西葫芦株型性状主基因-多基因混合遗传分析
2010-08-10陈凤真何启伟
陈凤真 何启伟
(1菏泽学院园林工程系,山东菏泽 274000;2山东省农业科学院蔬菜研究所,山东济南 250100)
西葫芦,即美洲南瓜(Cucurbita pepoL.)是冬春设施栽培的主要蔬菜作物之一。西葫芦从生长习性来分,主要有蔓生、矮生和半蔓生(Kasrawi,1995)。株高和叶片数是西葫芦重要的农艺性状,它们与西葫芦的产量及熟性性状有密切的相关性,研究它们的基因效应,对西葫芦的株型、产量及熟性性状的遗传改良有重要的指导意义(陈凤真,2008)。陈凤真等(2007)采用经典遗传学对西葫芦株高和叶片数进行了遗传分析。经典遗传学虽然能估计基因总体效应,但不能区分主基因、多基因效应。
本试验在李建友(2004)研究的基础上,利用盖钧镒(2005)的主基因-多基因混合分析方法,对西葫芦株型性状的遗传开展进一步的研究,旨在对西葫芦杂种优势产生的机理进行研究,并为开展西葫芦的株型育种提供参考依据。
1 材料与方法
1.1 试验材料
选用蔓生和矮生两种不同株型,果形差异较大且经6代以上自交选育的3个西葫芦自交系q-1(蔓生、白皮、长棒形)、23-4G(矮生、墨绿皮、粗短形)和A-7(矮生、深绿皮、粗短形)为试材。其中q-1和A-7分别来自奇台和阿尔及利亚的引进品种,23-4G由双丰特早自交分离选育而来。
1.2 试验方法
试验在山东农业大学蔬菜试验站进行。2005年春配制F1,组合1:q-1(♀)×23-4G(♂),组合2:q-1(♀)×A-7(♂)。2005年10月在日光温室中获得F2、回交世代BC1(F1×q-1)和BC2(F1×23-4G 和 F1×A-7)。
