Ubc9基因重组腺病毒的构建及其在HeLa细胞中的表达
2011-06-08李薇刘晓萍徐祥谭艳于业军李艳君任书亭贾跃伟
李薇,刘晓萍,徐祥,谭艳,于业军,李艳君,任书亭,贾跃伟
为了进一步研究 Ubc9 对细胞功能的影响,我们构建带有红色荧光标记(RFP)的含 Ubc9 基因的腺病毒载体,观察 Ubc9 在 HeLa 细胞中的表达情况,为今后研究 Ubc9 的功能提供一个工具。
1 材料与方法
1.1 质粒、菌株和主要生化试剂
穿梭质粒 pAdTrack-TO4、腺病毒骨架质粒pAdeasy-1、pCMV6-XL6-Ubc9 质粒、大肠杆菌DH5a、BJ5183 及腺病毒包装细胞 HEK293 细胞由第三军医大学大坪医院外研所赠予;转染试剂PolyJetTM购自美国 SignaGen 公司;质粒小量提取试剂盒、胶回收试剂盒以及 DNA 标记物购自北京康为世纪生物科技有限公司;无内毒素质粒大量制备试剂盒购自北京百泰克生物技术有限公司;DMEM 培养基、血清、胰蛋白酶购自美国 Gibco公司;限制性内切酶、T4 DNA 连接酶购自大连Takara 公司;琼脂糖及各种生化制剂由凌飞公司进口分装;PCR 引物由上海生工公司合成;基因测序由上海英潍捷基有限公司完成。
1.2 方法
1.2.1 PCR 法扩增目的 Ubc9 基因 以 pcmv6-xl6-Ubc9 质粒为模板进行 PCR 扩增。在引物中分别引入 BamH I 和 Xho I 的酶切位点,上游引物:5’ GGATCCATGTCGGGGATCGCCCTCAG 3’;下游引物:5’ TCCGCTCGAGTTATGAGGGCGCAAA CTTC 3’。PCR 反应参数:94℃预变性 2min,94℃变性 30 s,60℃退火 30 s,72℃延伸 30 s,循环 30 次,最后 72℃延伸 5min。PCR 产物经 2%琼脂糖凝胶电泳,切胶回收获得目的片段。
1.2.2 腺病毒穿梭质粒 pAdTrack-CMV-Ubc9 的构建 将 PCR 扩增的 Ubc9 基因产物和带有RFP 的穿梭质粒 pAdTrack-CMV 分别以 BamH I和 Xho I 进行双酶切,经 2%琼脂糖凝胶电泳鉴定并纯化回收 Ubc9 片段及 pAdTrack-CMV 穿梭质粒。用 T4 DNA 连接酶过夜连接,转化大肠杆菌 DH5a。挑选转化菌落,小量快速提取重组质粒。通过限制性酶切和测序分析进行鉴定。同时,以不含目的基因 Ubc9 的穿梭质粒 pAdTrack-CMV 作为载体对照。
1.2.3 含有 pAdEasy-1 的 BJ5183 菌的制备 挑取单一克隆 BJ5183 菌接种在含 LB 培养基摇瓶中,37℃,200 r/min,培养 16 h。取 200 μl 菌液加入 20 ml 新鲜的 LB 培养基,37℃,260 r/min,培养 2 h,直至 OD 值为0.4~0.5。取 1.5 ml 菌液置 1.5 ml 离心管中,冰上放置 10min。4℃,3000×g离心 10min,弃上清。
加入 1 ml 0.1 mol/L 的 CaCl2轻轻吹打,充分悬浮,冰浴 10min。4℃,3000×g离心 10min,弃上清。加 1 ml 0.1 mol/L 的 CaCl2重悬沉淀,冰浴 30min。4℃,3000×g离心 10min,弃上清。用 100 μl 0.1 mol/L 的 CaCl2重悬沉淀,即感受态制备完成。
在感受态细胞中,加入 5 μl pAdEasy-1 质粒,冰浴 30min。42℃热刺激 90 s。冰浴 5min。加LB 培养基 900 μl,37℃,200 r/min 培养 30min后 3000×g离心 3min。收集沉淀并留 100 μl 上清,弃多余部分,混匀,涂布到含氨苄青霉素的 LB板。37℃培养箱倒置培养,约 16 h。挑取单个菌落,扩增培养,获取含有腺病毒骨架质粒 pAdEasy-1的 BJ5183 感受态细菌,储存待用。
