朱鹮听觉发声中枢和内分泌核团内P物质的定位
2012-12-07张学博张信文孙颖郁曾少举
左 夺,张学博*,张信文,孙颖郁,曾少举,刘 斌
(1.海南师范大学 生命科学学院,海南 海口 571158;
2.北京师范大学 生命科学学院,北京 100875;
3.北京动物园,北京 100044)
朱鹮听觉发声中枢和内分泌核团内P物质的定位
左 夺1,张学博1*,张信文1,孙颖郁2,曾少举2,刘 斌3
(1.海南师范大学 生命科学学院,海南 海口 571158;
2.北京师范大学 生命科学学院,北京 100875;
3.北京动物园,北京 100044)
朱鹮为世界濒危物种、用免疫组织化学方法对朱鹮脑内听觉-发声中枢和内分泌核团中的P物质免疫阳性神经细胞、终末和纤维进行了定位:1)在听觉通路中的延髓耳蜗核、脑桥腹部外侧丘系核、中脑背外侧核壳、中脑丘间核壳、丘脑卵圆核壳等有P物质阳性纤维和终末;在脑桥橄榄核有P物质阳性神经细胞;2)在发声通路中的气管鸣管亚核、中脑丘间复合体背内侧核、丘脑前背外侧核内部、古纹状体腹部中间区有P物质阳性神经细胞和终末;3)在视前区前核、下丘脑外侧核,下丘脑腹内侧核等内分泌核团有P物质阳性神经细胞.结果提示:P物质在听觉通路和听觉旁通路、发声通路和生殖内分泌相关的核团中表达丰富,鸟脑中的P物质可能参与听觉和鸣叫行为以及交配和生殖等生理功能的调制.
朱鹮;P物质;发声中枢;听觉中枢;内分泌核团
朱鹮(Nipponia nippon)为世界濒危物种[1],国内外已对朱鹮的生态学及保护生物学等方面做了较详细的研究[2-3],但对其脑的解剖等的研究很少,对其脑内P物质(substance P,SP)的定位,国内外尚无报道.P物质是氨基酸类的神经调质,在鸟类的脑活动中起一定的调制作用,可以调制多种神经递质的释放,以此参予多种脑活动的调节[4-6].本实验用P物质免疫组化技术和图像仪分析系统,对P物质在朱鹮听觉发声中枢和内分泌核团内的分布做了定位分析.
1 材料和方法
1.1 焦油紫染色
实验动物:朱鹮脑(♂1只,意外死亡)由北京动物园赠送.将脑剥离出来,放入4%多聚甲醛固定2d后转入30%的蔗糖液.在低温恒冷切片机下把脑切成40 μm的切片,贴于用明胶处理过的载玻片上,用0.1%的焦油紫染色后进行脱水脱脂,将切片制成永久切片,于显微镜下镜检.
1.2 P物质-DAB免疫组化流程
0.01 mol/L PBS漂洗,5min×3次,3%正常羊血清(TritonX-100/PBS稀释)室温封闭30 min,兔抗P物质(中杉金桥,1∶200),4℃孵育过夜,室温反应2h,0.01 mol/L PBS漂洗,5min×3次,生物素标记的羊抗兔IgG(Vector,1∶400)室温反应2h,0.01 mol/L PBS漂洗,5min×3次,ABC复合物(Vector,1∶200),室温反应2h,0.01 mol/L PBS漂洗,5min×3次,DAB(Diaminobenzidine tetrahydra chloride)(6%)和30%H2O2(0.6%)的Tris-Cl稀释工作液避光显色30-40 min,蒸馏水漂洗、贴片、脱水、透明、封片.
