条纹锯对高温胁迫及胁迫后恢复方式适应的初步研究
2012-01-12曹金凤王成刚王志军
刘 超, 曹金凤, 马 甡, 王成刚, 王志军
(1. 中国海洋大学 教育部海水养殖重点实验室, 山东 青岛 266003; 2. 山东科合海洋高技术有限公司, 山东乳山 264513)
刘 超1, 曹金凤1, 马 甡1, 王成刚2, 王志军2
(1. 中国海洋大学 教育部海水养殖重点实验室, 山东 青岛 266003; 2. 山东科合海洋高技术有限公司, 山东乳山 264513)
研究了质量为148.88 g±17.29 g和16.32 g±2.23g两种规格的条纹锯(Centropristis striata)鱼种对高温的耐受性及高温胁迫后恢复方式的适应。结果表明: 大规格鱼种的高温半致死温度为 26.80℃,最适温度上限为20.38℃。小规格鱼种的高温半致死温度为29.26℃, 最适温度上限为25.11℃。依据实验鱼种耗氧率的恢复情况可以断定: 在暴露于高温半致死温度48 h或72 h条件下, 直接恢复方式略优于梯度恢复方式; 而暴露24 h条件下, 两种恢复方式的作用无明显差异。高温半致死温度暴露下, 要避免条纹锯达到半数死亡则积温不能超过(1 064.67±72.01)℃·h。
条纹锯(Centropristis striata); 高温半致死温度; 最适温度;恢复方式; 半致死积温
积温指某一时段内逐日平均温度累加之和。关于积温的研究已经有很多报道[10-12], 但鲜见关于鱼种积温与存活关系的报道。根据前期预备实验的观察, 实验中所用条纹锯在其高温半致死温度或更高温度下存活时间仅有数小时甚至数分钟, 为研究方便本实验使用“℃·h”作为积温单位来研究小规格鱼种在半致死温度暴露下以及降温过程中的半致死积温以及最佳存活积温(受试鱼种死亡 10%的积温)。测定条纹锯在高温半致死温度暴露下的半致死积温以及最佳存活积温, 为条纹锯经高温半致死温度胁迫后能否恢复到正常代谢水平提供一种判断方法。
1 材料与方法
1.1 实验材料
1.2 实验方法
1.2.1 条纹锯高温半致死温度及最适温度上限
大规格鱼种实验温度设置为 18、22、26、30、34 ℃。每个温度处理设置3个平行。同时在室温条件下(16 ℃)设置一个对照, 同样设置 3个平行。实验在 20 L的方形玻璃槽中进行, 持续曝气, 待实验用海水达到相应温度并稳定12 h后, 每个平行放养4尾大规格鱼种, 持续曝气, 投饵时间、投饵量以及换水量、换水频率均与暂养时相同, 每隔1 h观察一次,连续观察 96 h, 记录各温度处理中鱼种的死亡数量及时间。做出温度-存活率曲线, 用内插法计算出其高温半致死温度和最适温度上限。
小规格鱼种的实验温度设置为22、24、26、28、30 ℃, 以室温(16 ℃)作为对照, 各处理亦设置 3个平行, 每个平行组放养5尾鱼种。实验过程以及实验结果的处理同于大规格鱼种。
1.2.2 小规格鱼种高温暴露后恢复方式的比较及半致死积温
在容积为 40 L的大桶内, 将海水温度调节到1.2.1计算得到的小规格鱼种的高温半致死温度, 连续曝气, 稳定12 h后, 放养115尾小规格鱼种, 剔除在2 h内死亡的个体。分别在暴露24、48、72 h时, 各随机取出30尾(其中存活个体总数大于24尾)。其中15尾直接放入18 ℃的海水中, 72 h后随机取3尾非死亡个体应用间接碘量法测定其绝对耗氧率; 另外15尾放入 26 ℃海水中, 暴露 24 h后, 再将其转入22 ℃海水中暴露24 h, 最终转入18 ℃海水中, 24 h后从中任意取 3尾非死亡个体测定其绝对耗氧率。依据其绝对耗氧率与对照组绝对耗氧率的差别来判定恢复方式对实验鱼种高温胁迫后恢复的影响。计数小规格鱼种在恢复过程中的死亡数量及时间。由实验所得数据做出存活时间—温度曲线, 计算出恢复过程中条纹锯鮨的半致死积温。
