Visual modeling reveals cryptic aspect in egg mimicry of Himalayan Cuckoo (Cuculus saturatus) on its host Blyth’s Leaf Warbler (Phylloscopus reguloides)
2011-12-25YANGCanChaoCAIYanLIANGWei
YANG Can-Chao, CAI Yan LIANG Wei*
(1. College of Life Sciences, Hainan Normal University, Haikou 571158, China; 2. State Key Laboratory of Biocontrol, School of Life Sciences, Sun Yat-sen University, Guangzhou 510275, China)
Visual modeling reveals cryptic aspect in egg mimicry of Himalayan Cuckoo (Cuculus saturatus) on its host Blyth’s Leaf Warbler (Phylloscopus reguloides)
YANG Can-Chao1,2, CAI Yan1, LIANG Wei1,*
(1. College of Life Sciences, Hainan Normal University, Haikou 571158, China; 2. State Key Laboratory of Biocontrol, School of Life Sciences, Sun Yat-sen University, Guangzhou 510275, China)
Brood parasitism and egg mimicry of Himalayan Cuckoo (Cuculus saturatus) on its host Blyth’s Leaf Warbler (Phylloscopus reguloides) were studied in south-western China from April to July 2009. The cuckoo laid a white egg with fine brown markings on the blunt end. The eggs were conspicuously bigger than the host’s own, with 2.06 g in mass and 1.91 cm3in volume. Visual modeling showed that the cuckoo eggs, which from the human eye appeared to mimic the host eggs to a great extent, were completely different from the host eggs in both hue and chroma. The characters of the Himalayan Cuckoo nestling, reported for the first time, included two triangular and black patches on its gape, which appeared from four days old and became darker with age and growth. While this character also exists in nestlings of Oriental Cuckoo (C. optatus), it has not been found for otherCuculusspecies. Our results reveal cryptic aspects in the cuckoo-host egg color matching, which are not visible to the naked human eye, and indicate that high mimetic cuckoo eggs rejected by hosts, as determined by human observers in previous studies, might not be mimetic as birds see them.
Egg color; Egg matching; Visual modeling; Spectrophotometer;Cuculus saturatus;Phylloscopus reguloides; Robinson projection
Avian brood parasites exploit parental care of their hosts and generally reduce their reproductive success. Under this selective pressure, hosts should evolve defenses to reduce the impact of parasitism, while parasites should evolve strategies to counter such defenses (Davies, 2000). Most of these co-evolutionarybattles focus on the egg stage, whereby cuckoos lay mimetic eggs to cheat the hosts who, in turn, recognize and reject the cuckoo eggs. This interaction even provokes evolution of egg color polymorphism in both cuckoo and host (Yang et al, 2010b). Based on our observations, the Himalayan Cuckoo (Cuculus saturatus) parasitizes the Blyth’s Leaf Warbler’s (Phylloscopus reguloides) nest and lays mimetic eggs. However, avian and human visual systems differ in several respects. For instance, many bird species have ultraviolet-sensitive (UVS) photoreceptors as well as oil droplets that are absent in the human eye (Goldsmith et al, 1984; Vorobyev et al, 1998). Ultraviolet-visible reflectance spectrophotometry has revealed aspects of cuckoo-host egg matching that are not noticed using human vision (Cherry & Bennett, 2001). Previous research from Australia has also shown that the color of Pallid Cuckoo (C. pallidus) eggs differs between four host species, but closely mimicked the eggs of the host they parasitized using reflectance spectrophotometry (Starling et al, 2006). These results reveal that some cuckoos appear to have host-specific egg types not visually detectable by human observation, and indicate that cuckoo gentes could be more common than previously thought. In this study, we examined the egg morph and egg size of the Himalayan Cuckoo, and established visual modeling to evaluate its egg mimicry on Blyth’s Leaf Warbler based on bird vision. Furthermore, we report on the nestling characteristics of the Himalayan Cuckoo, which have not been described before.
1 Materials and Methods
1.1 Study area
The study was performed in Kuankuoshui Nature Reserve, Guizhou province, south-western China (28° 10' N, 107° 10' E) from April to July 2009. The study area is situated in a subtropical moist broadleaf and mixed forest with an altitude of approximately 1 500 m, interspersed with abandoned tea plantations, bamboo, shrubby areas, and open fields used as cattle pastures. The annual average temperature is 13.6 °C and the average annual precipitation is 1 210 mm (also see Yang et al, 2010 b).
