吗啡依赖大鼠不同脑区肌动蛋白及其结合蛋白的表达变化
2011-05-31闫彩珍郝晓玲王天魁
闫彩珍,于 亮,刘 川,郝晓玲,王天魁
(河北医科大学药理学教研室,河北石家庄 050017)
阿片类药物成瘾的主要特征表现为强迫性和持续性药物使用。目前研究认为药物成瘾行为的持久存在,可能与转录因子短暂调节所致的突触结构及相关神经回路的结构重塑有关[1]。成瘾药物所致持续性结构改变的一个备受关注的例子是,反复使用成瘾药物可引起与情感动机、奖赏和学习有关脑区神经元树突棘的结构产生持久改变[2],提示突触联系发生了可塑性变化。
树突棘是神经元之间形成兴奋性突触的位置,它们在神经元中呈现不同的形状,目前已证实树突棘形态动力学是由肌动蛋白动态调节的,肌动蛋白在棘突形态形成中发挥重要作用[3]。肌动蛋白是真核细胞最主要的骨架成份之一,它以单体(G-actin)和纤维状(F-actin)两种形式存在,并且单体与纤维状肌动蛋白之间可以相互转换。在神经元树突棘可以观察到与肌动蛋白有关的运动性,荧光漂白恢复技术分析显示树突棘F-actin可以发生快速的循环。药理学研究显示,控制肌动蛋白循环可以导致树突棘发生形态学改变,提示肌动蛋白在调节神经元树突棘的形态和动力学中发挥重要作用[4]。研究显示,长期吗啡处理可增加伏隔核F-actin的含量[5],条件性位置厌恶大鼠杏仁核和背侧海马肌动蛋白发生重构[6],但是对吗啡依赖不同时程大鼠不同脑区肌动蛋白的重构并不清楚,同时吗啡影响细胞骨架肌动蛋白表达及重构的分子机制也尚未阐明。
本研究拟从影响神经元形态和功能的骨架肌动蛋白入手,采用动物行为学、Western blot等方法,研究吗啡依赖大鼠部分脑区肌动蛋白及其结合蛋白p-cofilin的表达变化,探讨骨架肌动蛋白结合蛋白在吗啡诱导的神经元突触可塑中的作用,进一步深化阿片类药物成瘾的分子生物学机制。
1 材料与方法
1.1 材料
1.1.1动物♂ Sprauge Dawley(SD)大鼠,体质量(200±20)g,购自河北省实验动物中心。
1.1.2药物及试剂盐酸吗啡注射液购自沈阳第一制药厂;盐酸纳洛酮注射液购自北京华素制药股份有限公司;Actin抗体购自美国Santa Cruz公司;pcofilin抗体购自美国Cell Signaling公司;BCA蛋白定量试剂盒购自美国PIERCE公司;辣根过氧化酶标记的山羊抗小鼠、羊抗兔抗体购自美国KPL公司;DAB显色试剂盒购自北京中山生物技术有限公司。
1.1.3主要仪器5417R型低温离心机,德国Eppendorf公司;超高速离心机,美国 Beckman公司;AG135型电子天平,瑞士Mettler Toledo公司;1004型恒温水浴箱,德国GFL公司;SHY-200B型水平摇床,德国Leica公司;Millipore纯水仪,美国Millipore公司;DYY-11型多用电泳仪、DYC-Z22A型电泳槽,北京六一仪器厂;凝胶成像仪,美国Themo Foma公司。
1.2 方法
1.2.1大鼠吗啡依赖模型建立♂ SD大鼠42只,随机分为吗啡依赖1周组、吗啡依赖2周组和吗啡依赖4周组,各依赖组均设对照。每组7只大鼠,分笼喂养,自由进食、饮水,动物适应新环境3 d后进行实验。采用腹腔递增注射盐酸吗啡建立吗啡依赖模型,连续注射7 d,每天3次,剂量按5,10,15,20,30,40,50 mg·kg-1逐日递增,对照组给予同体积生理盐水。d 8,在各组大鼠中每组随机选取2只,于吗啡注射前2 h皮下注射盐酸纳洛酮5 mg·kg-1,观察戒断症状,并进行评分,判断大鼠吗啡依赖模型是否建立成功。然后将吗啡依赖1周组与对照组大鼠断头处死,分离相关脑区。吗啡依赖2周组、4周组继续分别给予50 mg·kg-1盐酸吗啡,每天1次,连续注射1或3周。在最后一次给药后5 h断头处死大鼠,迅速分离前额叶皮层、丘脑、海马及纹状体,置液氮快速冷冻后转移至-80℃冰箱保存。