2006年3月12日28 ℃恒温催芽,3月16日播种,3月30日定植到拱圆大棚(长60 m,跨度10 m)。亲本及F1各定植30株,回交世代BC1、BC2分别定植60株,F2定植90株,行距60 cm,株距50 cm;3次重复,随机区组设计。单干整枝,落蔓管理,其他管理同一般生产。拔秧时测量株高及统计叶片数,并对数据进行统计分析。
1.3 统计分析方法
采用植物数量性状主基因-多基因混合遗传模型(盖钧镒,2005)对西葫芦6个世代的株型性状进行联合分析,通过极大似然法(MaximuMlikelihood method)和 IECM算法(Iterated expectation and conditional maximization)对混合分布中的有关成分分布参数做出估计;然后利用AIC准则(Akaike’s information criterion)选择AIC值最小或较小的1个或几个备选模型,再通过一组适合性测验,共有5个统计量,即U12、U22、U32、nW2和Dn,其中U12、U22、U32为均匀性检验,nW2为Smirnov检验,Dn为Kolmogorov检验,根据结果选择最优模型,即选择统计量达到显著水平个数最少的模型作为最优模型;并估计主基因和多基因效应值、主基因遗传率等遗传参数。分析软件由南京农业大学国家大豆改良中心提供。
2 结果与分析
2.1 株高和叶片数测定值的次数分布
将各个世代的株高和叶片数次数分布分别列于表1、2。从表1、2可知,2个组合F1的株高性状存在杂种优势,2个组合的F1和F2叶片数的平均数低于低值亲本的平均数,说明西葫芦叶片数存在负向超亲优势。2个组合的F2株高均集中在150~350 cm之间,叶片数均集中在41~61片之间。由图1~4可以看出,2个组合的F2株高和叶片数都呈单峰较明显的偏态分布,表明主基因可能控制2个组合株高和叶片数的遗传。
2.2 株型性状遗传模型
