三磷酸腺苷对N9小胶质细胞的损伤作用及其机制*
2012-11-06王国红郭直岳尹雅玲魏林郁李新娟李东亮
王国红, 郭直岳, 尹雅玲, 魏林郁, 李新娟, 李东亮,2△
(1新乡医学院生理学与神经生物学教研室,河南 新乡 453003; 2复旦大学医学神经生物学国家重点实验室,上海 200032)
1000-4718(2012)09-1597-08
2012-03-12
2012-07-11
河南省2009年科技发展计划项目( No.092102310098);复旦大学神经生物学国家重点实验室开放课题(No.10-10);新乡医学院重点领域开放课题(No.ZD2011-2)
△通讯作者 Tel: 0373-3029104; E-mail: xyldl8@126.com
三磷酸腺苷对N9小胶质细胞的损伤作用及其机制*
王国红1, 郭直岳1, 尹雅玲1, 魏林郁1, 李新娟1, 李东亮1,2△
(1新乡医学院生理学与神经生物学教研室,河南 新乡 453003;2复旦大学医学神经生物学国家重点实验室,上海 200032)
目的研究三磷酸腺苷(ATP)对N9小胶质细胞的损伤作用及其机制。方法取对数生长期N9小胶质细胞,随机分为3组:(1)正常对照组:常规培养,不进行ATP处理; (2)ATP组:接种24 h后行ATP处理; (3)KN-62 (P2X7受体阻断剂)干预组: KN-62孵育30 min后行ATP处理,且KN-62存在于ATP作用整个过程。XTT法测各组N9小胶质细胞的活力;倒置相差显微镜观察细胞形态变化;流式细胞术检测细胞周期及凋亡;细胞免疫荧光检测P2X7受体表达;Western blotting检测细胞P2X7受体蛋白水平;ELISA检测细胞上清液中白细胞介素1β(IL-1β)含量。结果500 μmol/L ATP和1 mmol/L ATP对细胞活力损伤程度较小,作用24 h后,细胞活力仍可达88.5%±5.5%和88.2%±8.4%。当ATP浓度达到或高于2 mmol/L时,细胞活力迅速降低,细胞发生皱缩,且随ATP作用时间延长,细胞活力逐渐降低,细胞密度逐渐减少,细胞皱缩程度加重,而KN-62干预后细胞活力较ATP组明显增多,细胞密度及形态明显好于ATP组。细胞周期及凋亡检测结果显示,ATP可使细胞周期阻滞于S期,细胞凋亡比例明显较对照组增多(P<0.01),KN-62干预可显著减轻ATP引起的细胞周期阻滞,细胞凋亡比例显著减少(P<0.01)。免疫荧光显示,ATP及KN-62干预对N9小胶质细胞上P2X7受体的表达分布没有影响。Western blotting显示,正常对照组、ATP组及KN-62干预组间P2X7受体蛋白水平没有明显差异(P>0.05)。ELISA结果显示,ATP及KN-62干预对IL-1β的释放没有作用。结论高剂量ATP可诱发N9小胶质细胞损伤,其机制可能与P2X7受体介导的细胞周期阻滞和细胞凋亡有关,而与小胶质细胞的炎症反应无关。
三磷酸腺苷; 小胶质细胞; 受体,嘌呤能P2X7; KN-62
