紫果西番莲顶枯病病原菌生物学特性分析及室内杀菌剂毒力测定
2023-09-26黄艳花崔忠吉蒙成欧善生钟琪斌
黄艳花 崔忠吉 蒙成 欧善生 钟琪斌
(广西农业职业技术大学,广西 南宁 530007 )
紫果西番莲,学名鸡蛋果(Passiflora edulia Sims),又名百香果,是西番莲科西番莲属的草质藤本植物。原产安的列斯群岛,广植于热带和亚热带地区。植株寿命约20 年,经济寿命约8~10 年。花期6 月,果期11 月。果可生食或作蔬菜、饲料。入药具有兴奋、强壮之效。果瓤多汁液,加入重碳酸钙和糖,可制成芳香可口的饮料,还可用来添加在其他饮料中以提高饮料的品质。种子榨油,可供食用和制皂、制油漆等。花大而美丽,没有香味,可作庭园观赏植物。紫果西番莲具有“果汁之王”“摇钱树”等美称。[1]因其种植周期短、产生效益快,近年来被选择作为广西部分地区乡村振兴产业进行开发。随着种植规模不断扩大,紫果西番莲茎基腐病、炭疽病、病毒病和蚜虫等病虫危害也日趋严重,加之管理不当、技术欠缺,已严重影响紫果西番莲 产业健康发展。紫果西番莲顶枯病病原菌为暹罗炭疽菌 (Colletotrichum siamense)。发病初期引起西番莲顶芽坏死,病斑逐渐由顶端向下扩展,枝条由顶端向下干枯坏死,严重时整株枯死。该病害在广西11 月发病较重,最高发病率可达49.5%,造成大面积死芽枯枝,严重影响产量。[2]紫果西番莲顶枯病已对紫果西番莲产业可持续发展造成严重威胁,因此迫切需要掌握其病原菌生物学特性,并筛选出高效防治杀菌剂,为大面积科学防控提供参考依据。
暹罗炭疽菌是胶孢炭疽菌(C.gloeosporioides)复合物的物种之一[3],其可与其他炭疽菌混合引发鹅掌柴、荔枝、油茶、南天竹、紫山药、红瑞木和洒金珊瑚等多种植物炭疽病[4-10],也可独立引发多穗柯、棕竹、杨桃、菠萝蜜、百日草、大叶黄杨、红肉苹果等多种植物炭疽病[3,11-16]及多肉植物青星美人黑腐病[17]。张艳婷[18]研究认为,引起草莓茎基腐病的主要病原菌为暹罗炭疽菌;解小锋等[19]研究明确引起枣炭疽病的病原菌为暹罗炭疽菌;徐丹丹等[20]认为,广东咖啡炭疽病的优势病原菌为果生炭疽菌(C. fructicola)和暹罗炭疽菌。在暹罗炭疽菌生物学特性研究方面,罗敦文[21]研究发现,菠萝蜜蒂腐病菌(C. siamense)菌丝生长适宜环境条件为:PDA 和查彼培养基,温度28℃,pH 值7.0,碳源为葡萄糖、阿拉伯糖、麦芽糖和果糖,氮源为酵母浸粉和牛肉浸膏,病原菌菌丝致死温度为59℃处理10 min;刘在哲等[16]研究认为,红肉苹果炭疽病菌菌丝生长适宜环境条件为:PSA和SDA 培养基,温度25~30℃,pH 值8.0~10.0,持续光照或黑暗交替;王芳等[17]研究表明,最适青星美人黑腐病菌菌丝生长温度为30℃,pH 值为5.0,碳源为麦芽糖,氮源为酵母浸膏,光暗交替,最适病原产孢条件温度为30℃,pH 值为7.0,碳源为麦芽糖,氮源为蛋白胨,连续黑暗。