1.2.4 pAdEasy-1/Ubc9 腺病毒载体的构建 取1 μg pAdTrack-CMV-Ubc9 质粒 DNA,PmeI 酶切线性化,乙醇沉淀回收后,转化至含有腺病毒骨架质粒 pAdEasy-1 的 BJ5183 感受态细菌中,混匀,冰上静置 30min。42℃热刺激 90 s,冰浴5min,加 LB 培养基 900 μl,37℃,200 r/min 培养 20min。接种于含有卡那霉素的平板上,孵育16 h。挑取较小的 10 个克隆扩增培养,提取质粒。用 PacI 酶切鉴定质粒,将鉴定正确的重组质粒再次转化感受态 DH5a 菌。扩增培养,大量提取pAdEasy-1/ Ubc9 的重组腺病毒质粒。并用 PacI酶切使之线性化,保存于 –20℃待用。
1.2.5 重组腺病毒的包装及扩增 于转染前 12~18 h 铺 293 细胞,当细胞密度达 50%~70%时。采用脂质体法将 5 µg 线性化的重组质粒 pAdEasy-1/Ubc9 转染至 293 细胞中,12 h 后换新鲜培养基(具体操作参照 PolyJetTM说明进行转染,同时转染空载体作对照)。37℃5%CO2培养 5~7 d,待细胞出现病变时(7~10 d)收集细胞。于 –70℃和37℃水浴中反复冻融 4 次,3000×g离心 10min,收集上清。上清液中含原始重组腺病毒(Ad-Ubc9)。取重组病毒液重复感染 293 细胞 2~3 次,以大量扩增病毒。分装病毒液,保存于 –70℃。
2016年8月,游成令的女儿患上“急性普通B淋巴细胞L1型白血病”。这突如其来的噩耗,给这对农村走出来的夫妻带来了巨大的精神和经济压力。游成令后悔以前没有多关心女儿一点。在工作和家庭之间,她从来都是把工作放在第一位。
1.2.6 重组腺病毒感染 HeLa 细胞 取 HeLa细胞铺于 6cm 平皿中,密度为106个/ml。贴壁后吸弃培养液,用灭菌 PBS 洗 2~3 遍。加入含Ad-Ubc9 病毒液感染 HeLa 细胞。培养 2 h 后弃病毒液,PBS 洗 3 遍,加入含 0.5%小牛血清的DMEM 培养液培养 72 h。
1.2.7 Western blot 检测重组腺病毒蛋白的表达 感染 3 d 后收集细胞,提取总细胞蛋白(RIPA法)。取 10 µg 蛋白,用 12%SDS-PAGE 分离。PVDF 膜在转印液中平衡 10min,然后遵循胶在负极,膜在正极的原则进行转印,电压 90 V,4℃,2 h;取出 PVDF 膜用丽春红染液染色,约 5min,蛋白条带出现后,用去离子水漂洗;将 PVDF 膜置于 100 g/L 脱脂奶粉溶液中,封闭 2 h,兔抗Ubc9 血清(1∶1000),4℃过夜,TPBS 洗涤 3 次,10min/次;二抗浓度为1∶10000,室温孵育1 h,TPBS 洗涤 3 次,10min/次。ECL Kit 检测结果(同时用未感染病毒的 HeLa 细胞作对照)。
2 结果
2.1 目的片段(Ubc9)的扩增
以 pcmv6-xl6-Ubc9 质粒为模板,PCR 扩增出477 bp 的 Ubc9 片段。产物经 2%琼脂糖凝胶电泳可观察到约 477 bp 的条带,与预期结果相符(图1)。

图1 含 Ubc9 基因质粒 PCR 扩增结果Figure 1 Amplification of Ubc9 gene by PCR
2.2 酶切鉴定重组穿梭质粒 pAdTrack-CMV-Ubc9
BamH I 和 Xho I 双酶切产物电泳可观察到约 9 kb 和 477 bp 的 DNA 条带,经过碱基测序,Ubc9基因未见突变,表明含有Ubc9基因的穿梭载体构建成功,可用于后续实验(图2)。
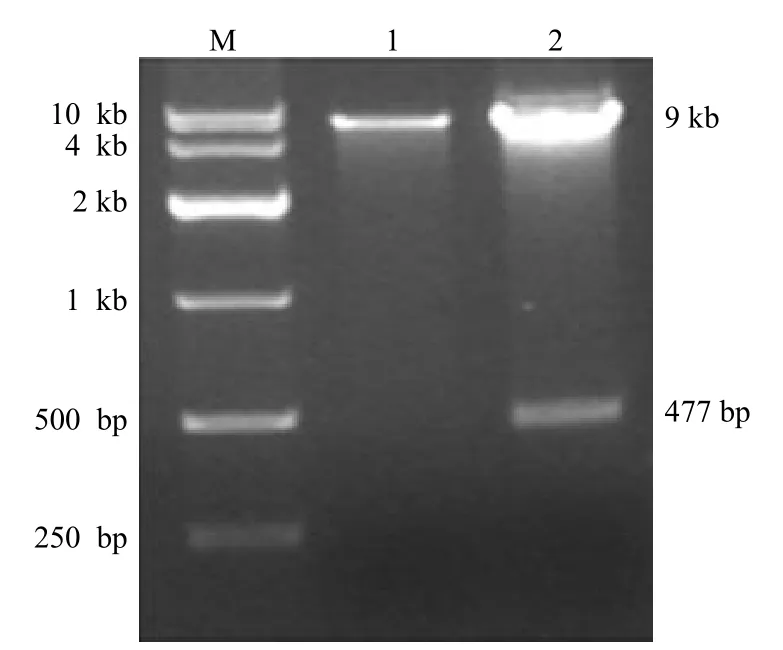
图2 琼脂糖凝胶电泳鉴定穿梭质粒 pAdTrack-CMV-Ubc9的酶切结果Figure 2 Restriction enzyme digestion analysis of shuttle plasmid pAdTrack-CMV-Ubc9 vectors
2.3 酶切鉴定重组质粒 pAdEasy-1/Ubc9
pAdEasy-1/Ubc9 质粒经 Pac I,酶切后获得大小两个片段,即 1 个大约 30 kb 和 1 个 4.5 kb的片段,表明穿梭质粒 pAdTrack-CMV-Ubc9 与腺病毒骨架质粒 pAdEasy-1 同源重组成功(图3)。
