白桦木聚糖内糖基转移酶XET基因序列及表达1)
2011-06-13王玉成吴英杰
张 岩 王玉成 吴英杰 王 超
(林木遗传育种与生物技术教育部重点实验室(东北林业大学),哈尔滨,150040)
木聚糖内糖基转移酶(xyloglucan endotransglycosylase,XET)是所有植物细胞壁重建过程中的一个关键酶,它利用转糖基机制在植物细胞壁中进行木聚糖多聚体的切割和组装[1]。在研究豌豆茎的伸长时发现,木聚糖链的酶切断裂,可使细胞壁膨胀疏松,促进细胞生长[2]。Nishitani首次纯化了XET蛋白[3-4],XET一经发现便在细胞分化、扩展和特化等多方面受到重视,有人甚至认为它可能是植物生长机制中的核心酶。目前已经从拟南芥(Arabidopsis thaliana)、大豆(Glycine max)、蕃茄(Solanum lycopersicum)、小麦(Triticum aestivum)、大麦(Hordeum vulgare)、玉米(Zea mays)等作物中分离出10多个XET基因成员,分别有不同的功能。陈昆松等[5]在用外源乙烯处理诱导猕猴桃软化的过程中发现XET活性快速增加,促进了细胞壁的膨胀疏松。Lu等[6]在香蕉果实软化过程中发现MA-XETI与香蕉果肉软化相关,并且与果皮的硬度密切相关。近些年,XET活性与木质部生长关系的研究逐渐受到重视。Israelsson等[7]对过量产生GA的转基因植株检测分析表明,增加木材形成速率的酶中就包括XET。桉树中实时RT-PCR分析表明,XET在次生木质部的表达水平是在叶子中表达的360倍,这说明了在次生木质部细胞壁的形成中XET蛋白起着重要作用[8]。与韧皮部相比,XET在拟南芥木质部有大量的表达,表明在木质部组织中,XETs可能参与了木葡聚糖链的移位使木质部细胞膨胀[9]。
白桦(Betula platyphyllaSuk.)为桦木科桦木属植物,是目前较为广泛应用的纸浆材树种之一。研究其次生细胞壁形成过程中的相关基因,对改良白桦的纸浆材特性具有重要的意义。笔者从白桦形成层相关组织cDNA文库中克隆得到了白桦XET基因全长cDNA序列,并对其进行了序列分析及蛋白质高级结构预测,利用Semi-quantative RT-PCR方法分析了该基因在根茎叶不同器官的表达情况,为进一步研究白桦XET基因的功能奠定基础。
1 试验方法
1.1 白桦XET基因的克隆和序列分析
本研究构建了白桦形成层相关组织cDNA文库,对文库克隆进行随机测序,运用NCBI的BLASTX和BLASTN程序对获得的序列进行同源性比对,获得XET的全长cDNA序列。
利用NCBI ORF Finder确定XET基因的开放读码框;利用NCBI的BlastP寻找保守区,预测蛋白所属家族;用ProtParam(http://au.expasy.org/cgi-bin/protparam)分析蛋白质的基本理化性质;用BlastX寻找相似性序列,并选择与其相似性较高的另外7种植物的XET蛋白氨基酸序列,用多序列联配程序ClustalX(1.83)进行多序列比对,构建XET基因进化树。
利用ProtScale软件、HNN软件等在线分析α-螺旋、β-转角、无规卷曲以及延伸连等蛋白的二级结构;利用网站http://au.expasy.Org/tools/中的 Swiss-Model程序进行同源建模,预测XET蛋白的空间结构。
1.2 sqRT-PCR鉴定XET基因在不同器官的表达
培养白桦组培苗[10]。选取生长旺盛的单株,洗去培养基,吸干水分。于根、茎、叶3个部位取材,液氮处理,置于冰箱中-80℃保存。
