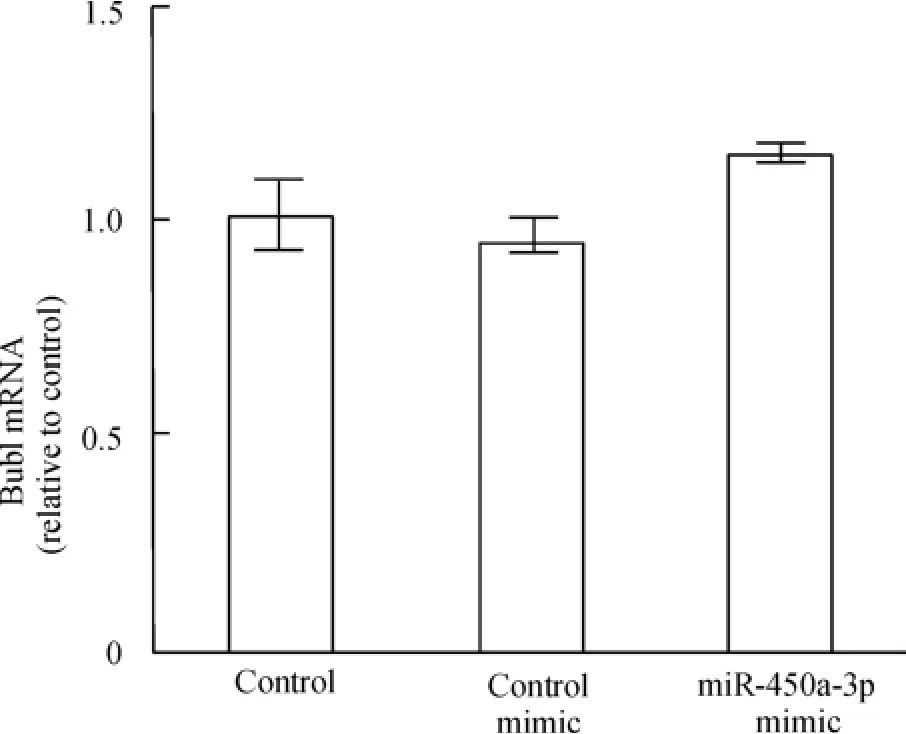
Figure 4. Bub1 mRNA expression in MEFs 48 h after transfected with miR -450a -3p mimic was detected by qRT -PCR. ±s.n=3.图4 qRT-PCR 检测miR-450a-3p mimic 转染前后Bub1 mRNA 的表达
Hoechst 染色检测细胞凋亡结果显示,经mimic处理后48 h,与空白对照组相比,实验组细胞凋亡的百分比明显增加,Hoechst 染色细胞阳性率由2.33%上升至7.85%(P <0.01)。而阴性对照组Hoechst染色细胞阳性率为2.58%,与空白对照组之间无显著差异。但在加入miR -450a -3p 的同时过表达Bub1,Hoechst 染色细胞阳性率相比较对照组不会出现明显的升高。这表明miR -450a -3p 可以引起MEFs 的凋亡,且加入外源性的Bub1 能干扰其促凋亡的作用,见图5B、C。
6 miR-450a-3p 对MEFs 细胞周期的影响
流式细胞术分析结果显示,经mimic 处理后48 h,与空白对照组相比,实验组细胞大多数被阻滞在G1/G0期,由35.89%上升至55.04%(P <0.01),而阴性对照组细胞G1/G0期占39.38%,与空白对照组之间无显著差异,见图6。这就说明在Mefs细胞中miRNA-450a-3p 所引起的Bub1 表达的下调,会导致SAC 功能异常,多数细胞过早地停止分裂,被阻滞在G1/G0期。

Figure 5. Effects of miR-450a -3p on proliferation and apoptosis of MEFs. A:the proliferation of MEFs was measured using the MTT;B:the apoptosis of MEFs was measured using Hoechst staining (×200);C:300 cells were counted in every experiment to calculate the Hoechst positive cell percentage. ±s.n=3. **P <0.01 vs control mimic group.图5 miR-450a-3p mimic 转染前后增殖和凋亡的变化

Figure 6. Effects of miR-450a-3p on cell cycle of MEFs. ±s.n=3. **P <0.01 vs control mimic group.图6 miR-450a-3p mimic 转染前后细胞周期的变化
7 miR-450a-3p 对细胞染色体数目的影响
染色体核型分析结果显示,与空白对照组相比,实验组细胞染色体数目异常率由4. 75% 上升至28.74%(P <0.01),而阴性对照组细胞染色体数目异常率为5.02%,与空白对照组之间无显著差异,见图7A。由miR -450a -3p 所引起的染色体数目异常有非整倍体和多倍体,其中非整倍体包括XXY 和2n-1,多倍体包括3n 和4n,见图7B。
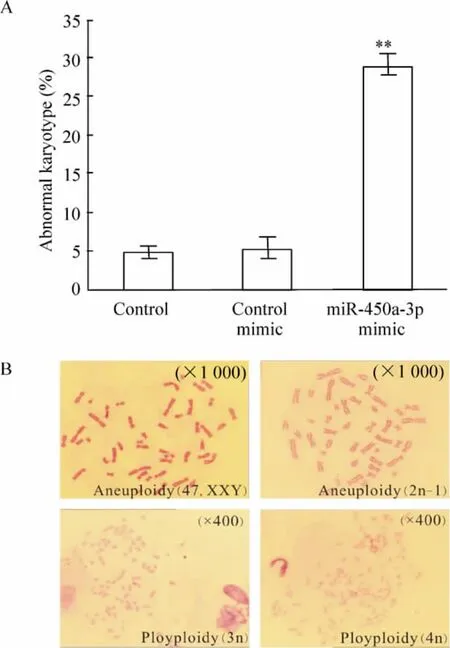
Figure 7. Abnormal karyotype in MEFs with overexpressed miR-450a -3p. A:abnormal karyotype cell percentage.±s.n=3. **P <0. 01 vs control mimic group. B:presentative abnormal karyotypes of MEFs in miR -450a-3p group (Giemsa staining).图7 miR-450a-3p mimic 转染前后染色质核型分析
讨 论