以2个组合的6个不同世代材料株高和叶片数观测值为基础,利用植物数量性状主基因-多基因混合遗传模型的多世代联合分析方法对6个世代群体进行分析,计算不同遗传模型的极大对数似然值和AIC值(表3)。
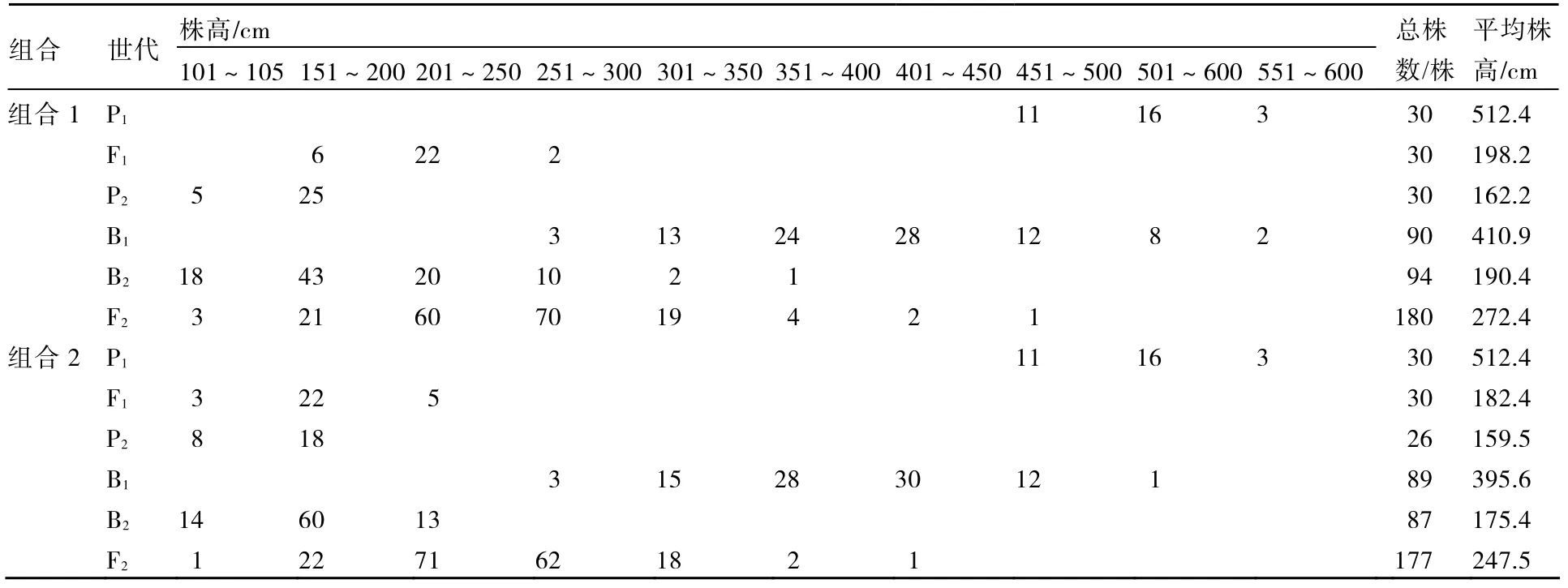
表1 各世代株高表型值次数分布

表2 各世代叶片数表型值次数分布

图1 组合1的F2株高次数分布

图2 组合2的F2株高次数分布
根据AIC准则,组合1的株高B-1模型和E模型AIC值较小,B-1和E模型可作为备选模型;组合2的株高B-1和E-1模型AIC值较小,B-1和E-1模型可作为备选模型;组合1与组合2的叶片数B-1模型AIC值最小,组合1叶片数的D-4模型和组合2叶片数的D-2、D-4模型AIC值与最小值接近,可作为备选模型。
进一步对株高和叶片数性状的B-1模型与其他备选模型进行适合性检验,选择统计量达到显著水平个数较少的模型作为最优模型。表4、5结果表明:在6个世代的30个统计量中,组合1的株高B-1模型有5个统计量达到显著水平,而E模型有15个统计量达到显著水平;组合1的叶片数B-1模型达到显著水平的统计量个数少于D-4模型;组合2的株高B-1模型的各统计量不显著,叶片数B-1模型有1个统计量达到显著水平,而株高和叶片数的其他备选模型均有10个以上的统计量达到显著水平;因此2个组合的株高和叶片数B-1模型不仅最佳而且最适。
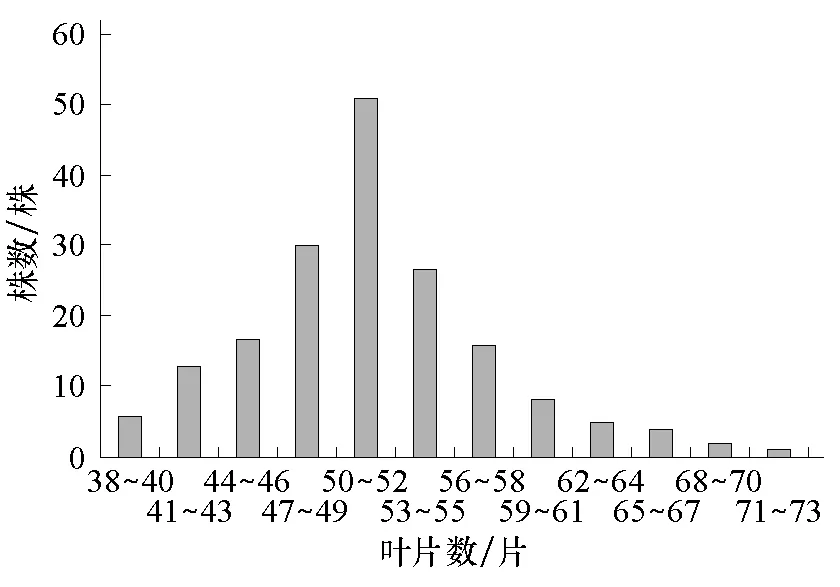
图3 组合1的F2叶片数次数分布
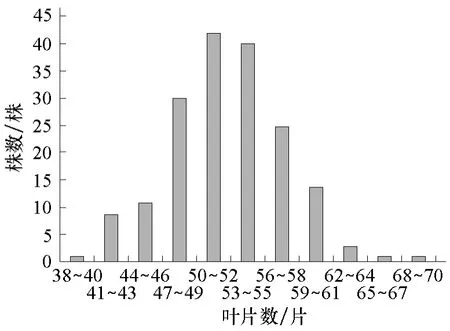
图4 组合2的F2叶片数次数分布
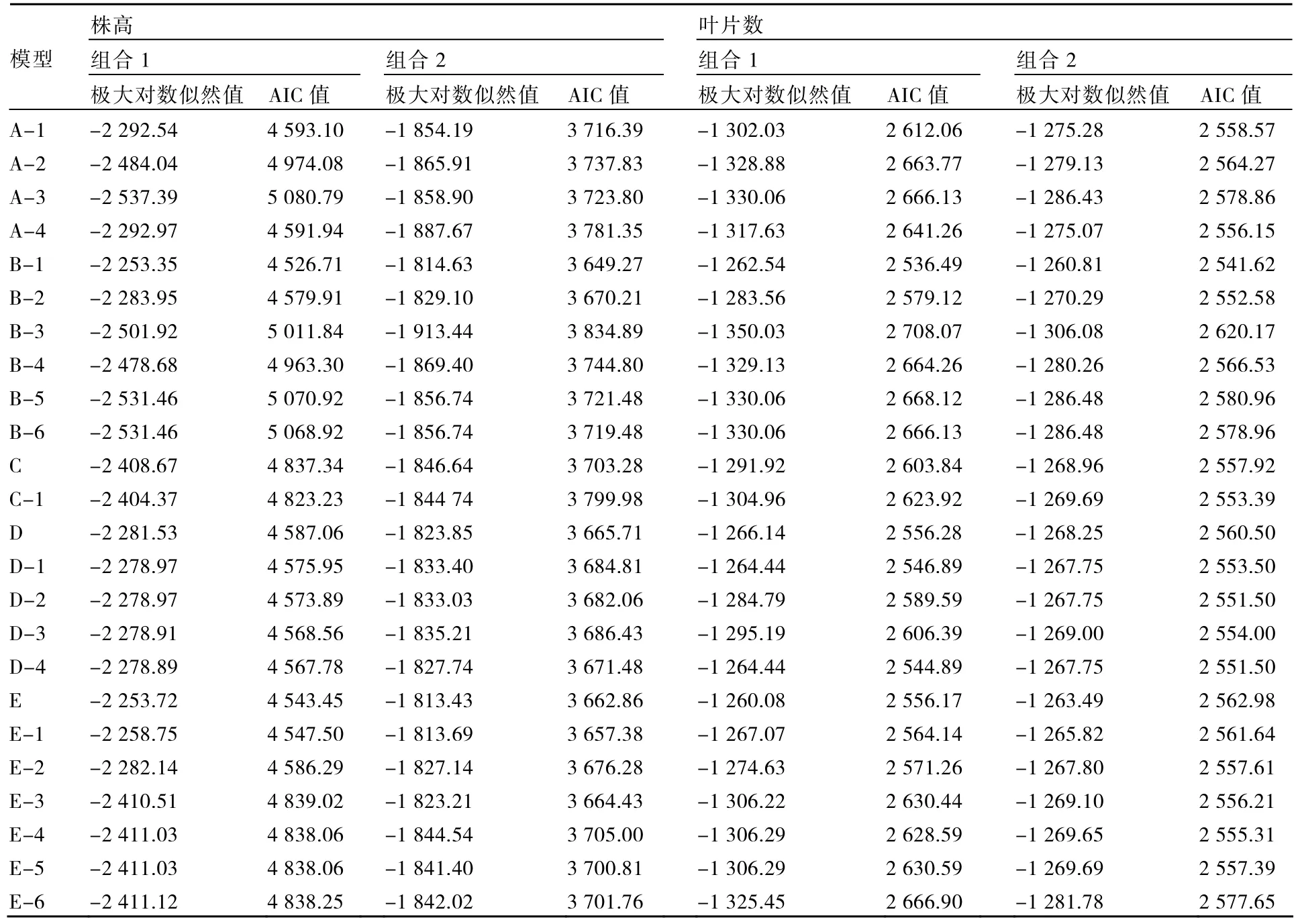