1.2 Field procedures
One clutch with one cuckoo and three host eggs was found and photographed on a Kodak Grey Card in a standardized manner with a Canon EOS camera. Nest habitat of the parasitized nest and the cuckoo nestling were also photographed. Egg size, length of tarsus, beak and wing of the cuckoo nestling were measured by using a digital caliper (111-101B, 150 mm, Guilin Guanglu Measuring Instrument Co. Ltd., Guilin, China). Egg mass and nestling body mass were measured by a pocket scale (DPS-015, 100 g, Viabetter Electronic Scales Limited., Shenzhen, China). Egg volume was calculated according to the formula published by Hoyt (1979). 1.3 Egg color quantification and visual modeling
We obtained spectral reflectance by spectrophotometer Avantes-2048 (Avantes, Netherlands), which provided an objective description of egg color. Six measurements per egg (two at the blunt, two at the middle, and two at the sharp parts of the egg) were surveyed and summarized as the mean of each egg (Yang et al, 2009). Very few spots were found on the cuckoo egg, thus they were not included in spectra measurement. To account for the differential stimulation of the four avian cone types, the spectra were mapped onto Goldsmith’s tetrahedral color space to analysis color patterns as processed by tetrachromatic visual system (Goldsmith, 1990) using the tetracolorspace program (Stoddard & Prum, 2008). Average spectral sensitivity curves for UVS-type retinas were used (Endler & Mielke, 2005). Essentially, each spectrum is represented by a point in a tetrahedron, in which the vertices correspond to exclusive stimulation of the ultraviolet (UV), blue (B), green (G) and red (R)-sensitive cones in the avian eye. Each color point can be described by its spherical coordinates (θ, φ, r), where angles θ and φ represent the horizontal (RGB) and vertical (UV) components of hue, respectively, and r is the length of the color vector in chroma or color saturation. To visualize hue distributions independently of chroma, we mapped colors onto a unit sphere centered on the achromatic origin using Robinson projection, where θ [−π; π] corresponds to longitude, and φ [−π/2; π/2] to latitude (Endler et al, 2005). As a measure of achromatic brightness, we calculated normalized brilliance following Stoddard & Prum (2008). Tetracolorspace program was performed in Matlab7 for Windows.
2 Results
The parasitized nest was built in a cavity on a soil slope and was about 2 m above the ground (Fig.1). The nest cup was made of moss, with some dry grass leaves around the entrance rim, and was 3.8 cm wide and 7.2 cm in deep. The Blyth’s Leaf Warbler laid immaculate white eggs, which were obviously smaller than the Himalayan Cuckoo egg (Tab. 1). Differential percentage of host eggs to cuckoo egg revealed that the cuckoo egg was slimmer than the host eggs in shape. The cuckoo egg was white in color with few brown spots on the blunt end. From the human eye, the cuckoo egg color appeared mimetic to the host eggs (Fig.2a). However, visual modeling showed that the cuckoo egg was completely unlike the host eggs in both hue and chroma (Fig.3). Distinct distribution on the vertical hue space also indicated that the ultraviolet component of the hue was also totally different between the cuckoo and host eggs (Fig.4).
The Himalayan Cuckoo nestling was naked at hatching (Fig.2b), weighed 1.78 g and measured 5.27 mm in tarsus length. Initially skin was flesh-color but became blackish after three days. Its gape was orange changing to orange-red and gape flanges yellow to orange. Two triangular patches on the palate adjoin the cutting edges of the upper mandible appeared from four days old and became darker with age and growth (Fig.2c).