近年来,国内外对miRNA 的研究不断增多,人们逐渐认识到其在基因表达调控中的重要性。miRNA 是一类大小约21 ~23 个核苷酸的非编码单链小分子RNA,由pri-miRNA 经Drosha 处理和Dicer 剪切所生成[4]。通常miRNA 可识别特异的mRNA,并通过不完全配对靶结合到mRNA 的3’-UTR,影响靶向mRNA 的稳定性或抑制其翻译,进而调节靶基因的表达[5],这就揭示了一个崭新的转录后基因调控机制[6]。而对miRNA 的研究,大多数都是通过体外构建miRNA 的模拟体miRNA mimics 实现的。Bernstein 等[7]指出,敲除Dicer 这一miRNA 生成过程中重要的核酸酶会导致小鼠胚胎在7.5 dpc 前死亡。同时,Perera 等[3]证明了敲除Bub1 的小鼠胚胎在3.5 dpc 后也会死亡。这些结果说明miRNA 在胚胎干细胞的增殖分化和早期胚胎发育中起着重要作用,而这一作用很可能是通过靶作用于Bub1 来实现的。
Bub1 作为CP 的主要组件蛋白之一[8],可以通过调节相应蛋白的活性起到调节纺锤体微管和着丝粒连接的作用,在有丝分裂过程中监控染色体正确排列和等量分离,为细胞有丝分裂的正常进行提供保障。此前有学者指出Bub1 失调所致的SAC 病变在肿瘤形成过程中有重要作用[9]。我们在前期实验中发现Bub1 基因的缺失会导致小鼠胚胎细胞有丝分裂出现异常,这不仅会导致细胞染色体数目的异常,还会改变细胞周期、中止细胞分裂甚至细胞的死亡。同时,在对人类自然流产胚胎细胞的分析中也发现有明显的Bub1 下调以及染色体核型异常[10]。
本实验室在前期研究中,通过生物信息学方法发现有8 种miRNA 均能靶作用于Bub1 3’-UTR,分别是miRNA -30a、30e、297b、450a -3p、466a -3p、467a、467e 和494,其中miR -450a -3p 的作用尤为显著。miR-450a-3p 在多种组织中广泛表达,但对其生物学功能知之甚少,仅仅有研究指出其在子宫内膜癌肉瘤和前列腺癌[11]中有表达差异,但是差异都不大。在本实验中,以Bub1 基因作为主要研究对象,通过萤光素酶报告实验证实了miR -450a -3p可以下调pMIR -3’的荧光素酶活性,即靶作用于Bub1 3’-UTR。随后,我们又用了2 个方法来验证以上结论:(1)将MRE -450a -3p 与miR -450a -3p mimic 共转染HEK-293 细胞。MRE 主要存在于mRNA 的3’UTR 中,内含7 个碱基大小的特征性位点能与相对应miRNA 的5’端高度结合[12]。我们可以看到共转染MRE -450a -3p 能明显抑制荧光素酶活性。(2)将Bub1 3’-UTR 区域的2 个miR -450a-3p 结合位点突变,构建了突变体萤光素酶报告基因载体,与miR -450a -3p mimic 转染HEK -293 细胞,与pMIR -3’相比,萤光素酶活性明显恢复,证明miR-450a-3p 通过与Bub1 3’-UTR 的结合位点相结合发挥作用。
我们采用Western blotting 和qRT - PCR 的方法,发现实验组的Bub1 蛋白水平有明显降低,但是mRNA 水平受实验干预因素的影响不大。这可能是因为miR-450a -3p 作为一个翻译抑制子,主要是对基因表达翻译水平的调节,但却不会影响靶基因mRNA 水平的降解。但同时也有学者指出,有一些miRNA 会诱导靶作用mRNA 的降解从而抑制蛋白质合成[13],且miR-10a 能够通过与核糖体蛋白mRNA 5’-UTR 结合增强翻译,而不是抑制翻译[14]。因此miRNA 调控基因表达方式的不同,也成为miRNA 研究中一个急需解决的问题。
本文结果同时也表明,过表达miR -450a -3p会通过下调Bub1 的表达而抑制MEFs 的增殖,这与Perera 等[3]指出的Bub1 缺失会影响MEFs 增殖的结论是一致的。究其抑制增殖的原因可能有2 个:(1)促进Mefs 细胞的凋亡;(2)将细胞大量阻滞在G1/G0期,使细胞过早地停止分裂。同时经过染色体核型分析,可以看到非整倍体、多倍体等异常核型频率增高,这与Perera 等[3]指出的Bub1 缺失致有丝分裂异常的结论也是一致的。
综上所述,miR -450a -3p 能够通过靶结合到Bub1 3’-UTR 并干扰Bub1 的表达,抑制胚胎细胞的增殖和发育,导致细胞染色体数目异常。为研究因染色体数目异常所导致的自然流产及产前诊断提供了新的思路,为提高临床妊娠质量奠定了基础。
[1] 雷 琼,王 琼,周灿权,等. 不同分子遗传学方法用于自然流产绒毛细胞遗传分析的效果[J]. 中华妇产科杂志,2009,44(4):253 -256.