三磷酸腺苷(adenosine triphosphate, ATP)是胞内能量代谢的主要储能和供能形式,由线粒体产生,正常情况下,释放到胞外的ATP量很少,为高纳摩尔级或低微摩尔级。当脑受到损伤(如缺血缺氧损伤)时,大量ATP释放到胞外,使胞外的ATP水平迅速升高,可达毫摩尔级,继而引发系列反应,导致细胞凋亡坏死,引起继发性脑损伤[1]。早在1998年,Francesco就曾将ATP命名为死亡因子,然而近些年研究发现[2-4],胞外ATP对有些细胞具有促增殖、抗凋亡的营养作用,而对另一些细胞则具有促进凋亡的损伤作用,甚至对同一种细胞在不同的作用时间分别表现出促增殖和促凋亡两种截然相反的效应,不仅如此,ATP发挥作用所涉及的机制和受体也各有不同。介导ATP作用的P2受体现已克隆出15个亚型,其中P2X7受体亚型近年来成为研究热点,研究发现P2X7受体在神经系统广泛存在并发挥重要作用,与脑缺血、癫痫、阿尔茨海默病、神经性疼痛等多种神经系统疾病密切相关,P2X7受体参与脑缺血后神经细胞的死亡及小胶质细胞的炎症反应[5-6]。小胶质细胞是中枢神经系统内固有的免疫效应细胞,目前被认为在脑损伤尤其是缺血性脑损伤中发挥着重要作用[7]。本研究采用不同剂量ATP处理N9小胶质细胞并结合P2X7受体阻断剂KN-62的使用,观察不同时间细胞活力、细胞形态、细胞周期和凋亡水平、P2X7受体表达和水平及胞外白细胞介素1β(interleukin-1β,IL-1β)含量的变化,以研究和探讨胞外ATP水平对小胶质细胞的作用及其所涉及的机制。
材 料 和 方 法
1材料
1.1主要试剂 四氮唑复合物{2,3-bis(2-methoxy-4-nitro-5-sulfophenyl)-5-[(phenylamino) carbonyl]-2H-tetrazolium hydroxide, XTT}、硫酸酚嗪甲酯(phenazinemethosulfate, PMS)、IMDM(Iscove’s modified Dulbecco’s medium)、碘化丙啶(propidium iodide, PI)、P2X7受体阻断剂KN-62均购自Sigma;RNase A (DNase free)、DAPI染色液、免疫荧光染色试剂盒——抗兔Alexa Fluor 488均购自碧云天生物技术研究所;兔抗鼠P2X7受体多克隆抗体购自Santa Cruz;小鼠IL-1β ELISA试剂盒购自上海依科赛生物制品有限公司;胎牛血清购自杭州四季青公司。
1.2细胞系 N9小胶质细胞由上海中国科学院惠赠。
2方法
2.1母液和工作液配制 ATP用无菌三蒸水配成浓度为500 mmol/L ATP母液,过滤后分装,避光保存于-20 ℃,用前培养基稀释成所需浓度的ATP工作液;KN-62用无菌DMSO溶解成浓度为1 mmol/L KN-62母液,分装后避光保存于-20 ℃,用前培养基稀释成特定浓度的KN-62工作液;培养基中加入特定量的ATP和KN-62母液配成KN-62干预工作液(含ATP)。
2.2细胞培养及实验分组 N9小胶质细胞采用IMDM培养基培养,内含5%胎牛血清、1×105U·L-1氨苄青霉素和1×105U· L-1链霉素,置于37 ℃、体积分数5%CO2培养箱内常规培养,取对数生长期细胞进行实验。实验分为正常对照组、ATP组和KN-62干预组。正常对照组:IMDM培养基常规培养;ATP组:细胞接种24 h后,更换为特定摩尔浓度的ATP工作液;KN-62干预组:ATP处理前30 min加入KN-62工作液,孵育30 min后,更换为KN-62干预工作液。