1.2.2戒断症状评分标准[7]① 跳跃、湿狗样抖动、扭体、摇头、打哈欠、扫尾:0分=无;1分=1~5次;2分=6~10次;3分 >10次。② 齿颤、咀嚼(次与次之间至少间隔30 s):0分=无;1分=1~10次;2分=11~20次;3分>20次。③ 流涎、流泪、竖毛、眼睑下垂、激惹和腹泻:0分=无;1分=轻度;2分=中度;3分=明显。
1.2.3脑组织总蛋白的制备脑组织按每100 mg组织加入1 ml的组织蛋白裂解液(50 mmol·L-1Tris-Cl,pH 7.4,150 mmol·L-1NaCl,1 mmol·L-1EGTA,1 mmol·L-1EDTA,1%NP40,1 mmol·L-1DTT,1 mmol·L-1PMSF,1 mg·L-1aprotintin,1 mg·L-1pepstatin A,1 mg·L-1leupeptin),在 DY-89Ⅰ型电动匀浆器下匀浆30 s,4℃ 12 000 r·min-1离心20 min,收集上清,其中包含p-cofilin蛋白成分。
1.2.4蛋白分步提取和Western印迹分析根据G-actin属于胞质可溶性蛋白,F-actin为细胞骨架相关蛋白,采用不同缓冲液对这两种蛋白进行分部提取。脑组织按每100 mg组织加入1 ml的组织蛋白裂解液(20 mmol·L-1Tris-Cl,pH 7.5,1 mmol·L-1Na3VO4,1%Triton X-100,5 mmol·L-1EGTA,1 mmol·L-1PMSF,1 mg·L-1aprotintin,1 mg·L-1pepstatin A,1 mg·L-1leupeptin),在 DY-89Ⅰ型电动匀浆器下匀浆30 s,冰浴30 min裂解后,4℃12 000 r·min-1离心30 min后取上清,即为G-actin的蛋白组分(含肌动蛋白单体G-actin)。沉淀按100 mg加入1 ml裂解液的比例重悬于缓冲液(10 mmol·L-1Tris-Cl,pH 7.5,150 mmol·L-1NaCl,1%Triton X-100,0.1%SDS,1 mmol·L-1脱氧胆酸钠,2 mmol·L-1EDTA,1 mmol·L-1PMSF,1 mg·L-1aprotintin,1 mg·L-1pepstatin A,1 mg·L-1leupeptin)中冰浴30 min,间断涡旋混匀,4℃ 12 000 r·min-1离心30 min后取上清,即为F-actin的蛋白组分[8]。BCA法蛋白定量,各取等量 G-actin组分及相应体积的F-actin组分,经SDS-PAGE电泳分离,电转至PVDF膜上(4 ℃,2h),50 g·L-1脱脂奶粉的TBST封闭1 h,分别加入小鼠β-actin单克隆抗体(1∶500)、兔抗GAPDH多克隆抗体(1∶500),4℃过夜,TBST洗膜3次,每次10 min,然后加入相应辣根过氧化物酶标记的二抗(37℃,1h),DAB显色。采用凝胶成像分析系统对Western blot区带进行定量分析,P-cofilin蛋白的相对含量用其灰度值与GAPDH的灰度值比值来表示,肌动蛋白重构用F-actin/G-actin来表示。
1.2.5统计学分析采用SPSS 15.0软件进行数据分析,Origin7.5软件进行图像处理。实验数据以±s表示,采用单因素方差分析,Dunnett检验进行组间比较。
2 结果
2.1行为学观察大鼠在注射纳洛酮30 min后,吗啡处理大鼠出现明显的湿狗样颤抖、吞咽、站立、跳跃、齿颤、上睑下垂、流涎、腹泻等戒断症状,对戒断症状进行评分,结果见Tab 1。与对照组相比,差异具有统计学意义(P<0.01),说明吗啡依赖大鼠模型建立成功。