在药剂防治研究方面,宋慧云等[11]研究表明,98.4%多菌灵、97%吡唑醚菌酯和98%福美双可作为防治多穗柯炭疽病的化学杀菌剂;罗敦文[21]研究认为,50%咪鲜胺锰盐对菠萝蜜蒂腐病病原菌抑制作用较强;王志华等[22]研究指出,多菌灵可湿性粉剂和苯醚甲环唑水分散粒剂对金森女贞炭疽病的抑制作用最强。紫果西番莲顶枯病是2022 年国内首次报道的新病害[2],目前针对其生物学特性及室内防治药剂筛选的研究尚无文献报道。本文采用菌丝生长速率法和孢子计数法分析不同培养条件对紫果西番莲顶枯病病原菌菌丝生长和产孢量的影响,以菌丝生长速率法对12 种杀菌剂进行室内毒力测定,旨在掌握该病原菌生长条件并筛选出防治高效的化学杀菌剂,为紫果西番莲顶枯病防控提供科学依据。
1 材料与方法
1.1 试验材料
供试菌株:紫果西番莲顶枯病病原菌暹罗炭疽菌(C.siamense),由广西农业职业技术大学植物病理学实验室分离、鉴定及保藏。
供试杀菌剂:供试药剂12 种,分别为咪唑类咪鲜胺(450 g/L 水乳剂,上海沪联生物药业(夏邑)股份有限公司)、甲基硫菌灵(70%可湿性粉剂,江苏龙灯化学有限公司)、抑霉唑(20%水乳剂,一凡生物科技集团有限公司);三唑类丙环唑(25%乳油,招远三联化工厂有限公司)、苯醚甲环唑(10%水分散粒剂,先正达南通作物保护有限公司);苯并咪唑衍生物类多菌灵(50%可湿性粉剂,四川润尔科技有限公司);有机硫类杀菌剂乙蒜素(80%油剂,南阳新卧龙生物化工有限公司);吡咯类杀菌剂腈菌唑(40%悬浮剂,一凡生物科技集团有限公司);取代苯类杀菌剂百菌清(75%可湿性粉剂,山东大成生物化工有限公司);三唑类与甲氧基丙烯酸酯类复配的苯甲·吡唑酯(40%悬浮剂,江苏云农化工有限公司);三唑类与咪唑类复配的苯甲·咪鲜胺(20%微乳剂,山东中新农生物科技有限公司);甲氧基丙烯酸酯类与三唑类复配的肟菌·戊唑醇(75%水分散粒剂,山东荣邦化工有限公司)。
供试仪器:供试培养箱为韶关市泰宏医疗器械有限公司生产的珠江牌生化培养箱,型号:LRH-250 A。
1.2 病原菌生物学特性分析
1.2.1 病原菌菌丝生长、产孢及孢子萌发对不同pH的适应性
参照闪瑶等[23]方法并略加修改,分别配置1.0 mol/L HCl、1.0 mol/L NaOH 和PDA 培养基并灭菌,在无菌条件下用HCl 和NaOH 把PDA 培养基的pH 分别调为2.0、3.0、4.0、5.0、6.0、7.0、8.0、9.0、10.0、11.0、12.0 共11 个梯度,倒入培养基平板,待冷却后将直径6.0 mm 的病原菌菌饼接种于不同pH 值平板中央,每处理4 次重复,28℃黑暗培养。4 d 后采用十字交叉法测量菌落直径,10 d 后每皿用10 mL 无菌水配制孢子悬浮液,用血球计数板统计产孢量[24](下同)。用上述配置已灭菌好的HCl 和NaOH 在无菌条件下将无菌水的pH 值调至2.0~12.0,分别用不同pH 值的无菌水加在相同条件下培养10 d的病原菌菌落配制成孢子悬浮液,用不同pH 值的孢子悬浮液涂抹在相应pH 值的PDA 培养基平板上,置28℃恒温培养7 h,显微镜下观察孢子萌发情况,芽管长度达分生孢子、宽度计为萌发,每处理连续统计孢子100 个以上,4 次重复,用计数器统计分生孢子在上述不同pH 值条件下的萌发率。