2.3XTT法测细胞活力 取对数期细胞,调整细胞浓度为2×108cells·L-1,接种于96孔培养板,每孔100 μL细胞悬液,按上述方法分组,每组设5个复孔,ATP处理一定时间后,用磷酸盐缓冲液现配XTT溶液(1.00 g·L-1)及PMS溶液(0.15 g·L-1),二者按一定比例混合后,每孔加入50 μL XTT/PMS混合液,上述配液及加液过程需避光操作,之后置培养箱中37 ℃孵育2 h,酶标仪测定450 nm波长处吸光度(A)。根据下列公式计算各组细胞存活率:存活率(%)=A处理组-A空白组/A对照组-A空白组×100%,实验重复3次。
2.4形态学观察 将培养板置于倒置相差显微镜下观察各组细胞形态变化,各组均取板孔正中视野进行观察并拍照。
2.5流式测细胞周期及凋亡 取对数生长期细胞,调整细胞浓度为3×108cells·L-1,接种于培养瓶中,24 h后行各组处理,ATP作用3 h和6 h后,收集各组培养液,平衡盐缓冲液漂洗2次并收集缓冲液,离心收集培养液及缓冲液中的细胞,胰酶消化收集瓶中其余贴壁细胞,1 000 r/min 离心5 min后弃去上清液,PBS重悬洗涤2次,细胞沉淀重悬于0.5 mL PBS缓冲液中,缓慢加入5 mL冰预冷的75%乙醇,4 ℃固定过夜。细胞上机检测前先离心去乙醇,PBS洗涤2次,加入100 mg/L的 RNase A(DNase free)重悬细胞,37 ℃孵育30 min以消化RNA,离心后加入50 mg/L 的 PI 4 ℃避光染色30 min,200目滤网过滤,然后用流式细胞术进行细胞周期时相动力学及细胞凋亡检测,根据所测得的DNA 分布图用细胞周期及凋亡分析软件进行细胞周期及凋亡统计分析。
2.6免疫荧光细胞化学染色 取对数生长期细胞,调整细胞浓度为1×108cells·L-1,接种于6孔板内,板孔内事先放有灭菌处理的长条玻片,进行爬片,24 h后行各组处理,于不同时点取出爬片,用预热至37 ℃的PBS液漂洗3次,每次2 min,洗掉残余培养液,免疫染色固定液常温固定10 min。PBS 洗涤3次,每次5 min,0.1% Triton X-100作用5 min,PBS洗涤3次,每次5 min,3% BSA室温孵育20 min,甩去多余液体不洗,滴加P2X7受体Ⅰ抗(PBS1∶100稀释)4 ℃过夜,PBS漂洗3次,每次5 min,滴加Alexa Fluor 488荧光标记Ⅱ抗(免疫荧光染色Ⅱ抗稀释液1∶500稀释) 4 ℃避光孵育60 min,PBS漂洗3次,每次5 min,滴加DAPI染色液室温复染核5 min,PBS漂洗3次,每次5 min,抗荧光淬灭封片液封片,荧光显微镜下观察拍照,随机选取5个不同视野用Image-Pro Plus 6.0专业图像分析软件分析,得到的平均吸光度表示该视野平均荧光强度。