表3 株高和叶片数模型的极大对数似然值和AIC值
2.3 株型性状遗传参数估计
2.3.1 株高遗传参数的估计 各组合株高性状遗传参数估计值列于表6。1阶遗传参数结果表明:组合1和组合2两对主基因显性效应值之和(ha+hb)分别为-93.60和-8.01;组合1与组合2的两对主基因加性效应值之和(da+db)分别为151.89和 180.76;加性×加性上位性互作效应(i)分别为34.77和13.70,加性×显性上位性互作效应之和(jab+jba)分别为13.71和8.67;2个组合的两对主基因显性效应值之和(ha+hb)均为负值,且小于两对主基因加性效应值之和,故F1株高性状无超亲优势;因此西葫芦的株高性状均以加性效应为主,其次均为加性×加性上位性互作效应;由表6中的2阶参数估计值可知:组合1与组合2的分离后代的差异均是由主基因引起的,2个组合的F2株高的主基因遗传率分别为67.00 %和74.21 %,分别还有33.00 %和25.79 %的变异是由环境因素决定的,说明环境对西葫芦株高性状有一定的影响,宜早代选择。

表4 株高的适合性检验

表5 叶片数的适合性检验
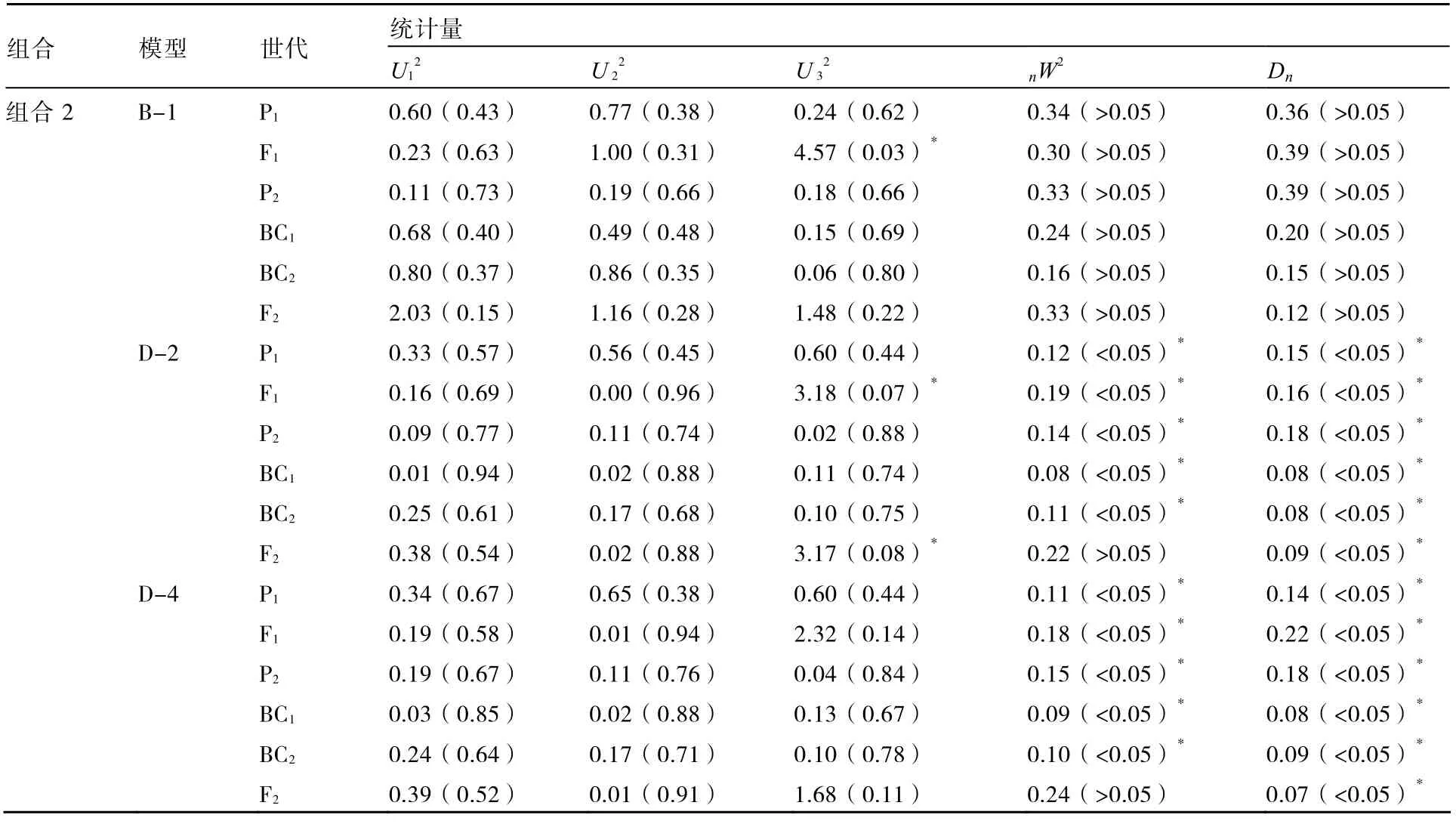
续表

表6 株高、叶片数的各遗传参数估计值
2.3.2 叶片数性状遗传参数估计 将2个组合叶片数的适合模型下遗传参数估计值列入表6。1阶遗传参数结果表明:组合1和组合2的主基因显性效应值之和(ha+hb)分别为-2.04和-0.84,两对主基因加性效应值之和(da+db)分别为-1.20和-0.63,加性×加性上位性(i)分别为-6.13和-1.52、加性×显性上位性互作效应之和(jab+jba)分别为-8.89和-2.03,显性×显性上位性互作效应值(l)分别为-6.46和-1.31;2个组合的主基因显性效应值之和为负值,说明F1叶片数性状减少,表现负向超亲优势;在遗传效应值中加性效应值之和较大,因此2个组合的叶片数性状以加性和显性效应为主。