Tab 1 The withdrawal score in morphine dependent rats of different groups(±s)

Tab 1 The withdrawal score in morphine dependent rats of different groups(±s)
**P <0.01 vs control.C:control;M:morphine treatment
Group Withdrawal score 3.1 ±2.4 M 27.8 ±5.6 C**
2.2吗啡依赖大鼠前额叶皮质中F-actin/G-actin、p-cofilin蛋白表达的变化与对照组相比,吗啡依赖各组总actin的表达无变化,吗啡依赖1周组大鼠前额叶皮质内F-actin/G-actin的比值没有变化,吗啡依赖2周组和4周组大鼠前额叶皮质内F-actin/G-actin的比值分别上调53%(P<0.05)和102%(P<0.01)。吗啡依赖1周组大鼠前额叶皮质内pcofilin表达量没有变化,吗啡依赖2周组和4周组大鼠前额叶皮质内p-cofilin表达量分别上调93%(P<0.01)和88%(P<0.01)(Fig 1)。
2.3吗啡依赖大鼠海马中F-actin/G-actin、p-cofilin蛋白表达的变化与对照组相比,吗啡依赖1周组和2周组大鼠海马中F-actin/G-actin的比值没有变化,吗啡依赖4周组海马中F-actin/G-actin比值上调94%(P<0.01)。吗啡依赖1周组大鼠海马中p-cofilin蛋白表达量没有变化,吗啡依赖2周组和4周组大鼠海马中p-cofilin蛋白表达量分别上调45%(P<0.05)和30%(P<0.05)(Fig 2)。
2.4吗啡依赖大鼠纹状体中F-actin/G-actin、pcofilin蛋白表达的变化与对照组相比,吗啡依赖各组大鼠纹状体内F-actin/G-actin比值没有变化。吗啡依赖1周组大鼠纹状体内p-cofilin蛋白表达量没有变化,吗啡依赖2周组和4周组的大鼠纹状体内p-cofilin的蛋白表达量分别下调25%(P<0.05)和30%(P<0.05)(Fig 3)。
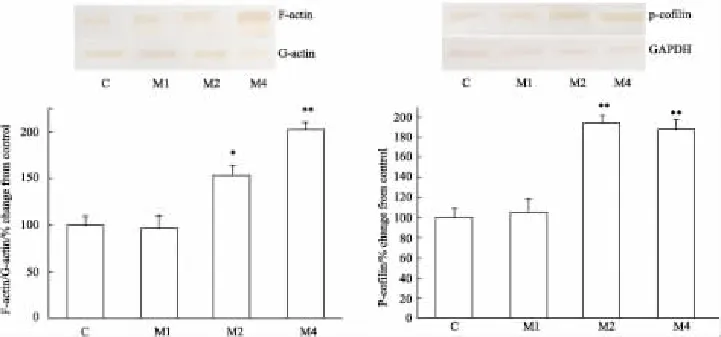
Fig 1 Expressions of F-actin/G-actin and p-cofilin in prefrontal cortex of morphine dependent rats(±s,n=5)

Fig 2 Expressions of F-actin/G-actin and p-cofilin in hippocampus of morphine dependent rats(±s,n=5)
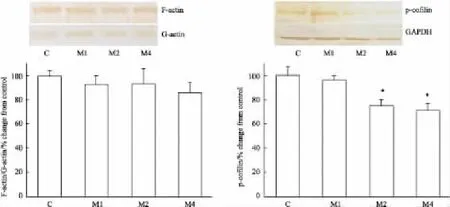
Fig 3 Expressions of F-actin/G-actin and p-cofilin in striatum of morphine dependent rats(±s,n=5)
2.5吗啡依赖1、2和4周组大鼠丘脑中均未发现F-actin/G-actin、p-cofilin蛋白表达量发生明显变化
3 讨论