同时在容积为10 L的水桶中, 装入2/3体积的海水, 温度调节至1.2.1计算得到的高温半致死温度,持续曝气, 温度稳定12 h后, 放入30尾小规格鱼种,剔除在2 h内死亡的个体和部分成活个体, 水桶中最后仅保留15尾成活个体。每隔1 h观察一次, 计数鱼种的死亡数量及时间。由实验所得数据做出存活时间—温度曲线, 计算出持续暴露在半致死温度下条纹锯的半致死积温。
1.3 数据处理
进行积温计算时采用如下公式:K=N×(T-C)
其中:K为积温值, 单位为℃·h;N为暴露时间,单位为小时(h);T为暴露温度, 单位℃;C为驯养温度, 单位为℃。
2 实验结果
2.1 条纹锯高温半致死温度及最适温度上限
两种规格鱼种的半致死时间以及存活率见表1、表2。

表1 大规格鱼种的半致死时间以及96 h存活率Tab. 1 The semi-lethal time and 96-h survival rate of larger-sized fingerling
利用表1和表2的数据应用内插法计算得到如下实验结果(表3)。
2.2 小规格鱼种高温暴露后恢复方式的比较

表2 小规格鱼种的半致死时间以及96 h存活率Tab. 2 The semi-lethal time and 96-h survival rate of smaller-sized fingerling

表3 大规格鱼种和小规格鱼种最适温度上限和高温半致死温度Tab. 3 Semi-lethal temperature and upper limit of optimum temperature of two kinds of fingerlings
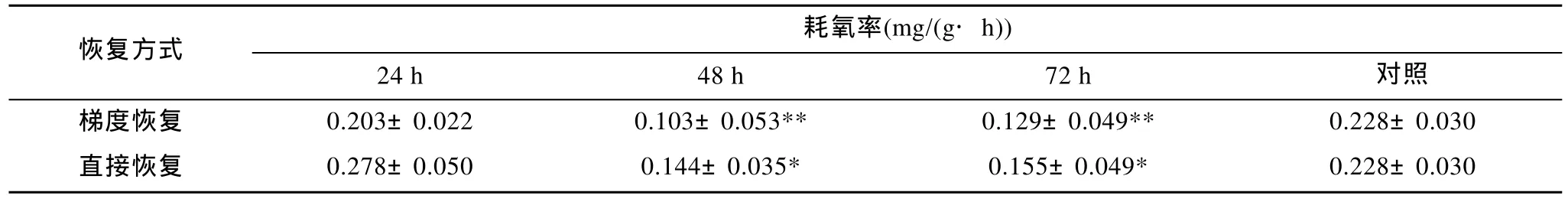
表4 不同暴露时间下恢复至18 ℃时鱼种的耗氧率Tab. 4 The oxygen consumption rate at 18 ℃ after recovery
从耗氧率的处理结果可以看出对于在高温半致死温度下暴露24 h的鱼种, 两种恢复方式均能使其恢复到正常的耗氧范围。而对于在高温半致死温度下暴露48 h和72 h的鱼种, 虽然经两种方式恢复至18 ℃24 h后均未恢复到正常的耗氧范围, 但是从差异的显著程度可以判断: 直接恢复方式要略优于梯度恢复方式。
2.3 条纹锯的半致死积温

图1 小规格鱼种的积温示意图Fig. 1 Accumulated temperature of smaller-sized fingerling
持续暴露于高温半致死温度下的鱼种, 由实验观察得到最佳存活时间时间(10%死亡)为54 h(M点,死亡2尾), 半致死时间为82 h(C点, 死亡8尾), 积温设为K1; 直接恢复方式的条纹锯, 半致死时间为100 h, 其中29℃下暴露了72 h(B点, 死亡5尾),18℃暴露了28 h(F-H,死亡3尾, 共计死亡8尾), 积温设为K2; 梯度恢复方式时, 半致死时间为92 h, 其中29℃暴露72 h(死亡7尾), 26℃暴露20 h(D-E,死亡1尾, 共计死亡8尾), 积温设为K3。
计算积温时, 取C=16 ℃应用积温公式数据处理如表5所示。
由此, 可以初步断定在半致死温度下至少暴露72 h的条纹锯, 达到半数死亡所经暴露积温为:(1064.67±72.01)℃·h。
从图1可以看出其最佳存活时间为54 h, 其在高温半致死温度暴露下的最佳存活积温的数值同四边形AMNO面积的数值相同, 故其高温半致死温度暴露下的最佳存活积温为702 ℃·h。

表5 小规格鱼种的半致死积温Tab. 5 Semi-lethal accumulated temperature of smallersized fingerling
3 讨论
3.1 条纹锯高温半致死温度及最适温度上限