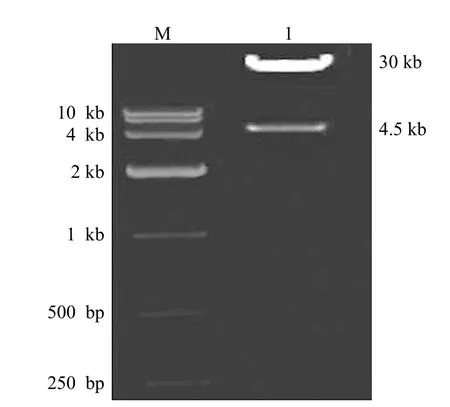
图3 琼脂糖凝胶电泳鉴定 pAdEasy-1/Ubc9 的酶切结果Figure 3 Identification of recombinant adenovims vector by restriction enzyme digestion
2.4 重组腺病毒 Ad-Ubc9 的包装
将带有红色荧光标记的线性化 pAdEasy-1/Ubc9 转染 293 细胞后,在荧光显微镜下,Ad-Ubc9 成功包装细胞呈现 RFP 阳性(图4)。
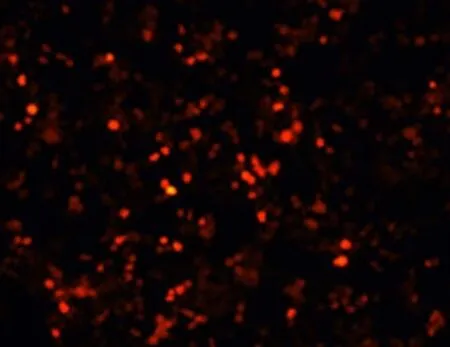
图4 转染 293 细胞后 RFP 的表达Figure 4 RFP expression after plasmid transfected HEK293 cells
2.5 Ubc9 基因在靶细胞中的表达
Western blot 检测可见感染重组腺病毒 Ad-Ubc9 的 HeLa 细胞中呈现 Ubc9 蛋白高表达(图5)。
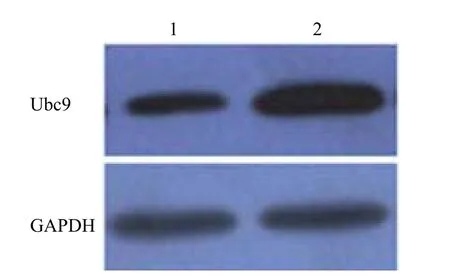
图5 Ubc9 在 HeLa 细胞中表达Figure 5 Analysis Ubc9 expression in HeLa cells by western blot
3 讨论
SUMO 的生化反应过程是一系列独特的活化酶级联反应过程,最终使其与底物蛋白结合完成蛋白质修饰,从而调节细胞活动。自 SUMO 发现的十多年来,大量研究集中在酶反应体系和蛋白质相互作用方面,而 SUMO 修饰的功能学研究甚少。Ubc9 作为唯一的 E2 酶,SUMO 化修饰的完成依赖于 Ubc9 的表达,近年来对 Ubc9 的研究成为热点。有报道 Ubc9 基因的敲除可导致鸡细胞的双核细胞数量增加和鼠类细胞的细胞核形态异常以及染色体的缺失[9-10]。在果蝇细胞中,Ubc9 功能的缺失引起造血细胞的有丝分裂紊乱[11]。酵母菌中Ubc9 基因突变,会使其对引起 DNA 损伤的因素更加敏感[12-13]。为了更广泛地研究 Ubc9在细胞中的功能,我们构建表达 Ubc9 的腺病毒表达载体。
重组腺病毒是复制缺陷型的腺病毒载体,也是目前基因治疗研究和临床试验中应用最广泛的病毒载体。传统上,双载体(即穿梭质粒和骨架质粒)同源重组过程是在哺乳动物细胞中来实现的。然而,由于哺乳动物细胞中同源重组的低效性以及某种程度上的不可预见性,使得该过程成为重组腺病毒制备的限速步骤。与传统方法不同的是,该试验中两种质粒的重组是在细菌(BJ5183)中而非真核细胞中完成。由于质粒转化细菌的效率极高,细菌繁殖速度快,且 BJ5183 菌中的 Rec A 可表达很强的重组酶活性,因此,细菌中同源重组法克服了细胞内质粒共转染效率低和同源重组率低的缺点,大大缩短了实验周期,且显著提高了成功率。另外,这种重组质粒转染哺乳动物包装细胞株后,在共转染的基因编码的红色荧光(RFP)的指示下,阳性转染重组病毒的细胞容易被观察和追踪。该系统加快了各种目的重组腺病毒的制备和鉴定过程。
本实验通过限制性酶切分析及碱基序列测定,均证明已正确构建重组穿梭质粒 pAdTrack-CMVUbc9。在 HEK293 细胞中成功包装重组腺病毒Ad-Ubc9。本研究所产生的重组腺病毒 Ad-Ubc9能在体外感染 HeLa 细胞,并成功表达出 Ubc9 蛋白,为进一步研究 Ubc9 基因在细胞中的功能奠定了基础。
[1]Li XD, Yang S, Qiu YF, et al.SUMOylation: an important post-translational modification in vivo.Chin J Cell Biol, 2008, 30:142-146.
[2]Sternsdorf T, Jensen K, Freemont PS.Sumo.Curr Biol, 2003, 13(7):R258-R259.