1.3 图像采集与数据分析
应用Olympus明视野光学显微镜以及Cool-SNAP数码照相系统,采集X区的P物质免疫染色图像,朱鹮核团的定位和命名参考鸽、金丝雀脑立体定位图谱和张信文等以前的工作[7-9]为依据.选取核团的中央截面及内外各200 μm截面共3张脑片进行统计,在每张脑片核团内部和周围区域分别取3个不同的约500 μm2的长方形,用图像分析软件(Image Pro Plus软件)分别计算其灰度(gray value),我们将3张截面核团周围区域内9个选定区域的平均灰度值减去核团内部共9个选定区域的平均灰度值的差值定义为消减平均灰度(sub⁃tracted mean gray value),即背景物平均灰度与阳性物平均灰度之差作为统计对象.经SPSS 13.0软件的K-S检验确认各组数据为正态分布后,应用统计分析软件Prism 4进行单因素方差分析(LSD),比较多组间差异,实验结果列表1,当P<0.05时差异显著;当P<0.01时差异极显著.
2 结果
2.1 延髓和脑桥内P物质的定位
在朱鹮的延髓舌下神经核(nXII)和气管鸣管亚核,舌下神经(NXII)内有许多P物质标记的神经终末和纤维(见图1-1),舌下神经核和气管鸣管亚核呈不规则形,位于延脑的背侧,矢状切面积约1.26 mm2.在朱鹮的延髓耳蜗核(nCo)和角状核有浓重的P物质免疫阳性神经终末(见图1-2,表1),耳蜗核和角状核位于第四脑室的腹侧,延髓的背外侧,矢状切面积约1.09 mm2.在听神经(NⅧ)有一些P物质阳性神经纤维(见图1-2).在脑桥的橄榄核(SO)有P物质的免疫阳性细胞(见图1-3),橄榄核位于脑桥尾部的腹侧缘,呈球形,矢状切面积为0.69 mm2.在脑桥腹部外侧丘系核(VLV)有P物质阳性神经终末(见图1-4).