Fig.1 Parasitized nest and its habitat of Blyth’s Leaf Warbler

Fig. 2 Egg and nestling of Himalayan Cuckoo
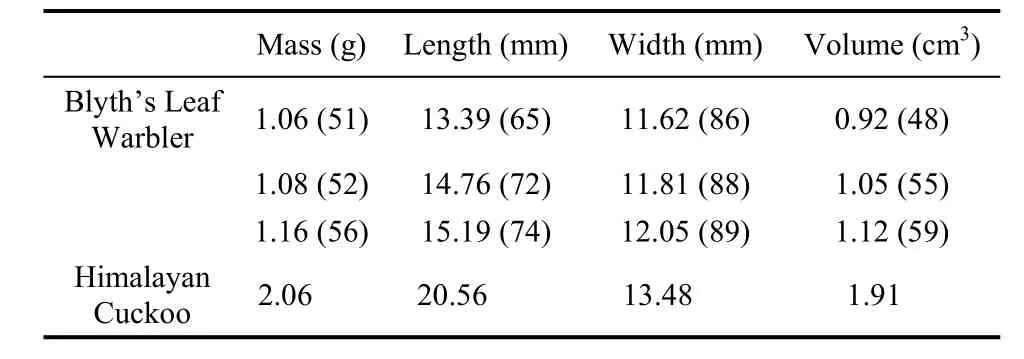
Tab. 1 Egg size and mass of Blyth’s Leaf Warbler and Himalayan Cuckoo in the observed nest. Shown are values with differential percentage of host eggs to cuckoo egg in brackets

Fig. 3 Robinson projection of egg color hue of the Himalayan Cuckoo and Blyth’s Leaf Warbler
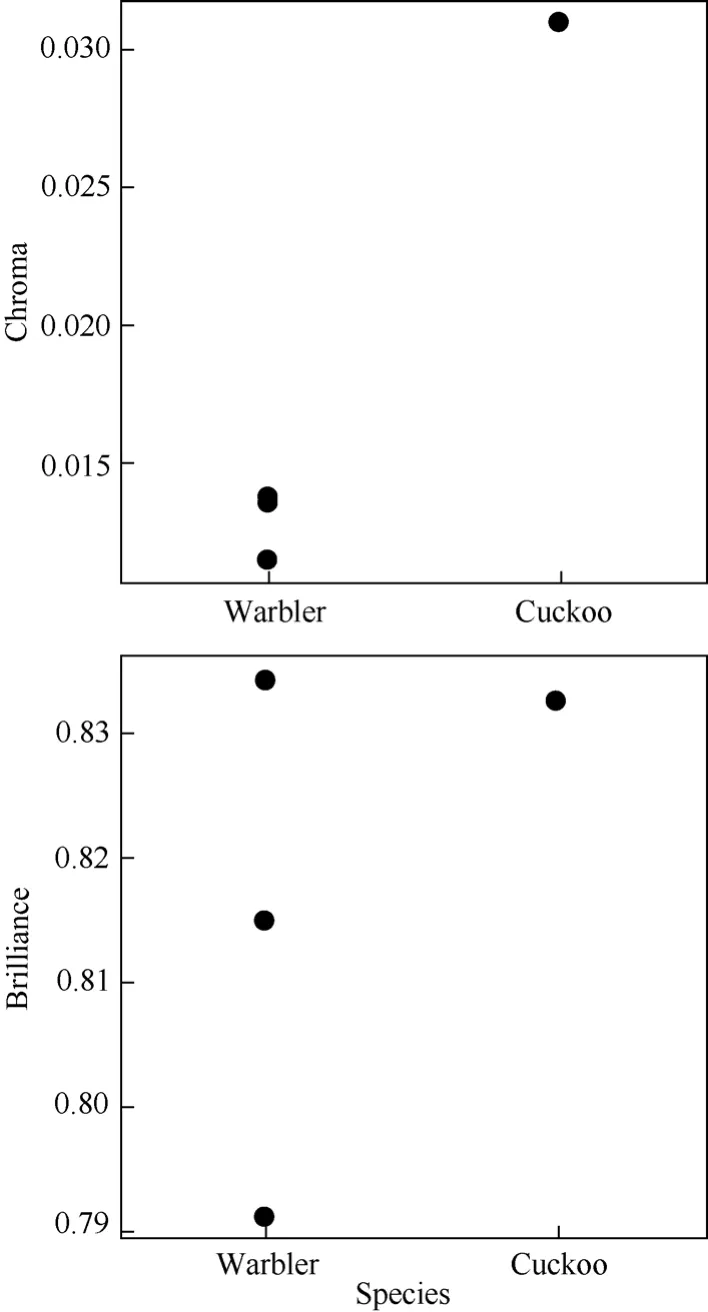
Fig. 4 Chroma and normalized brilliance as a measure of achromatic brightness of the Himalayan Cuckoo and Blyth’s Leaf Warbler eggs
3 Discussion
The major hosts of Himalayan Cuckoo arePhylloscopusspecies. In Russia, they parasitize Chiffchaff (P. collybita), Eastern Crowned-warbler (P. coronatus), Pale-legged Leaf-Warbler (P. tellennipes), Tree Pipit (Anthus hodgsoni) and Black-throated Accentor (Prunella atrogularis) (Irisov, 1967; Cramp, 1985; Knystautas & Sibnev, 1987). In Japan, cuckoos are reared by the Bush Warbler (Cettia diphone), Narcisus Flycatcher (Ficedula narcissina), and Short-tailed Bush Warbler (C. squameiceps) (Higuchi, 1998). In China, the Himalayan Cuckoo parasitizes Yellow-bellied Prinia (Prinia flaviventris) (La Touche, 1934; Zhang, 1980) and Rufous-capped Babbler (Stachyris ruficeps) (Lin, 2008). Eggs are reddish brown and mimetic to prina and babbler in Taiwan (La Touche, 1934; Lin, 2008).