[3]Johnson ES.Protein modification by SUMO.Annu Rev Biochem,2004, 73:355-382.
[4]Hayashi T, Seki M, Maeda D, et al.Ubc9 is essential for viability of higher eukaryotic cells.Exp Cell Res, 2002, 280(2):212-221.
[5]Seufert W, Futcher B, Jentsch S.Role of a ubiquitin-conjugating enzyme in degradation of S- and M-phase cyclins.Nature, 1995,373(6509):78-81.
[6]McDoniels-Silvers AL, Nimri CF, Stoner GD, et al.Differential gene expression in human lung adeno-carcinomas and squamous cell carcinomas.Clin Cancer Res, 2002, 8(4):1127-1138.
[7]Mo YY, Yu Y, Theodosiou E, et al.A role for Ubc9 in tumorigenesis.Oncogene, 2005, 24(16):2677-2683.
[8]Moschos SJ, Athanassiou H, Edington H, et al.Proteomic analysis of melanoma-in fi ltrated lymph nodes gains insights into the molecular pro fi le of metastatic melanoma.Proc Amer Assoc Cancer Res,2005(46).
[9]Hayashi T, Seki M, Maeda D, et al.Ubc9 is essential for viability of higher eukaryotic cells.Exp Cell Res, 2002, 280(2):212-221.
[10]Nacerddine K, Lehembre F, Bhaumik M, et al.The SUMO pathway is essential for nuclear integrity and chromosome segregation in mice.Dev Cell, 2005, 9(6):769-779.
[11]Chiu H, Ring BC, Sorrentino RP, et al.dUbc9 negatively regulates the Toll-NF-kappa B pathways in larval hematopoiesis and drosomycin activation in Drosophila.Dev Biol, 2005, 288(1):60-72.
[12]Mao Y, Sun M, Desai SD, et al.SUMO-1 conjugation to topoisomerase I: A possible repair response to topoisomerase-mediated DNA damage.Proc Natl Acad Sci U S A, 2000, 97(8):4046-4051.
[13]Jacquiau HR, vanWaardenburg RC, Reid RJ, et al.Defects in SUMO(small ubiquitin-related modi fi er) conjugation and deconjugation alter cell sensitivity to DNA topoisomerase I-induced DNA damage.J Biol Chem, 2005, 280(25):23566-23575.