2.7Western blotting测P2X7受体蛋白水平 各组细胞用冰预冷PBS洗涤2次,加入细胞裂解液,冰上裂解30 min后,用细胞刮刀收集细胞,离心取上清,用BCA法测定蛋白质浓度。各组取变性蛋白30 μg进行SDS-PAGE电泳分离蛋白质后,转至PVDF膜上,经漂洗、封闭后加入兔源小鼠P2X7受体多克隆抗体(稀释度为1∶200),4 ℃孵育过夜,TBST洗膜3次,每次15 min,加碱性磷酸酯酶标记的山羊抗兔Ⅱ抗37 ℃孵育1 h,TBST洗膜3次,每次15 min,BCIP/NBT显色并扫描,以Band Scan 5.0凝胶图像处理软件进行各条带灰度分析。以GAPDH为内参,各组P2X7受体与GAPDH条带灰度比值表示该P2X7受体相对含量。
2.8ELISA法测培养上清液中IL-1β含量 收集各组细胞上清液,离心后,依照试剂盒说明书操作,检测各组细胞培养上清液中IL-1β含量。
3统计学处理
结 果
1ATP对N9小胶质细胞活力的影响
XTT法检测细胞活力结果如图1所示,ATP对N9小胶质细胞活力的损伤具有一定的剂量依赖性,500 μmol/L ATP和1 mmol/L ATP对N9小胶质细胞活力的影响较小,作用24 h后,细胞活力分别为88.5%±5.5%和88.2%±8.4%。当ATP增加到2 mmol/L 时,对细胞活力的损伤程度明显增大,作用1 h、3 h、6 h、12 h和24 h后N9小胶质细胞活力分别为71.2%±5.9%、67.5%±5.1%、61.2%±5.4%、18.9%±6.6%和11.6%±1.1%。而当ATP浓度为5 mmol/L ATP和10 mmol/L ATP时,6 h后细胞活力分别为16.8%±1.6%和16.1%±2.0%,24 h后细胞活力仅为10.0%±0.3%和11.0%±0.8%。
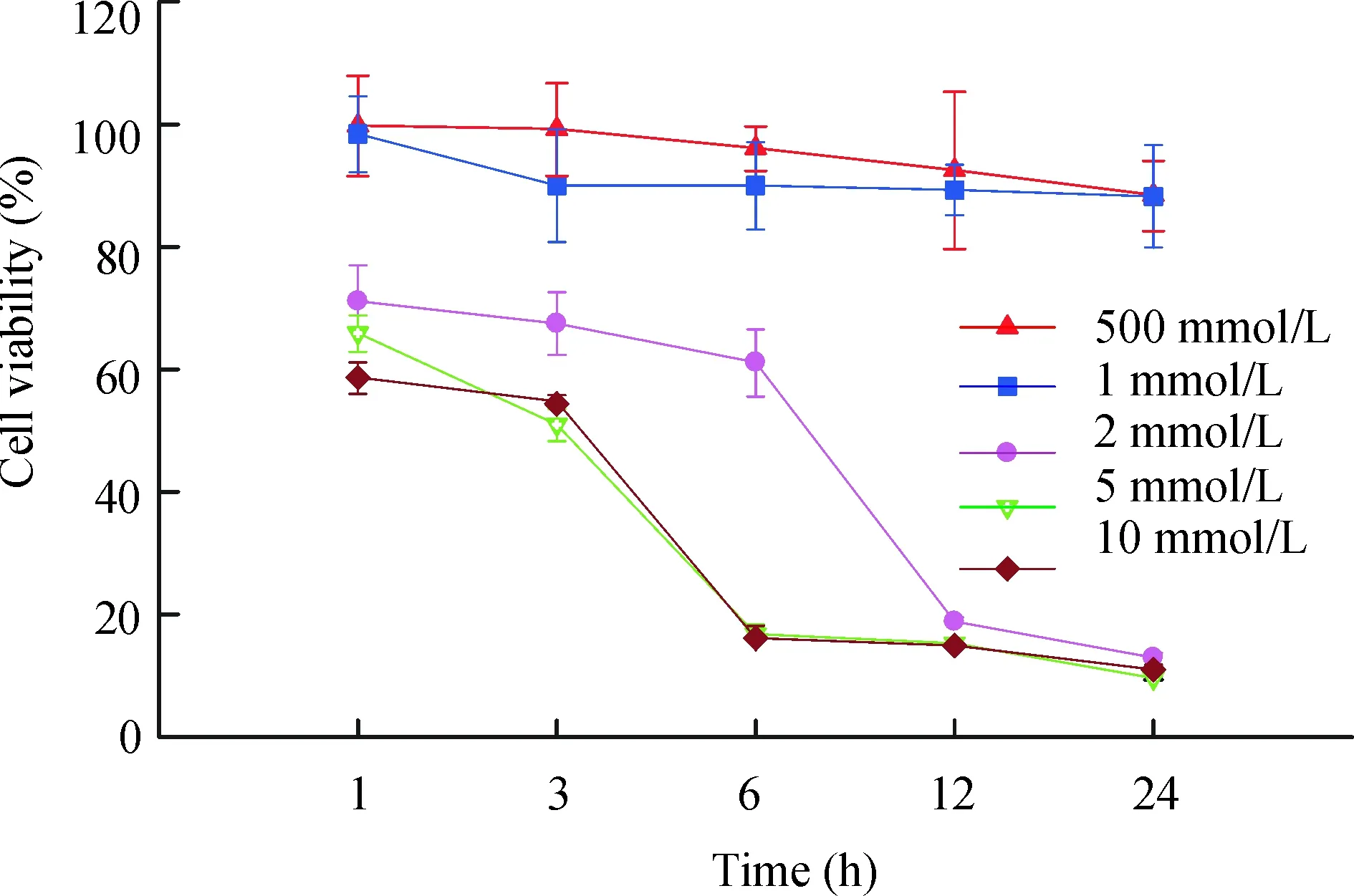
Figure 1. Effect of ATP on the cell viability of N9 microglia.
图1ATP对N9小胶质细胞活力的影响
2KN-62对ATP损伤的N9小胶质细胞活力的影响
如图2所示,0.1%DMSO作用24 h后,N9小胶质细胞活力为99.9%±5.2%,表明浓度≤0.1% DMSO对N9小胶质细胞活力无影响。2 mmol/L ATP作用1 h、3 h、6 h、12 h和24 h后N9小胶质细胞活力分别为71.2%±5.9%、67.5%±5.1%、61.2%±5.4%、18.9%±6.6%和11.6%±1.1%。而100 nmol/L KN-62干预后在2 mmol/L ATP作用的各时点细胞活力依次为66.5%±5.0%、64.9%±7.8%、58.9%±2.6%、19.1%±1.1%和16.3%±6.3%。而1 μmol/L KN-62干预后细胞活力在各时点分别为78.1%±5.9%、74.0%±7.6%、71.0%±4.7%、56.3%±3.7%和49.8%±2.8%。结果表明100 nmol/L KN-62对ATP损伤的N9小胶质细胞没有保护作用,而1 μmol/L KN-62干预则对ATP损伤的N9小胶质细胞具有较好的保护作用。
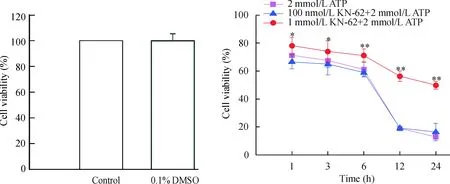
Figure 2. Effect of KN-62 on the cell viability of N9 microglia injured by ATP.*P<0.05,**P<0.01vs2 mmol/L ATP group.