表1 朱鹮脑内SP免疫阳性物质的部分灰度值Tab.lThe density of the SP positive substances in the brain in Nipponia nippon
2.2 中脑和间脑内P物质的定位
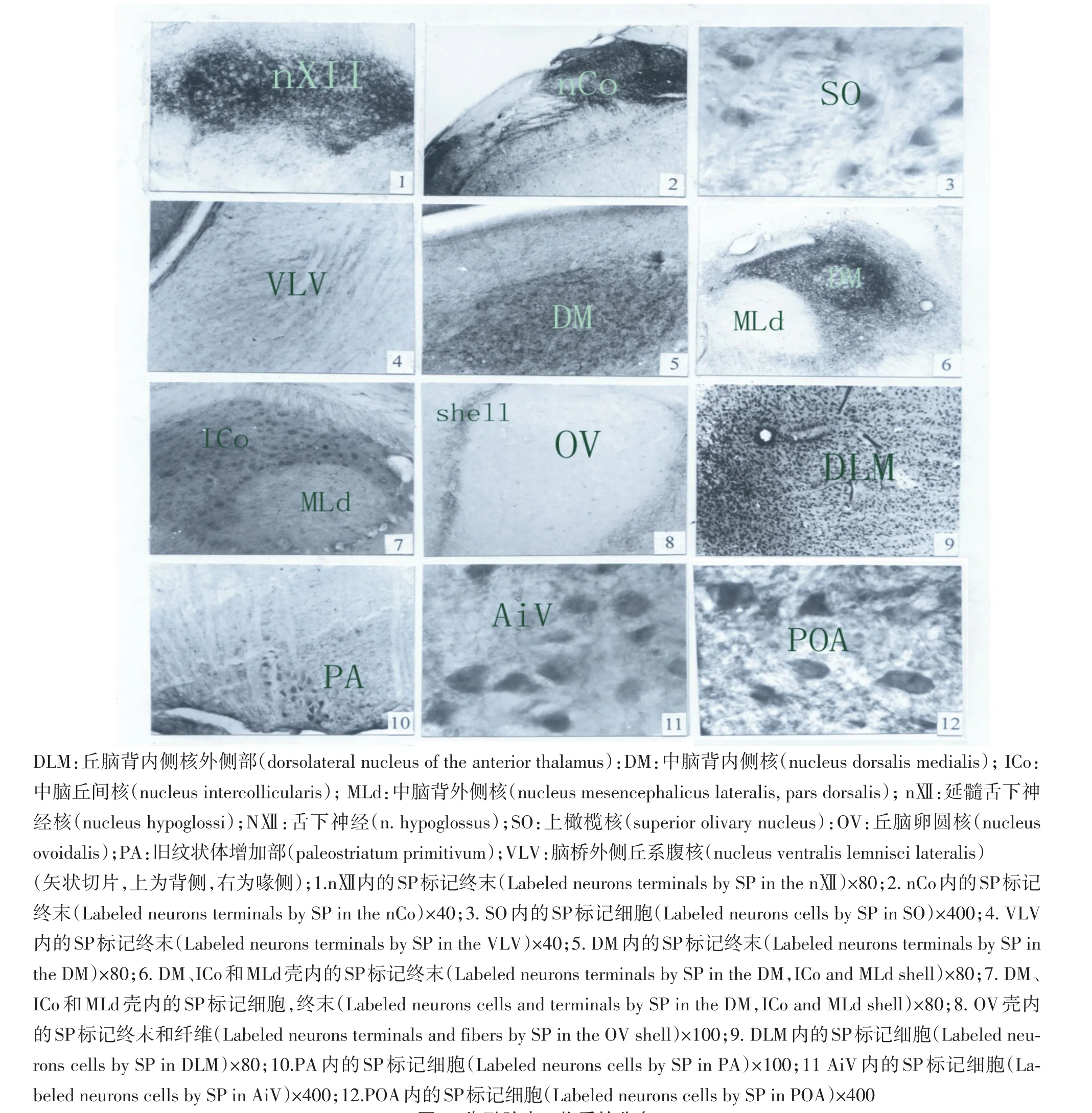
图1 朱鹮脑内P物质的分布Fig.1 The distribution of substance Pin the brain of Nipponia Nippon
在中脑主要观察到:在中脑丘间复合体背内侧核(DM)有密集的P物质免疫阳性神经终末,矢状切面积约0.64 mm2(见图1-5).在中脑背外侧核(MLd)壳(见图1-6,表1)和中脑丘间核(ICo)壳有许多P物质免疫阳性终末和细胞,浓密的标记纤维分布在ICo和MLd周围区(见图1-7).ICo壳分为内侧丘间核(mICo)和外侧丘间核(lICo),通过焦油紫染色将ICo壳、DM和MLd壳区分开来.标记细胞、终末与纤维分布在中脑背外侧核壳上(见图1-6,7).在卵圆核壳(OV-Shell)有许多浓重的P物质免疫阳性神经终末和纤维(见图1-8,表1),OV-Shell包括OV核前部(Ova),Ov核后部(Ovp).在下丘脑腹内侧核(VMN)、VMN-Shell区有许多浓重的P物质免疫阳性细胞和纤维(见表1).下丘脑外侧区内有许多浓重的P物质免疫阳性细胞(见表1).在丘脑前背外侧核内部(DLM)有许多浓重的P物质免疫阳性椭圆形细胞和终末(见图1-9,表1).
2.3 端脑内P物质的定位
在旧纹状体增加部(PA)内有许多的P物质神经细胞(见图1-10),位于LMD下方.在古纹状体腹部中间区(AiV)内有许多浓重的P物质免疫阳性神经细胞(见图1-11,表1).在视前区前核(POA)有浓重的P物质免疫阳性神经细胞和纤维(见图1-12,表1).