2阶遗传参数结果表明:组合1和组合2叶片数性状的遗传均由两对主基因控制,组合1主基因在分离后代BC1、BC2与F2的遗传率分别为:66.53 %、62.74 %和68.07 %;组合2的 BC1、BC2与F2的主基因遗传率分别为:63.67 %、60.89 %和71.96 %,说明环境对西葫芦叶片数性状有一定的影响,2个组合的F2叶片数的遗传率分别有31.93 %和28.04 %的变异是由环境因素决定的,因此对叶片数的选择也宜在早代。
3 结论与讨论
西葫芦良好的株型性状是实现设施栽培高产、早熟和优质理想品种的必要条件。西葫芦株型育种的目的是选择株型紧凑的品种,本研究运用多世代联合分析方法,对西葫芦株型性状进行了遗传分析,并为西葫芦育种提供了依据。
许多学者采用主基因-多基因混合遗传分析方法对蔬菜的株高及叶片数进行了研究。辣椒和普通白菜(小白菜)的株高分别符合1对加性主基因+加性-显性多基因模型与1对负向完全显性主基因+加性-显性多基因遗传模型(陈学军和陈劲枫,2006;韩建明 等,2008);而黄瓜株高受1对加性-显性主基因+加性-显性多基因共同控制,以主基因加性效应为主(嵇怡 等,2009);刘二艳等(2009)对青花菜花球荚叶性状进行了遗传分析,结果表明不同的群体同一性状的遗传模型不同。可见不同蔬菜株高和叶片数的遗传模型并不一致,但其株高和叶片数均以主基因控制为主;而且不同植物或同一种植物不同基因型株高性状的遗传模型也不一致,可见材料的不同基因型对同一性状的遗传有着重要的影响;此外,环境对株高和叶片数有不同的影响。