CTAB法提取白桦根、茎、叶总RNA,经DNaseI(Promega)消化处理,去除DNA污染。取1 μg总RNA进行反转录,反转录反应体系和操作参照PrimerScriptTMRT Reagent Kit(TaKaRa)。将反转录产物稀释10倍,用作sqRT-PCR的模板。选择TUB作为内参基因。内参和XET基因的定量PCR引物如表1所示。
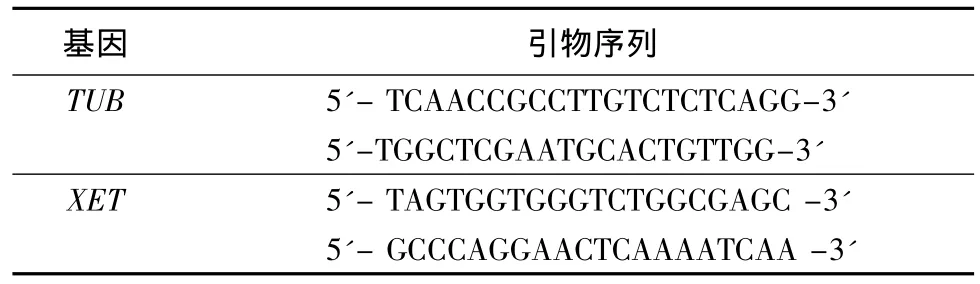
表1 sqRT-PCR引物序列
sqRT-PCR 反应体系:10×Buffer 2 μL,dNTP(2.5 μmol/L)0.4 μL,上、下游引物各 1 μL(20 μmol/L),TaqDNA Polymerase 0.25 μL,cDNA 2 μL,去离子水补至20 μL,设置3 次重复。PCR反应程序:94℃预变性3 min,94℃变性30 s、58℃退火30 s、72℃延伸30 s,循环30次,72℃延伸7 min。反应产物利用1%EB-琼脂糖凝胶电泳检测。运用Quantity One Analysis Software对电泳结果相对定量,通过调整上样量使内参均一化,按照调整后内参的上样量分析XET基因的表达量,进而用Quantity One Analysis Software得出定量分析的数据。
2 结果与分析
2.1 白桦XET基因全长cDNA的获得及序列分析
通过对白桦形成层相关组织cDNA文库随机测序和序列比对分析,获得白桦XET基因全长cDNA序列。该cDNA编码区序列自74位的ATG起,止于954位的TAG,全长882 bp,见图 1。3'UTR 135 bp,5'UTR 73 bp。开放读码框编码由 293个氨基酸残基组成的多肽,编码蛋白的分子质量为33.1 ku,理论等电点为5.49,为酸性蛋白质;负电荷残基(Asp+Glu)数为34,正电荷残基(Arg+Lys)数为23个;不稳定系数为29.32,为稳定的蛋白质。

图1 白桦XET基因序列及由此推导的氨基酸序列
2.2 白桦XET蛋白家族预测
利用NCBI的BlastP寻找保守区,结果表明白桦XET基因具有一个保守区,属于糖基水解酶16超家族,见图2。利用Pfam程序找到一个XET C-terminu超家族的保守区。
2.3 XET多序列比对及进化树
对XET基因进行BlastX比对分析,发现各物种间该基因的蛋白质序列相似度较高,选取8种植物XET蛋白的氨基酸序列,运用Clustalx(1.83)进行多序列比对,并创建进化树,如图3、图4所示。从图中可以看出,各物种的XET蛋白质序列相对保守,但是N端序列保守性不高,这与蛋白家族保守区预测的结果相似,该基因的大部分序列位于保守区内。进化树分析得出,白桦XET基因与杨树(Populus tremula)、蓖麻(Actinidia deliciosa)XET基因的同源性较其他物种高。
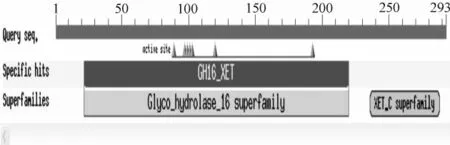
图2 BlastP推导的XET氨基酸序列的保守区预测

图3 8个物种XET蛋白的多序列比对分析

图4 XET基因进化树
2.4 白桦XET蛋白二级结构
运用ExPASy Proteomics tools中的SOPMA软件在线分析XET蛋白的二级结构,如图5所示。组成白桦XET蛋白的293个氨基酸中,25个氨基酸可能形成α-螺旋,104个氨基酸可能形成延伸,147个氨基酸可能形成无规卷曲,17个氨基酸可能形成β-转角。
2.5 白桦XET蛋白三级结构
通过在ExPDB晶体图像数据库中(http://www.rcsb.org/pdb/)搜索相似的序列,得到一个同源性较高的蛋白质结构数据。以此序列的结构信息为原子模型登录Swiss-Model服务器上进行结构排列,并运用结构仿真模拟(ProModⅡ程序)和能量最小化分析(GROMOS96程序)构建目标序列的结构[11-13],模建结果见图 6。利用 Swiss-viewer3.7 对模建结果进行检测,计算得出Ramachandran图(图7),由图可见,模拟得到的白桦XET蛋白三维结构的Φ角和Ψ角有95%位于Ramachandran图中合理区域,从理论上表明此三维结构是可靠的。