Studies using ultraviolet-visible reflectance spectrophotometry found that the Red-chested Cuckoo (C. solitarius) eggs were mimetic to the host eggs, although they looked non-mimetic to the human eye (Cherry & Bennett, 2001). These results had the potential to explain the longstanding paradox that some host species accept cuckoo eggs that appear non-mimetic to the human eye. Conversely, between Himalayan Cuckoo and Blyth’s Leaf Warbler we found that the mimetic cuckoo egg based on human vision was non-mimetic based on bird vision. Our results reveal a cryptic aspect in cuckoo-host egg color-matching not detected from the human eye, and indicate that high mimetic cuckoo eggs rejected by hosts might not be mimetic as birds see them. Another spectrophotometry study on egg color-matching in the Lesser Cuckoo (C. poliocephalus) and its Brownish-flanked Bush Warbler (C. fortipes) host shows that cuckoo egg mimicry is consistent from both the human and bird eye (Yang et al, 2010a). The main defenses used by hosts against parasitism include rejecting odd-looking eggs from their nests (Kilner 2006). Although the Himalayan Cuckoo egg was non-mimetic to Leaf Warbler eggs, the host had accepted the cuckoo egg. This implied that the Blyth’s Leaf Warbler might be an acceptor or intermediate rejecter. To further understand this issue, experimental parasitism should be conducted to test egg recognition ability of the Leaf Warbler. The black patches on the gape of the Himalayan Cuckoo nestling have also been found on the nestlings of Oriental Cuckoo (C. optatus), but not in otherCuculuscuckoo species (Higuchi & Sato, 1984; Higuchi, 1998; Yoshino, 1999; Tojo et al, 2002; Wang et al, 2004; Payne, 2005).
Acknowledgements:We thank the Forestry Department of Guizhou Province and Kuankuoshui National Nature Reserves for support and permission to carry out this study. We are grateful to two anonymous referees for their comments on the manuscript.
Cherry MI, Bennett ATD. 2001. Egg colour matching in an African cuckoo, as revealed by ultraviolet-visible reflectance spectrophotometry [J].Proc R Soc Lond B, 268: 565-571.
Cramp S. 1985. The Birds of the Western Palearctic [M]. Oxford: Oxford University Press.
Davies NB. 2000. Cuckoos, Cowbirds and Other Chest [M]. London: Poyser.
Endler JA, Mielke PW. 2005. Comparing entire colour patterns as birds see them [J].Biol J Linn Soc, 86: 405-431.
Endler JA, Wescott DA, Madden JR, Tobson T. 2005. Animal visual systems and the evolution of color patterns: sensory processing illuminates signal evolution [J].Evolution, 59: 1795-1818.
Goldsmith TH. 1990. Optimization, constraint, and history in the evolution of eyes [J].Q Rev Biol, 65: 281-322.
Goldsmith TH, Collins JS, Licht S. 1984. The cone oil droplets of avian retinas [J].Vision Res, 24: 1661-1671.
Higuchi H. 1998. Host Use and Egg Color of Japanese Cuckoos [M]// Rothstein SI, Robinson SK. Parasitic Birds and Their Hosts. Oxford: Oxford University Press, 80-93.
Higuchi H, Sato S. 1984. An example of character release in host selection and egg colour of cuckoosCuculussp.In Japan [J].Ibis, 126: 398-404.
Hoyt DF. 1979. Practical methods of estimating volume and fresh weight of bird eggs [J].Auk, 96: 73-77.
Irisov EI. 1967. The Himalayan Cuckoo in the south-eastern Altai [J].Ornitologiya, 8: 335-336.
Kilner RM. 2006. The evolution of egg colour and patterning in birds [J].Biol Rev, 81: 383-406.
Knystautas AJV, Sibnev JB. 1987. Die Vogelwelt Ussuriens. A. Ziemsen [M]. Lutherstadt: Verlag.
La Touche JDD. 1934. A Handbook of the Birds of Eastern China [M]. London: Taylor and Francis.
Lin RX. 2008. Observation of brood parasitism on Rufous-cappd Babbler by Himalayan Cuckoo [J].Nat Conserv Quarterly, 64: 58-62.
Payne RB.2005. The Cuckoos [M]. Oxford: Oxford University Press.
Starling M, Heinsohn R, Cockburn A, Langmore NE. 2006. Cryptic gentes revealed in pallid cuckoosCuculus pallidususing reflectance spectrophotometry [J].Proc R Soc Lond B, 273: 1929–1934.