[2] Rieder CL,Cole RW,Khodjakov A,et al. The checkpoint delaying anaphase in response to chromosome monoorientation is mediated by an inhibitory signal produced by unattached kinetochores [J]. J Cell Biol,1995,130(4):941 -948.
[3] Perera D,Tilston V,Hopwood JA,et al. Bub1 maintains centromeric cohesion by activation of the spindle checkpoint[J]. Dev Cell,2007,13(4):566 -579.
[4] Cai X,Hagedorn CH,Cullen BR. Human microRNAs are processed from capped,polyadenylated transcripts that can also function as mRNAs[J]. RNA,2004,10(12):1957-1966.
[5] Zhang R,Su B. Small but influential:the role of microRNAs on gene regulatory network and 3'UTR evolution[J].J Genet Genomics,2009,36(1):1 -6.
[6] Bartel DP. MicroRNAs:genomics,biogenesis,mechanism,and function [J]. Cell,2004,116(2):281 -297.
[7] Bernstein E,Kim SY,Carmell MA,et al. Dicer is essential for mouse development[J]. Nat Genet,2003,35(3):215 -217.
[8] Boyarchuk Y,Salic A,Dasso M,et al. BUB1 is essential for assembly of the functional inner centromere[J]. J Cell Biol,2007,176(7):919 -928.
[9] Masuda A,Takahashi T. Chromosome instability in human lung cancer:possible underlying mechanism and potential consequences in the pathogenesis[J]. Oncogene,2002,21(45):6884 -6897.
[10] 施 琼,袁泰先,王 箭,等. 有丝分裂关卡基因致染色体数目异常自然流产胚胎发生的机制[J]. 中华检验医学杂志,2008,21(3):309 -315.
[11] Castilla MÁ,Moreno -Bueno G,Romero - Pérez L,et al. Micro-RNA signature of the epithelial-mesenchymal transition in endometrial carcinosarcoma[J]. J Pathol,2011,223(1):72 -80.
[12] Nahvi A,Shoemaker CJ,Green R. An expanded seed sequence definition accounts for full regulation of the hid 3’UTR by bantam miRNA[J]. RNA,2009,15(5):814 -822.
[13] Liu Q,Fu H,Sun F,et al. MiR-16 family induces cell cycle arrest by regulating multiple cell cycle genes[J].Nucleic Acids Res,2008,36(16):5391 -5404.
[14] Ørom UA,Nielsen FC,Lund AH. MicroRNA-10a binds the 5’UTR of ribosomal protein mRNAs and enhances their translation[J]. Mol Cell,2008,30(4):460 -471.