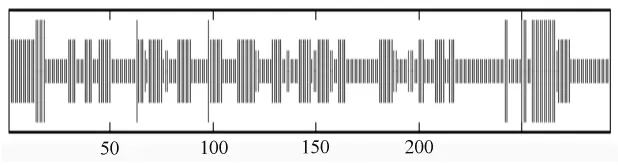
图5 白桦XET蛋白的二级结构分析

图6 白桦XET蛋白的空间结构

图7 Ramachandran图
2.6 白桦XET基因在不同器官的表达
通过sqRT-PCR方法分析白桦XET基因在根茎叶3个器官的表达情况,首先对内参基因PCR产物的电泳结果进行定量,通过调整上样量,使内参基因表达量均一化,然后依据调整后的上样量对相应的XET基因PCR产物进行电泳检测,如图8;运用Quantity One Analysis Software对电泳结果进行相对定量,结果显示,XET基因在根、茎和叶3个器官内都有较高的表达水平,其中在茎中表达量最高,在叶中表达量相对最低。
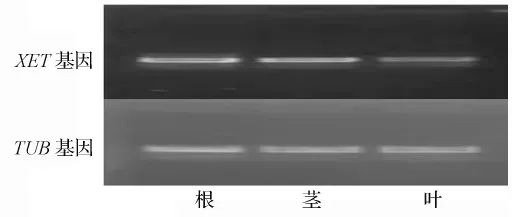
图8 白桦XET基因在不同器官的表达
3 结论与讨论
XET能够催化木聚糖链的裂解来使细胞壁松弛,在调节细胞分化、扩展和重构过程中起作用,进而调节果实软化和木质部发育等,属于糖基水解酶家族的一员。糖基水解酶家族是一组广泛存在的,水解两个或更多碳水化合物之间或者碳水化合物和非碳水化合物半族之间糖苷接点的酶。本文在XET蛋白家族预测中找到了一个糖基水解酶超家族的保守区和XET C-terminu超家族保守区,确定所获得基因为XET基因。运用同源模建方法,以原子质量为1 μ序列的结构信息为原子模型,构建了白桦XET基因编码蛋白的三维结构,并利用Swiss-viewer 3.7对模建结果进行检测,计算得出Ramachandran图。Ramachandran图是反映立体化学质量(stereochemical quality)的参数,它通过分析Φ角和Ψ角的分布方式大致评估模拟的结构是否与自然结构趋势相同。黄色区域是最理想的Φ角和Ψ角分布区域,而蓝色区域外部则为不合理区域[14]。由图7可见,模拟得到的白桦XET蛋白的三维结构的Φ角和Ψ角有95%位于Ramachandran图中合理区域,从理论上表明该XET蛋白的三维结构是可靠的。序列决定功能,这为今后研究白桦XET蛋白结构和生物学功能奠定了基础。
细胞壁的形态建成是一系列酶作用的结果,尤其是细胞壁修饰酶,有研究表明XET蛋白就是一类重要且与细胞伸长密切相关的细胞壁松弛酶。植物细胞初生细胞壁的形成是由同步生长和嵌入生长共同决定的,此时,编码细胞壁修饰酶类的基因会大量表达。有研究表明,杨树中16个XET基因在发育的木质部中表达,其中5个高度表达,1个在木质部中特异表达[15]。XET基因在杨树应压木中表达量升高,说明其参与了次生细胞壁形成的生理程序[16]。本研究通过sqRT-PCR方法分析了白桦XET基因在生长中苗木不同器官的表达情况,结果显示,XET在根,茎和叶中都有较高的表达水平,说明生长中的根、茎和叶组织正在进行细胞的形态建成,XET在白桦这3种组织的细胞壁建成过程中发挥了作用。此XET基因在白桦根和茎中的表达水平高于叶组织,而且Chao Wang等[17]研究发现此白桦XET基因在形成层发育的不同时期具有时序性表达特性,推测此XET基因与维管组织次生细胞壁的建成有关。
[1]Fry S C.Polysaccharide-modifying enzymes in the plant cell[J].Annu Rev Plant Physiol Plant Mol Biol,1995,46:497-520.