1.2.2 病原菌菌丝生长、产孢及孢子萌发对不同温度的适应性
参照梁萍等[24]方法并略加修改,在PDA 平板上培养4 d 的菌落边缘用直径为6 mm 的打孔器取菌饼,将菌饼接种于PDA 培养基中央,分别置于不同温度(4℃、8℃、12℃、16℃、20℃、24℃、28℃、32℃、36℃、40℃)恒温培养箱中培养,每个温度4 次重复。参照1.2.1 方法测量菌落直径及产孢量;参照方中达[25]方法采用琼胶平板表面萌发法测定孢子萌发率,用灭菌水将孢子洗刷下来后,调制成浓度为5.00×106个/mL 孢子悬浮液,把孢子悬浮液均匀涂抹在水琼脂培养基平板上[18],置28℃条件培养7 h,显微镜下观察孢子萌发情况,芽管长度达分生孢子宽度计为萌发,每处理连续统计孢子100 个以上,用计数器统计萌发率,每处理4 次重复(下同)。
1.2.3 病原菌菌丝生长、产孢对不同光照的适应性
参照蒙成等[26]方法并略加修改,将6 mm 病原菌菌饼分别接种于PDA 培养基平板中央,进行全暗(0 lx)、12 h 光暗交替(150 lx)、12 h 光暗交替(200 lx)、12 h 光暗交替(300 lx)、连续光照(300 lx)5 种光照环境处理。全黑暗处理使用4 层锡箔纸包裹遮光。每处理4 次重复,置28℃人工气候箱培养4 d,测量菌落直径,培养10 d 测定产孢量。
1.2.4 病原菌菌丝生长、产孢对不同碳源的适应性
参照闪瑶等[23]方法并略加修改,用等质量葡萄糖、山梨醇、麦芽糖、甘露醇、乳糖、可溶性淀粉、D-木糖、甘油8 种碳源分别置换Czapek 培养基(硝酸钠3.0 g、磷酸氢二钾1.0 g、硫酸镁0.5 g、氯化钾0.5 g、硫酸铁0.01 g、蔗糖30.0 g、琼脂20.0 g、水1000.0 mL)中的蔗糖,配制成不同碳源培养基。设无碳源培养基为对照。将6 mm 病原菌菌饼分别接种于上述培养基平板中央,每处理4 次重复,置28℃培养箱培养4 d 测量菌落直径,观察记录菌落形态特征。10 d 计算产孢量。
1.2.5 病原菌菌丝生长、产孢对不同氮源的适应性
参照闪瑶等[23]方法并略加修改,用等质量酵母粉、牛肉浸膏、甘氨酸、蛋白胨、L-精氨酸、L-丙氨酸、尿素、L-赖氨酸8 种氮源分别代替Czapek培养基的硝酸钠,配制不同氮源培养基。设无氮源培养基为对照。将6 mm 病原菌菌饼分别接种于上述培养基平板中央,每处理4 次重复,置28℃培养箱培养4 d 测量菌落直径,观察记录菌落形态特征。10 d 计算产孢量。
1.2.6 病原菌菌丝生长、产孢对不同培养基的适应性