图2KN-62对ATP损伤的N9小胶质细胞活力的影响
3ATP及KN-62干预对N9小胶质细胞形态的影响
如图3所示,对照组N9小胶质细胞多呈圆形,部分细胞有突起。随培养时间延长,细胞密度逐渐增大。ATP处理1h后,细胞突起消失,胞体回缩变圆,随着ATP处理时间延长,细胞皱缩程度增加,部分细胞开始破裂,处理12 h及24 h后,与对照组相比,细胞密度明显减少,大部分细胞严重皱缩死亡。而KN-62干预组细胞较为铺展,随时间延长,KN-62干预组细胞密度显著高于ATP组,形态也更接近于正常对照组。KN-62显现出拮抗ATP毒性损伤的保护作用。

Figure 3. Effects of ATP and KN-62 intervention on the morphology of N9 microglia.Scale bar=300 μm.
图3ATP和KN-62干预对N9小胶质细胞形态的影响
4ATP及KN-62干预对N9小胶质细胞凋亡的影响
如图4A和D所示,对照组没有细胞凋亡,而ATP作用3 h和6 h后,部分细胞发生凋亡,凋亡细胞比例分别为14.26%±0.72%和15.42%±0.93%,而KN-62可使ATP作用相应时点的凋亡细胞比例分别下降至1.69%±0.09%和1.31%±0.07%。此结果表明,2 mmol/L ATP可以诱导N9小胶质细胞凋亡,而阻断剂KN-62则可以通过减少细胞凋亡、拮抗ATP引起的细胞损伤而起到保护作用。


图4ATP和KN-62干预对N9小胶质细胞周期和凋亡的影响
5ATP及KN-62干预对N9小胶质细胞周期的影响
细胞周期检测结果如图4A、B和C所示,ATP作用3 h及6 h后,正常对照组、ATP组及KN-62干预组G0/G1期细胞比例无明显差异(P>0.05)。而ATP组S期细胞比例明显高于正常对照组(P<0.01),而G2/M期细胞比例却明显低于对照组(P<0.01)。KN-62干预组与ATP组相比,S期细胞比例明显降低(P<0.05),而G2/M期细胞比例则显著增多(P<0.05)。这些结果表明,ATP可使N9小胶质细胞阻滞于S期,抑制细胞分裂增殖,而KN-62干预后可使细胞周期向正常方向恢复。
6ATP及KN-62干预对N9小胶质细胞P2X7受体表达的影响
如图5所示,P2X7受体在N9小胶质细胞中广泛地表达于胞膜、胞质及核膜。ATP作用后,细胞胞体和胞核发生皱缩,细胞密度逐渐减少,但就单个细胞而言,P2X7受体表达部位并未见明显改变,由于细胞皱缩变小,所以荧光强度稍强于对照组(P<0.05)。而KN-62干预组的细胞形态和密度更接近于正常对照组,细胞上P2X7受体表达分布也未见明显改变。

图5ATP和KN-62干预对P2X7受体表达的影响
7ATP及KN-62干预对N9小胶质细胞P2X7受体蛋白水平的影响
如图6所示,P2X7受体蛋白在N9小胶质细胞中表达水平较高,ATP作用3 h后,正常对照组、2 mmol/L ATP组和KN-62+2 mmol/L ATP组3组之间P2X7受体蛋白的表达无明显差异(P>0.05),表明ATP作用及KN-62干预对P2X7受体蛋白的表达没有调节作用。
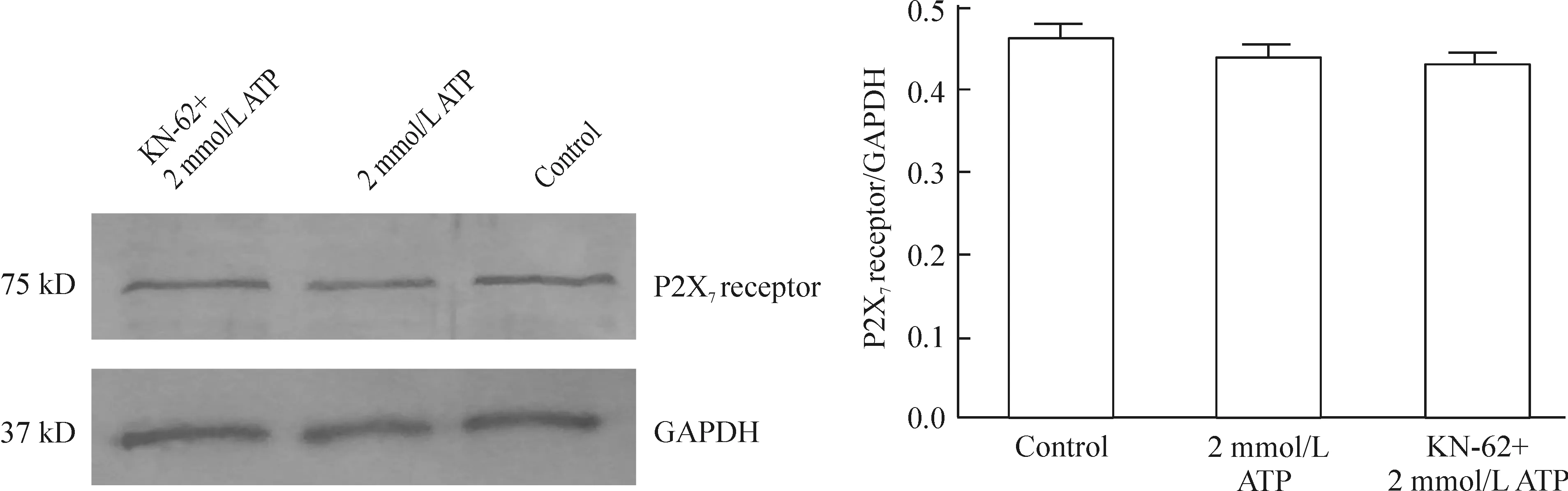

图6ATP和KN-62干预对N9小胶质细胞P2X7受体蛋白表达的影响
8ATP及KN-62干预对N9小胶质细胞培养液中IL-1β的影响
如表1所示,对照组细胞培养液中IL-1β水平低于试剂盒检测下限4 ng·L-1,1 h后低、高ATP及KN-62干预组细胞培养液中的IL-1β仍然维持在较低水平,且各组间无差异(P>0.05)。随着ATP处理时间延长,各组IL-1β水平未出现明显变化(P>0.05),表明ATP及KN-62处理不增加N9小胶质细胞释放IL-1β。