本研究明确了西葫芦株高和叶片数性状受加性-显性-上位性两对主基因(B-1)遗传模型控制,株高以基因的加性效应为主,叶片数以加性效应和显性效应为主;F2的主基因遗传率较高,环境影响相对较小,表明西葫芦株型育种宜早代选择。就丰产而言,株型紧凑的西葫芦品种更高产(陈凤真,2008)。因此,对西葫芦株型性状的遗传改良,可选择植株矮小、叶片数少、节间距短的亲本材料,通过杂交、回交转移主基因,选育株型紧凑的材料。
陈凤真,何启伟,樊治成,盛金.2007.西葫芦8个农艺性状的遗传效应分析.园艺学报,34(5):1183-1188.
陈凤真.2008.西葫芦农艺性状的遗传、遗传图谱的构建及遗传多样性的研究〔博士论文〕.泰安:山东农业大学.
陈学军,陈劲枫.2006.辣椒株高遗传分析.西北植物学报,26(7):1342-1345.
盖钧镒.2005.植物数量性状遗传体系的分离分析方法研究.遗传,27(1):130-136.
韩建明,侯喜林,史公军,陈沁滨.2008.不结球白菜株高性状主基因+多基因遗传分析.南京农业大学学报,31(1):23-26.
嵇怡,徐强,陈学好.2009.黄瓜株高性状遗传模型分析.扬州大学学报:农业与生命科学版,30(3):75-79.
李建友.2004.西葫芦主要性状杂种优势及其形成机理的研究〔硕士论文〕.泰安:山东农业大学.
刘二艳,刘玉梅,方智远,杨丽梅,庄木,张扬勇,袁素霞,孙继峰,李占省,孙培田.2009.青花菜花球‘荚叶’性状主基因+多基因遗传分析.园艺学报,36(11):1611-1618.
Kasrawi MA.1995.Diversity in landraces of summer squash froMJordan.Genetic Resources and Crop Evolution,42:223-230.