在鱼类生存的环境中, 温度具有多方面的生态作用, 直接或间接影响鱼类的摄食、生长和存活等方面。Buentello等[13]研究报道, 22.8 ℃条件下斑点叉尾(Ictalurus punctatus)的摄食量和饲料转化效率均处于最高值。龙华[14]报道, 在一定范围内, 较高温度鱼类生长快, 较低温度鱼类生长慢。超出一定范围的高温条件下, 由于生物活性物质失活, 导致鱼类停止生长或死亡。
据雷霁霖等[15]报道, 条纹锯适宜生存温度为5~31℃, 最适水温为17~25℃。本实验结果与其差别较大, 其原因可能是本实验用的鱼种为刚刚经过越冬期的鱼种, 对高温的适应能力还未达到最大限度, 实验所得结果较低。由实验结果可以看出, 小规格鱼种的最适温度上限、高温半致死温度均高于大规格鱼种。所以, 小规格鱼种对高温的耐受力要优于大规格鱼种。
姜礼燔[16]认为鱼类热致死的机制主要有以下几点: (1)神经系统的破坏; (2)酶系统和呼吸中心损害;(3)血液系统破坏; (4)组织结构的破坏。此外, 闫茂仓[17]、谢刚[18]分别研究了鱼体质量对条石鲷(Oplegnathus fasciatus)幼鱼、卷口鱼(Ptychidio jordani)耗氧率的影响, 发现其耗氧率均随鱼体质量增加而降低。据黄玉瑶等[19]的研究可以推测小规格鱼种对高温的耐受力要优于大规格鱼种原因为小规格鱼种处于生长发育过程中, 代谢水平高, 可对高温产生的诸如神经系统破坏等危害进行及时修复, 使其在高温条件下能更长时间地存活。
3.2 条纹锯对高温的适应
3.3 条纹锯的半致死积温及最适积温
[1]徐涛, 马甡, 王志军. 低温对条纹锯生长及能量收支的影响[J]. 海洋科学, 2007, 31(4): 9-12.
[2]阳刚, 刘超, 马甡, 等. 条纹锯对盐度胁迫的耐受力及淡水胁迫恢复的初步研究[J]. 海洋湖沼通报,2011, 2: 73-78.
[3]曹金凤, 阳刚, 马甡, 等. 不同温度条件下条纹锯耗氧率与窒息点的初步研究[J]. 齐鲁渔业, 2009,27(12): 1-3.
[4]Charles F C, Randal L W. Comparison of four commercial diets and three feeding rates for black sea bass,Centropristisstriatafingerlings[J]. Journal of Applied Aquaculture, 2004, 16(3/4): 131-146.
[5]Rachel A H. David L B, Terence M B. The effect of photoperiod on the reproduction of black sea bassCentropristisstriata[J]. Acquculture, 2003, 218: 651-669.
[6]Kenneth W, Able L, Stanton H.Movement of juvenile black sea bassCentropristisstriatain a southern New Jersey Estuary[J]. Journal of Experimental Biology and Ecology, 1997, 213: 153-167.
[7]Kevin R S, Theodore I J S. Development of nursery systems for black sea bassCentropristisstriata[J]. Journal of the World Aquaculture Society, 2003, 34(3): 359-367.
[8]王波, 朱明远, 毛兴华.养殖新品种——美洲黑石斑鱼[J].河北渔业, 2003, 5: 26-27.