[2]McDougall G J,Fry S C.Xyloglucan oligosaccharides promote growth and activate cellulose:evidence for a role of cellulose in cell expansion[J].Plant Physiol,1990,93:1042-1048.
[3]Fry S C,Smith R C,Renwick K F,et al.Xyloglucan endotransglycosylase,a new wall-loosening enzyme activity from plants[J].Bio Chem,1992,282:821-828.
[4]Okazawa K,Sato Y,Nakagawa T,et al.Molecular cloning and cDNA sequencing of endoxyloglucan transferase,a novel class of glycosyltransferase that mediates molecular grafting between matrix polysaccharides in plant cell walls[J].Biology Chem,1993,268:25364-25368.
[5]陈昆松,李方,张上隆.猕猴桃果实成熟进程中木葡聚糖内糖基转移酶mRNA水平的变化[J].植物学报,1999,41(11):1231-1234.
[6]Lu W J,Nakano R,Kubo Y,et al.Cloning and expression analysis of an XET cDNA in the peel and pulp of banana fruit ripening and softening[J].Acta Botanica Sinica,2004,46(3):355-362.
[7]Israelsson M,Erikssonl M,Hertzberg M,et al.Changes in gene expression in the wood-forming tissue of transgenichybrid aspen with increased secondary growth[J].Plant molecular biology,2003,52(4):893-903.
[8]Paux E,amasloukht M,Ladoucel N,et al.Identifieation of genes preferentially expresse during wood formation in EucalyPtus[J].Plant Molecular Biology,2004,55(2):263-280.
[9]Oh S,Park S,Han K H.Transcriptional regulation of secondary growth in arabidopsis thaliana[J].Journal of Experimental Botany,2003,393(54):2709-2722.
[10]陶静,詹亚光,姜静,等.白桦组培再生系统的研究(Ⅰ)[J].东北林业大学学报,1998,26(5):6-9.
[11]Arnold K,Bordoli L,Kopp J.The swiss-model workspace:a webbased environment for protein structure homology modelling[J].Bioinformatics,2006,22:195-201.
[12]Schwede T,Kopp J,Guex N.Swiss-model:an automated protein homology-modeling server[J].Nucleic Acids Research,2003,31:3381-3385.
[13]Guex N,Peitsch M C.Swiss-model and the Swiss-Pdbviewer:an environment for comparative protein modelling[J].Electrophoresis,1997,18:2714-2723.
[14]胡松年.基因表达序列标签(EST)数据[M].杭州:浙江大学出版社,2005:206-215.
[15]Nishikubo N,Takahashi J,Roos A A,et al.Xyloglucan endotransglycosylase-mediated xyloglucanre arrangements in developing wood of hybrid aspen[J].Plant Physiol,2011,155(1):399-413.
[16]Nishikubo N,Awano T,Banasiak A,et al.Xyloglucan endotransglycosylase(XET)functions in gelatinous layers of tension wood fibers in poplar-a glimpse into the mechanism of the balancing act of trees[J].Plant Cell Physiol,2007,48(6):843-55.
[17]Chao Wang,Yucheng Wang,Guiping Diao,et al.Isolation and characterization of expressed sequence tags(ESTs)from cambium tissue of birch(Betula platyphyllaSuk.)[J].Plant Mol Biol Rep,2010,28:438-449.