表1 ATP和KN-62干预对N9小胶质细胞培养液中IL-1β的影响
讨 论
1970年,Burnstock首先提出ATP在胞外可以充当信号分子的角色,并将其归类为非胆碱能、非肾上腺素能的神经递质。近些年来研究表明,ATP作为信号分子参与中枢及外周神经系统的多种生理和病理过程,如胚胎和出生后早期发育、突触可塑性、认知功能、神经性疼痛、痛觉过敏、神经炎症反应、损伤、缺血、神经退化性疾病、癫痫等[8-9]。目前关于ATP功能的研究正受到越来越多的关注。正常情况下,细胞外液中的ATP水平很低,但当中枢神经系统受到损伤(如缺血损伤)后,大量的ATP被释放到胞外,导致细胞外液中的ATP浓度迅速升高,最终引起周围神经元及其它细胞的继发性损伤和死亡。研究发现[10],ATP对恶性肿瘤细胞如肝癌细胞、白血病细胞、结肠移植癌、人肺巨细胞癌及表皮样癌等具有抑制增殖和诱发凋亡的作用。而本研究结果显示,低剂量的ATP对N9小胶质细胞活力的影响很小,当ATP浓度达到或大于2 mmol/L ATP时,短时间内就可以迅速引发N9小胶质细胞活力降低,细胞形态发生改变,随着ATP作用时间延长,细胞活力进一步降低,形态改变越来越明显,P2X7受体阻断剂KN-62可以拮抗ATP的损伤作用,ATP作用时间越长,KN-62的拮抗作用就越明显。此结果提示,P2X7受体介导了ATP对N9小胶质细胞的毒性损伤作用。
为进一步探讨ATP对N9小胶质细胞的损伤机制,采用流式细胞仪观察ATP及KN-62干预对N9小胶质细胞周期及凋亡的影响。一个完整的细胞周期包括:(1)G1期(gap1):DNA复制前期;(2)S期(synthesis phase):DNA复制期;(3)G2期(gap2):有丝分裂前期;(4)M期(mitosis):有丝分裂期。实验结果显示,ATP对G0/G1期没有影响,但是可将细胞阻滞在S期,不能进入G2/M期,从而抑制细胞的分裂和增殖,且ATP处理可引起细胞发生较大比例凋亡,而KN-62干预之后,细胞S期阻滞情况得以缓解,细胞凋亡比例明显下降。这些结果表明,ATP可通过P2X7受体的介导诱发细胞周期阻滞和凋亡从而造成对N9小胶质细胞的损伤。
ATP的信使作用由膜上的P2受体介导,P2受体分为离子门控型P2X受体和G蛋白偶联型P2Y受体两类。目前,P2X受体已克隆出7个亚型,分别为P2X1-7,其中P2X7受体因其生理功能较为独特且在多种生理病理过程中发挥重要作用,近些年来备受关注。体内研究发现,病理情况下如脑缺血缺氧损伤后,往往会发生小胶质细胞的大量激活增多和P2X7受体表达增加[5,11-12],陈翀等[13]用ATP刺激原代培养的小胶质细胞1 h后,免疫荧光检测发现1 mmol/LATP和2 mmol/L ATP刺激组P2X7受体表达量增加,而3 mmol/L ATP刺激组细胞发生明显凋亡,P2X7受体表达量减少。而本实验中免疫荧光和Western blotting检测结果显示,ATP及KN-62干预不改变P2X7受体在N9小胶质细胞上的分布。2 mmol/L ATP作用3 h后,对照组、ATP组及KN-62干预组之间P2X7受体蛋白表达水平无明显差异。这些结果提示在N9小胶质细胞,ATP的毒性损伤作用只是通过激活P2X7受体的功能而实现的,并不涉及其分布和表达的改变。本实验结果与前人体内实验结果不一致可能是由于在体内复杂的内环境中,脑缺血损伤后往往出现小胶质细胞的大量激活增多,而P2X7受体则在小胶质细胞上有广泛表达,因此,P2X7受体表达增多就不足为奇了。而体外实验中若培养条件有所不同,细胞对ATP的反应敏感度就有差异。且文献体外实验中只是根据细胞免疫荧光染色这一定性实验结果判断ATP上调了P2X7受体的表达,而本实验结合免疫荧光定性实验和Western blotting定量实验证明,ATP及KN-62干预不影响N9小胶质细胞P2X7受体的分布和表达水平,因此结果更为可靠。