3 讨论
P物质是兴奋性神经肽,也是一种脑肠肽,属于速激肽家族一类,具有相似羧基端氨基酸序列和生物活性的神经肽,广泛地分布在中枢和周围神经组织中,具十一肽序列Arg1-Pro2-Lys3-Pro4-Gln5-Gln6-Phe7-Phe8-Gly9-Leu10-Met11-NH.P物质在中枢神经系统中参与多种生理过程,如疼痛,学习和记忆等,主要存在于中脑、下丘脑、视前区、基底神经节及边缘系统[4-6].本实验结果表明,P物质在朱鹮脑中分布于听觉通路和听觉旁通路、发声控制系统和生殖内分泌相关的核团等三个系统中表达丰富,这些提示P物质可能与鸟类的听觉和鸣叫行为等多种生理功能有关.
3.1 听觉通路中的P物质定位
鸟类的听觉通路主要由听神经(NⅧ)先投射到延髓耳蜗核的角状核和巨细胞核团,再投射到层状核、脑桥橄榄核和脑桥腹部外侧丘系核,然后再投射到中脑背外侧核、丘脑卵圆核,最后投射到端脑L听区[10-11],另外由中脑MLd壳、ICo壳、丘脑OV壳和新纹状体旁听区等组成旁听觉通路[12].本实验在朱鹮的听觉通路和旁听觉通路的多数核团:听神经、延髓耳蜗核、脑桥橄榄核和脑桥腹部外侧丘系核、中脑MLd壳、ICo壳和丘脑OV壳内有许多的P物质免疫阳性神经细胞和纤维,这些结构都参与听觉中继的生理反应[13].P物质大量分布在MLd壳、OV壳等核团的壳上,说明P物质在旁听觉通路中可能起重要的调制作用.
3.2 发声控制通路中的P物质定位
P物质广泛分布于发声控制通路中,已知鸟类的鸣叫由脑内发声控制通路所控制,鸟类的发声通路由端脑古纹状体腹部中间区→中脑丘间复合体背内侧核→气管鸣管亚核,然后支配气管鸣管肌[10-11,14].本实验结果表明,在朱鹮发声控制通路脑区古纹状体腹部中间区、中脑丘间复合体背内侧核和气管鸣管亚核富含P物质胞体及纤维,提示P物质作为神经调质,可能通过兴奋其他神经递质或受体,从而在发声控制等功能中发挥一定的调制作用.
3.3 P物质在听觉-发声和生殖内分泌核团中的定位
已知鸟类求偶炫耀,交配和生殖活动与生殖内分泌有关的脑区,包括视前区前核、下丘脑外侧核,下丘脑腹内侧核和下丘脑的各个亚核有着密切的关系[12].而鸟类的听觉和发声,以及生殖内分泌三个系统之间有密切的联系,其中听觉高位中枢L听区投射到发声高位中枢[10-11],听觉反馈调控发声行为[15-16].下丘脑的视前区前核、下丘脑外侧核,下丘脑腹内侧核与旁听觉通路有直接的神经联系,由此听觉和发声以及生殖内分泌神经核团组成了发声—听觉—内分泌下丘脑轴系统[10-12].本实验结果表明,在发声—听觉—内分泌下丘脑轴系统内均有P物质免疫阳性神经细胞或纤维的分布,提示鸟脑中的P物质可能与听觉和鸣叫行为,以及交配和生殖活动等多种生理调控有关.其发声—听觉—内分泌下丘脑轴系统的调控机制还有待于进一步深入的研究.
[1]刘荫增.朱鹮在秦岭的重新发现[J].动物学报,1981,27(3):273.
[2]范光丽,吴建云,马新武.朱鹮的前后肢比较解剖学观察[J].动物医学进展,1999,20(2):34-37.
[3]Yamasina Y,Nakanishi G.Nipponia nippon[M].Tokyo:Newton Books,1983.
[4]Nakanishi S.Mammalian tachykinin receptors[J].Annu.Rev Neurosic,1991,14(1):123-126.
[5]Otsuka Y K.Neurotransmitter functions of mammalian tachykinins[J].Physiol Rev,1993,73(2):229-308.