吗啡滥用可造成神经系统突触的可塑性改变,以往的研究表明[9-10],突触可塑性包括很多环节,其中树突棘的形态学改变被认为是促成各种刺激事件诱导的行为适应性改变的重要基础,各种的刺激性事件就包括吗啡成瘾。这样,吗啡等精神药物诱导的动物模型所表现出来的行为的可塑性与成瘾性被联系到树突蛋白的持续性适应性改变,现在的研究认为树突棘的形态学改变与其骨架肌动蛋白的重构有密切关系。肌动蛋白以单体和多聚体两种表型存在,单体的肌动蛋白是由一条多肽链构成的球形分子,又称球状肌动蛋白(G-actin),肌动蛋白的多聚体形成肌动蛋白丝,称为纤维状肌动蛋白(F-actin)。肌动蛋白作为一种动态结构,持续进行组装和解聚,而体内肌动蛋白的装配及在细胞中的特性受肌动蛋白结合蛋白及上游信号分子的调节[3]。
Cofilin是中枢神经系统重要的肌动蛋白结合蛋白,Cofilin通过切断F-actin,增加F-actin末端肌动蛋白亚单位的解离速度来调节其动力学[11]。Cofilin的活性依赖于它的磷酸化状态,其受各种激酶及RhoGTPases的调节。RhoGTPase可通过 PAK、ROCK2/LIMK通路使cofilin磷酸化而降低其活性[12-13]。NMDA 受体激活后可以通过 NMDA/Ca2+/calcineurin/SSH通路使cofilin脱磷酸化而激活[14-15]。除了受磷酸化的调节之外,cofilin还与其它肌动蛋白结合蛋白发生相互作用,共同调节棘突的稳定性[16]。以上的研究提示,cofilin在树突棘可塑性中可能发挥关键作用。然而p-cofilin在吗啡依赖形成过程中的变化,国内外尚无报道。
本研究通过建立大鼠吗啡依赖模型,选择与吗啡依赖关系密切的脑区:海马、前额叶皮质、丘脑以及纹状体,测定了吗啡依赖状态下F-actin、G-actin、p-cofilin在上述脑区的表达变化。研究结果显示,在慢性吗啡处理2周和4周大鼠的前额叶皮质以及吗啡处理4周的海马中F-actin/G-actin的比值均出现明显上调。F-actin/G-actin比值的变化提示肌动蛋白骨架出现了重构,但是吗啡依赖不同时程其变化不同,不同的脑区其变化亦不相同。
本研究显示,肌动蛋白结合蛋白p-cofilin在吗啡处理2周和4周的海马和额叶皮质中均出现了表达上调,与F-actin的表达量增加相一致,提示吗啡诱导的肌动蛋白重构可能与cofilin的磷酸化状态有关。但p-cofilin在纹状体中的表达却减少了,提示不同的脑区可能存在着不同的调节机制。Cofilin的活性可影响神经元特异性肌动蛋白结合蛋白drebrin与F-actin的结合,进而影响棘突肌动蛋白网络的稳定性[16],而drebrin的异常可影响突触可塑性,导致机体不同程度的认知障碍[17]。行为学的研究表明cofilin与认知功能存在密切联系[18],在阿尔采末病病人大脑的额皮质和海马中,发现异常的cofilin-actin的棒状结构。由于应激诱导的这种棒状结构在轴突和树突的形成,能够导致神经性变性疾病[19]。在吗啡成瘾过程中均伴有不同程度的认知和行为障碍,提示p-cofilin吗啡依赖诱导的肌动蛋白重构过程中可能发挥重要作用,但其上游信号调节过程还有待于进一步研究。
总之,我们的研究首次证明,大鼠吗啡依赖时,伴随着脑组织中肌动蛋白水平的变化,提示吗啡依赖时大鼠脑组织中肌动蛋白发生了重构,肌动蛋白结合蛋白p-cofilin在吗啡诱导的肌动蛋白重构中可能发挥重要作用。我们的结果对进一步阐明吗啡成瘾的细胞及分子生物学机制提供了实验依据。
[1]Grewal S S,Fass D M,Yao H,et al.Calcium and cAMP signals differentially regulate cAMP-responsive element-binding protein function via a Rap1-extracellular signal-regulated kinase pathway[J].J Biol Chem,2000,275(44):34433 -41.