近几年小胶质细胞成为研究焦点,作为中枢神经系统内的免疫细胞,它在急、慢性脑损伤疾病中的作用受到空前的关注。越来越多的研究发现小胶质细胞在脑损伤尤其是脑缺血缺氧损伤中发挥着非常重要的作用。缺血缺氧损伤后,小胶质细胞迅速激活,同时损伤的脑细胞释放出大量的ATP,激活小胶质细胞上的P2X7受体,随后小胶质细胞合成并释放大量的炎性介质,如IL-1β,引发炎性级联效应,最终导致其周围脑细胞的继发性损伤[5-6,14]。而Mingam等[15]体外研究发现采用ATP处理混合培养的原代胶质细胞不能增加胞外液中IL-1β的水平。本研究结果也显示,ATP虽然激活N9小胶质细胞上的P2X7受体通道,但是却没有促进IL-1β向胞外释放。这些体外实验结果提示,在小胶质细胞ATP虽可激活其上的P2X7受体,但却不能引发炎症反应。可见胞外ATP单独诱导的细胞损伤与小胶质细胞的炎症反应无关。看来体内胞外ATP诱导的继发性脑损伤的机制还远未阐明,研究ATP嘌呤信号通路或许能发现神经保护作用的新靶点,给脑损伤的防治带来希望。
综上所述,胞外高浓度的ATP(≥2 mmol/L)可迅速诱发N9小胶质细胞的细胞周期阻滞和凋亡从而损伤细胞,而P2X7受体特异性阻断剂KN-62则可通过减轻细胞周期阻滞和抑制细胞凋亡而拮抗ATP的损伤作用,表明ATP对N9小胶质细胞的损伤机制与P2X7受体介导的细胞周期阻滞和凋亡有关;且由于ATP及KN-62均不影响N9小胶质细胞炎症因子的释放,所以ATP对N9小胶质细胞的损伤机制不涉及小胶质细胞的炎症反应。至于ATP通过P2X7受体引发细胞周期阻滞和凋亡所涉及的具体通路和分子机制,除了P2X7受体外还有哪些受体及分子也参与了ATP对小胶质细胞的损伤过程,这些问题的阐明还需要更多的研究工作。
[1] Skaper SD, Debetto P, Giusti P. The P2X7purinergic receptor: from physiology to neurological disorders[J]. FASEB J, 2010, 24(2): 337-345.
[2] 秦 葵,朱忠宁,任雷鸣,等.外源性三磷酸腺苷对人食管癌Eca-109和肝癌SMMC-7721细胞增殖和周期的影响[J].基础医学与临床,2007,27(9):975-980.
[3] 陈燕花,陈振兵,康 皓,等.三磷酸腺苷对小鼠肌源性干细胞增殖和细胞周期的影响[J].中华实验外科杂志,2006, 23(2):170-172.
[4] Wen LT, Knowles AF. Extracellular ATP and adenosine induce cell apoptosis of human hepatoma Li-7A cells via the A3 adenosine receptor[J]. Br J Pharmacol, 2003, 140(6): 1009-1018.
[5] Skaper SD. Ion channels on microglia: therapeutic targets for neuroprotection[J]. CNS Neurol Disord Drug Targets, 2011, 10(1): 44-56.