Stoddard MC, Prum RO. 2008. Evolution of avian plumage color in a tetrahedracolor space: A phylogenetic analysis of new world buntings [J].Am Nat, 171: 755-776.
Tojo H, Nakamura S, Higuchi H. 2002. Gape patches in Oriental Cuckoo [J].Ornithol Sci, 1: 145-149.
Vorobyev M, Osorio D, Bennett ATD, Marshall NJ, Cuthill IC. 1998. Tetrachromacy, oil droplets and bird plumage colours [J].J Comp Physiol A, 183: 621-633.
Wang Z, Jia CX, Sun YH. 2004. Parasitized breeding and nestling growth in Oriental Cuckoo [J].Chn J Zool, 39: 103-105.
Yang CC, Cai Y, Zhang SP, Liang W. 2009. Quantitative analysis of bird egg color by using fiber spectrophotometer [J].Chn J Ecol, 28: 346-349.
Yang CC, Cai Y, Liang W. 2010 a. Brood parasitism and egg mimicry on Brownish-flanked Bush Warbler (Cettia fortipes) by Lesser Cuckoo (Cucuculs poliocephalus) [J].Zool Res, 31: 555-560.
Yang CC, Liang W, Cai Y, Shi SH, Takasu F, Møller AP, Fossøy F, Moksnes A, Røskaft E, Stokke BG. 2010 b. Coevolution in action: Disruptive selection on egg colour in an avian brood parasite and its host [J].PLoS ONE, 5(5): e10816.
Yoshino T. 1999. Cuckoos-brood Parasitic Birds in Japan [M]. Tokyo: Bunichisogoshuppan.
Zhang WF. 1980. Field Guide to the Birds of Taiwan [M]. Taiwan: Tunghai University.
以鸟类视觉模型揭示中杜鹃对冠纹柳莺的卵色模拟
杨灿朝1,2, 蔡 燕1, 梁 伟1,*
(1.海南师范大学 生命科学学院,海南 海口571158; 2.中山大学 生命科学学院有害生物控制与资源利用国家重点实验室,广东 广州510275)
于2009年4—7月, 采用光谱仪量化卵色和建立鸟类视觉模型的方法, 在贵州宽阔水自然保护区对中杜鹃(Cuculus saturatus)寄生冠纹柳莺(Phylloscopus reguloides)的卵色模拟进行了研究。中杜鹃产白色卵带极少数而微小的棕色斑, 明显大于宿主卵, 重2.06 g, 体积1.91 cm3。从人眼看, 中杜鹃卵对宿主卵在很大程度上是模拟的, 但视觉模型表明, 两者的卵色在色调和色度上都完全分离, 揭示了人眼探测不到的卵色模拟情况。该文首次对中杜鹃的雏鸟特征进行描述, 在4日龄以后雏鸟嘴裂中出现三角形黑斑, 并随着日龄的增长而更加明显, 这种特征在霍氏中杜鹃(C. optatus)的雏鸟中也存在, 但未见于其他种类的杜鹃雏鸟。
卵色; 卵色匹配; 视觉模型; 光谱仪; 杜鹃; 冠纹柳莺; 罗宾逊投影
Q959.732; Q959.739; Q436
A
0254-5853-(2011)04-0451-05
2011-01-06;接受日期:2011-05-19
杨灿朝,中山大学博士后,研究方向为鸟类行为与生态学
10.3724/SP.J.1141.2011.04451
date: 2011-01-06; Accepted date: 2011-05-19
s: This work was supported by National Natural Science Foundation of China (30860044; 31071938); Program for New Century Excellent Talents in University (NCET-10-0111);China Postdoctoral Science Foundation (20110490967) funded project
*Corresponding author (通信作者), Fax: +86-898-65818360, E-mail: liangwei@hainnu.edu.cn
杂志排行

Zoological Research的其它文章
- 悬尾应激对小鼠空间记忆及其反转学习的损伤效应
- Behavioral migration diversity of the Yangtze River Japanese Eel, Anguilla japonica, based on otolith Sr/Ca ratios
- Afferent and efferent pathways in the visual system of the freshwater snail Planorbarius corneus
- Notch signaling dependent differentiation of cholangiocyte-like cells from rhesus monkey embryonic stem cells
- Metabolism and thermoregulation between Mrs Hume’s Pheasant (Syrmaticus humiae) and Elliot’s Pheasant (S. ellioti)
- Localization of stationary pronuclei during conjugation of Paramecium as indicated by immunofluorescence staining