[6]Liu S J,Ju G.Substance P-link immunoreactive nerve fi⁃bers in the pars distalis of the adenohypophys is of macaque monkeys[J].Neurosci Lett,1998,94:1-4.
[7]张信文,蓝书成.黄雀上纹状体腹侧尾核的中枢联系—HRP法研究[J].解剖学报,1993,24(4):381-390.
[8]张信文,蓝书成.家鸽、黄雀和黄喉鵐、耳蜗核的定位与比较[J].动物学报,1994a,40(1):51-57.
[9]张信文,蓝书成.黄雀、黄喉鵐角状核和层状核的听觉通路及比较[J].动物学报,1994b,40(4):390-398..
[10]张信文,蓝书成.黄雀古纹状体栎核的中枢联系—HRP法研究[J].解剖学报,1994a,25(1):20-26.
[11]张信文,蓝书成.黄雀新纹状体前部巨细胞核及嗅叶X区的纤维联系[J].解剖学报,1994b,25(2):146-149.
[12]Cheng M F,Zuo M X.Proposed pathway for vocal self-stimulation:Met-enkephalinergic projections linking the midbrain vocal nucleus,auditory responsive thalamic regions and neurosecretory hypothalamus[J].J Neurobi⁃ol,1994,5:361.
[13]Durand S E,Liang W,Brauth S E.Methione enkephhalin immunoreactivity in the brain of the bundgerigar(melop⁃sittacus):Similarities and differences and differences with respect to oscine songbirds[J].Comp Neurol,1998,393(2):145-168.
[14]Veenman C L,Wild J M,Reiner A.Organization of the avian"corticostriatal"projection system:Aretrograde and anterograde pathway tracing study in pigeons[J].Comp Neurol,1995,354(1):87-102.
[15]Vates G E,Broome B M,Mello C V,et al.Auditory path⁃ways of caudal telencephalon and their relation to the song system of adult male zebra finches(Taenopygia gutta⁃ta)[J].Comp Neuro,1996,366(5):613-642.
[16]Vates W,Vicario D S,Nottebohm F.Reafferent thalamo"Cortical"loops in the song system of oscine songbirds[J].Comp Neurol,1997,380:275-290.
The Localization of Substance P in the Vocal Center and the Audi⁃tory Center and Endocrine Nuclei in the Brain of Nipponia Nippon
ZUO Duo1,ZHANG Xuebo1*,ZHANG Xinwen1,SUN Yingyu2,ZENG Shaoju2,LIU Bin3
(1.College of Life Science,Hainan Normal University,Haikou571158,China;
2.College of Life Science,Beijing Normal University,Beijing100875,China;
3.Zoo Beijing,Beijing100044,China)
The distribution of substance P in the brain ofNipponia nipponwas investigated by use of immunohisto⁃chemical methods,and the gray density of numerical value was measured with the image processing system.1)The sub⁃stance P labeled terminal and cells were distributed in the auditory center such as the nCo,superior olivary nucleus(SO),nucleus ventralis lemnisci lateralis(VLV),the shell regions of mesencephalicus lateralis,pars dorsalis(MLd shell),the nucleus ovoidalis shell(Ov shell)et al;2)In the vocal control nuclei such as the nucleus hypoglossi,pars tracheosyringealis(nXIIts),nucleus dorsalis medialis(DM)and the nucleus intercollicularis(ICo),dorsolateral nucle⁃us of the anterior thalamus(DLM),AiV of Nipponia nippon.3)In the endocrine nuclei such as nucleus ventromedialis hypothalamic(VMN),nucleus lateralis hypothalamic(LHy),nucleus preopticus anteror(POA)et al;The results indi⁃cate that substance P may play an important physiological role in the auditory-vocal-endocrine hypothalamic axis.
Nipponia nippon;Substance P;Vocal center;Auditory center;Endocrine nuclei
Q 958
A
1674-4942(2012)01-0081-05
2011-12-20
国家自然科学基金资助项目(31160205)
*通讯作者
黄 澜