[6] Friedle SA, Curet MA, Watters JJ, et al. Recent patents on novel P2X7receptor antagonists and their potential for reducing central nervous system inflammation[J]. Recent Pat CNS Drug Discov, 2010, 5(1): 35-45.
[7] Jonathan RW, Ines PK, Thomas M. Microglia in ischemic brain injury[J]. Future Neurol, 2010, 5(2): 227-246.
[8] Khakh BS, North RA. P2X receptors as cell-surface ATP sensors in health and disease[J]. Nature, 2006, 442(7102): 527-532.
[9] 宫庆娟,陈金生,黄乔东,等.小胶质细胞SFKs在ATP诱导的脊髓背角LTP中的作用[J].中国病理生理杂志, 2011,27(8):1563-1568.
[10]Völkl T, Ogilvie A, Neuhuber W, et al. Cell death induced by uridine 5’-triphosphate (UTP) in contrast to adenosine 5’-triphosphate (ATP) in human epidermoid carcinoma cells (A-431) [J]. Cell Physiol Biochem, 2008, 22(5-6):441-454.
[11]Franke H, Günther A, Grosche J, et al. P2X7receptor expression after ischemia in the cerebral cortex of rats[J].J Neuropathol Exp Neurol, 2004, 63(7):686-699.
[12]Yanagisawa D, Kitamura Y, Takata K, et al. Possible involvement of P2X7receptor activation in microglial neuroprotection against focal cerebral ischemia in rats[J]. Biol Pharm Bull, 2008, 31(6):1121-1130.
[13]陈 翀,吕 军,向正华,等.P2X7介导的小胶质细胞形态和受体表达的变化[J].解剖学杂志,2008,31(1):37-39.
[14]顾 云,庄 重.糖皮质激素对脑内炎症反应的促进作用[J].中国病理生理杂志,2011,27(10):2030-2034.
[15]Mingam R,De Smedt V,Amédée T, et al.Invitroandinvivoevidence for a role of the P2X7receptor in the release of IL-1β in the murine brain[J]. Brain Behav Immun, 2008, 22(2): 234-244.
InjuryeffectofadenosinetriphosphateonN9microglia
WANG Guo-hong1, GUO Zhi-yue1, YIN Ya-ling1, WEI Lin-yu1, LI Xin-juan1, LI Dong-liang1,2
(1DepartmentofPhysiologyandNeurobiology,XinxiangMedicalUniversity,Xinxiang453003,China;2StateKeyLaboratoryofMedicalNeurobiology,FudanUniversity,Shanghai200032,China.E-mail:xyldl8@126.com)
AIM: To investigate the injury effect of adenosine triphosphate (ATP) on N9 microglia.METHODSN9 microglia in logarithmic growth phase was randomly divided into 3 groups. In control group, the cells were cultured without ATP treatment. In ATP group, the cells were treatment with ATP after cultured for 24 h. In KN-62 intervention group, after pretreatment with KN-62 for 30 min, ATP was added in the cells. The cell viability was assessed by XTT assay. Cellular morphological changes were observed under phase-contrast microscope. The cell cycle and apoptosis were detected by flow cytometry. The expression of P2X7receptor was examined by immunofluorescence staining. The protein levels of P2X7receptor were measured by Western blotting. The concentration of IL-1β in the culture supernatant was detected by ELISA.RESULTSATP at dose of 500 μmol/L and 1 mmol/L only caused small damage to the cell viability of N9 microglia. The cell viability was 88.5%±5.5% and 88.2%±8.4% after treated with ATP for 24 h,respectively. The cell viability dropped rapidly and cell shrinkage occurred when the concentration of ATP increased to 2 mmol/L or higher. With the extension of experiment time, the cell viability and cell density decreased further and cell shrinkage was getting worse. KN-62 intervention improved the viability of N9 microglia injured by ATP. The morphology and density of N9 microglia in KN-62 intervention group were much better than those in ATP group. ATP arrested N9 microglia at S phase and increased cell apoptosis significantly (P<0.01vscontrol group). KN-62 intervention obviously relieved the cell cycle arrest and decreased the cell apoptosis caused by ATP (P<0.01). ATP and KN-62 intervention had no effect on the distribution of P2X7receptor. The protein levels of P2X7receptor had no significant difference among the 3 groups (P>0.05). ATP and KN-62 intervention had no effect on the release of IL-1β.CONCLUSIONHigh dose of ATP damages N9 microglia and its mechanism may be related to cell cycle arrest and apoptosis mediated by P2X7receptor but not to inflammatory response caused by microglia.
Adenosine triphosphate; Microglia; Receptors,purinergic P2X7; KN-62
R363
A
10.3969/j.issn.1000-4718.2012.09.011