[9]卢继武, 刘敏, 李豪.条纹锯鮨鱼的生物学特性与养殖潜力[J]. 渔业现代化, 2006, 6: 21-22.
[10]关忠志, 刘吉明, 李东占. 香鱼性腺发育与积温关系的初步研究[J]. 水产学杂志, 2008, 21(2): 33-36.
[11]王云果, 李孟楼, 高智辉. 花斑皮蠹发育起点温度和有效积温研究[J]. 西北农业学报, 2008, 17(4): 208-210.
[12]陈禄仕. 利用积温和昆虫发育历期推测死亡时间的研究[J]. 中国法警学杂志, 2007, 22(4): 236-237.
[13]J. Alejandro B, Delbert M G Lii, William H N. Effects of water temperature and dissolved oxygen on daily feed consumption, feed utilization and growth of channel cat fish(Ictalurus punctatus)[J]. Aquaculture, 2000,182: 339-352.
[14]龙华. 温度对鱼类生存的影响[J]. 渔业现代化, 2005,2: 21-22.
[15]雷霁霖, 卢继武.条纹锯鮨鱼的品种优势和养殖前景[J]. 海洋水产研究, 2007, 28(5): 111-115.
[16]姜礼燔. 热冲击对鱼类影响的研究[J]. 中国水产科学, 2007, 7(2): 77-81.
[17]闫茂仓, 单乐州, 谢起浪. 温度、盐度及体重对条石鲷幼鱼耗氧率和排氨率的影响[J]. 海洋科学进展,2008, 26(4): 486-495.
[18]谢刚, 许淑英, 祁宝嵛, 等. 卷口鱼耗氧规律的研究[J]. 大连水产学院学报, 2002, 17(2): 89-94.
[19]黄玉瑶.鲤鱼、白鲢、鳊鱼的鱼苗、鱼种耗氧量的研究[J]. 动物学报, 1975, 21(1): 78-88.
[20]陈品建, 王重刚, 陆皓, 等. 真鲷幼鱼消化酶活性与温度的关系[J]. 厦门大学学报(自然科学版), 1998,37(6): 931-935.
The adaptability of black sea bass (Centropristis striata) to semi-lethal temperature and recovery mode from exposure to semi-lethal temperature
LIU Chao1, CAO Jin-feng1, MA Shen1, WANG Cheng-gang2, Wang Zhi-jun2
(1.Key Laboratory of Mariculture, Ministry of Education, Ocean University of China, Qingdao, 266003, China;2.Shandong Kehe Marine Hi-technology CO., Ltd, Rushan, 264513, China)
Feb.,28, 2011
Centropristis striata; semi-lethal temperature ; optimum temperature; recovery mode ; semi-lethal accumulated temperature
In this paper, two kinds ofCentropristis striata(148.88 g±17.29 g and 16.32 g±2.23 g) were used to investigate the adaptability of black sea bass to semi-lethal temperature and the recovery mode from exposure to semi-lethal temperature. By the interpolation method, we got that the larger-sized fingerling’s semi-lethal temperature and the upper limit of optimum temperature were 26.80 ℃ and 20.38 ℃, respectively. And the smaller-sized fingerling’s semi-lethal temperature and the upper limit of optimum temperature were 29.26 ℃ and 25.11 ℃, respectively. By measuring the oxygen consumption rate (OCR), we found the direct mode was more propitious for fingerling’s recovery than gradient mode after continuous exposure to semi-lethal temperature for 48 h or 72 h. But both of the modes could make them recover successfully after being exposed for 24 h. In addition, the semi-lethal accumulated temperature should be less than 1064.67 ℃·h±72.01 ℃·h at semi-lethal temperature in order to exempt semi-lethal rate.
Q142.9 文献标识码: A 文章编号: 1000-3096(2012)09-0054-05
2011-02-28;
2011-09-29
国家科技成果转化资金项目(2007GB2C600501)
刘超(1984-), 男, 山东潍坊人, 硕士研究生, 主要从事养殖生态研究, 电话: 0532-82032195, E-mail: liuchaofighting@163.com.; 马甡, 通信作者, 电话: 0532-82032041, E-mail: mashen@ouc.edu.cn
谭雪静)