[2]Robinson T E,Kolb B.Structural plasticity associated with exposure to drugs of abuse[J].Neuropharmacology,2004,47(Suppl 1):33-46.
[3]Ethell I M,Pasquale E B.Molecular mechanisms of dendritic spine development and remodeling[J].Prog Neurobiol,2005,75(3):161-205.
[4]Pollard T D,Borisy G G.Cellular motility driven by assembly and disassembly of actin filaments[J].Cell,2003,112(4):453 -65.
[5]Toda S,Shen H W,Peters J,et al.Cocaine increases actin cycling:effects in the reinstatement model of drug seeking[J].J Neurosci,2006,26(5):1579 -87.
[6]Hou Y Y,Lu B,Li M,et al.Involvement of actin rearrangements within the amygdala and the dorsal hippocampus in aversive memories of drug withdrawal in acute morphine-dependent rats[J].J Neurosci,2009,29(39):12244 -54.
[7]Maldonado R,Koob G F.Destruction of the locus coeruleus decreases physical signs of opiate withdrawal[J].Brain Res,1993,605(1):128-38.
[8]温进坤,史建红,郑 斌,等.SM22α对血管平滑肌细胞肌动蛋白聚合和交联的调节[J].中国应用生理学杂志,2008,24(4):393-6.
[8]Wen J K,Shi J H,Zhang B,et al.The molecular mechanisms of SM22α in cytoskeleton remodeling of vascular smooth muscle cells[J].Chin Appl Physiol,2008,24(4):393 -6.
[9]Fukazawa Y,Saitoh Y,Ozawa F,et al.Hippocampal LTP is accompanied by enhanced F-actin content within the dendritic spine that is essential for late LTP maintenancein vivo[J].Neuron,2003,38(3):447 -60.
[10]Yuste R,Bonhoeffer T.Morphological changes in dendritic spines associated with long-term synaptic plasticity[J].Annu Rev Neurosci,2001,24:1071 -89.
[11]DesMarais V,Ghosh M,Eddy R,Condeelis J.Confilin takes the lead[J].J Cell Sci,2005,118(pt 1):19 -26.
[12]Asrar S,Meng Y,Zhou Z,et al.Regulation of hippocampal longterm potentiation by p-21-activated protein kinase1(PAK1)[J].Neuropharmacology,2009,56(1):73 -80.
[13]Zhou Z,Meng Y,Asrar S,et al.A critical role of Rho-kinase ROCK2 in the regulation of spine and synaptic function[J].Neuropharmacology,2009,56(1):81 -9.
[14]Carlisle H J,Manzerra P,Marcora E,Kennedy M B.SynGAP regulates steady-state and activity-dependent phosphorylation of cofilin[J].J Neurosci,2008,28(50):13673 -83.
[15]Wang Y,Shibasaki F,Mizuno K.Calcium signal-induced cofilin dephosphorylation is mediated by Alingshot via calcineurin[J].J Biol Chem,2005,280(13):12683 -9.
[16]Kojima N,Shirao T.Synaptic dysfunction and disruption of postsynaptic drebrin-actin complex:a study of neurological disorders accompanied by cognitive deficits[J].Neurosci Res,2007,58(1):1-5.
[17]贾丽洁,罗 艳,张富军,等.Drebrin对突触可塑性的影响以及认知功能障碍的研究进展[J].中国药理学通报,2010,26(8):989-92.
[17]Jia L J,Luo Y,Zhang F J,et al.Advance in research of the effects of Drebrin on synaptic plasticity and related cognitive dysfunction[J].Chin Pharmacol Bull,2010,26(8):989 -92.
[18]Rust M B,Gurniak C B,Renner M,et al.Learning,AMPA receptor mobility and synaptic plasticity depend on n-cofilin-mediated actin dynamics[J].EMBO J,2010,29(11):1889 -902.
[19]Peterson T S,Camden J M,Wang Y,et al.P2Y2 nucleotide receptor-mediated responses in brain cells[J].Mol Neurobio,2010,41(2-3):356-66.