参照梁萍等[24]方法并略加修改,制备6 种培养基。马铃薯葡萄糖培养基(PDA):马铃薯200.0 g、葡萄糖20.0 g、琼脂20.0 g、水1000.0 mL;马铃薯蔗糖培养基(PSA):马铃薯200.0 g、蔗糖20.0 g、琼脂20.0 g、水1000.0 mL。玉米粉培养基(CMA):玉米粉30 g,琼脂20 g,水1000 mL。汁液培养基(西番莲汁液):健康新鲜西番莲枝叶200.0 g,水熬煮30 min,琼脂20.0 g,水1000.0 mL。燕麦培养基(OA):燕麦片30.0 g、琼脂20.0 g、水1000.0 mL);琼脂培养基(WA):琼脂15.0 g,水1000.0 mL。6 种培养基pH 自然。将6 mm 病原菌菌饼分别接种于上述培养基平板中央,每处理4 次重复,置28℃培养箱培养4 d 测量菌落直径,观察记录菌落形态特征。10 d 计算产孢量。
1.3 防治杀菌剂室内毒力测定
参照刘俏等[27]方法并略加修改,采用菌丝生长速率法测定杀菌剂对病原菌抑制效果。经3 次预试验,确定各杀菌剂的梯度浓度。在无菌操作条件下,将12 种杀菌剂按照倍数法稀释成不同浓度母液;再从每个倍数的母液中吸取1.0 mL 药液加入至99.0 mL 的55℃、无菌PDA 培养基中制作成含药培养基。杀菌剂终浓度见表1。以不加杀菌剂的PDA培养基作为对照。充分摇匀后倒入直径9 cm 培养皿中,每皿15.0 mL。待皿内培养基凝固后,在其中央接种直径6 mm 的供试病原菌菌饼。每处理4 次重复。28℃培养箱暗培养4 d 后采用十字交叉法测量菌落直径,计算各杀菌剂对菌丝体生长的相对抑制率。
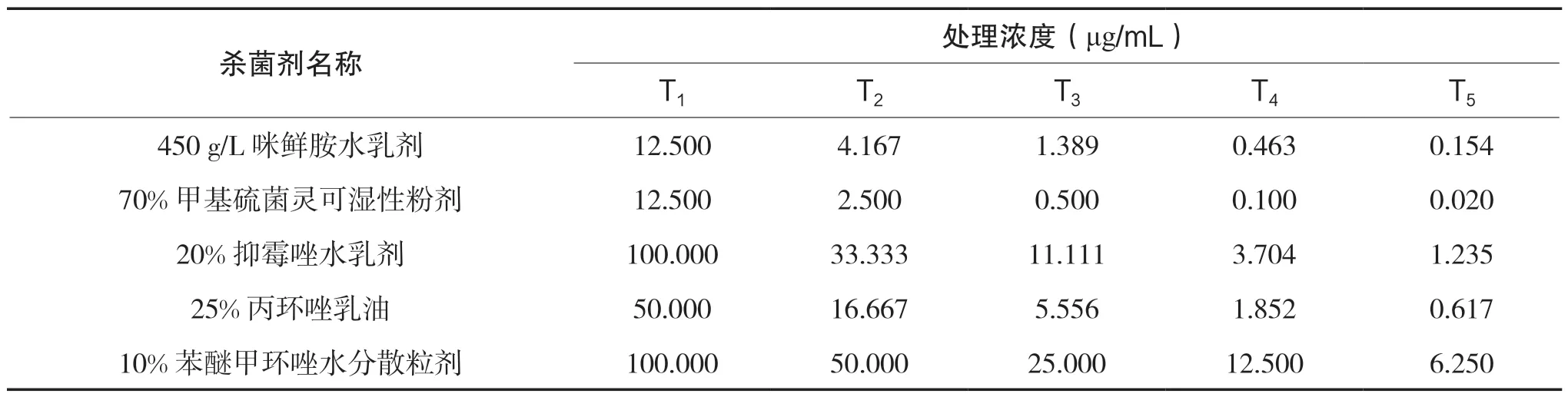
表1 供试杀菌剂及其处理浓度
菌落直径=测定菌落直径-菌饼直径
相对抑制率(%)=(对照菌落直径-处理菌落直径)/对照菌落直径×100%
采用Excel 2010、DPS 7.0 和SPASS 16.0 进行统计分析。将杀菌剂质量浓度(μg/mL)转换成以10为底的对数值(x),菌丝生长相对抑制率换算成几率值(y),以浓度对数值为横坐标,以相对抑制率几率值为纵坐标,作毒力回归直线,计算毒力回归方程(y=ax +b)和相关系数(r), 根据回归方程计算出抑制中浓度EC50值。毒力回归方程能有效反应该药剂质量浓度与抑菌效果的关系,EC50即抑制或杀死50%的目标菌所需要的杀菌剂浓度,是衡量杀菌剂毒力的最可靠标准,用EC50值可以对不同种类的杀菌剂毒力大小进行互相比较。[28]
2 结果与分析
2.1 紫果西番莲顶枯病病原菌生物学特性
2.1.1 病原菌菌丝生长、产孢及孢子萌发对不同pH值的适应性
紫果西番莲顶枯病病原菌暹罗炭疽菌菌丝在值为2.0 时不生长;pH 值为3.0~12 时均能生长;在pH 值为5.0~9.0 时生长较快,菌落直径达77.38~83.50 mm。各处理间差异不显著(P>0.05,下同),但均极显著高于其他pH 值条件下的菌落直径(P<0.01,下同),其中pH 值为6.0 时菌落直径最大,为83.50 mm,且显著高于pH 值为5.0、7.0和8.0 的菌落直径(P<0.05,下同),但与pH 值为9.0 时的菌落直径差异不显著。pH 值为9.0 时的菌落直径与pH 值为5.0、7.0 和8.0 的菌落直径也无显著差异。在pH 值为4.0 时菌丝生长较快,其菌落直径为67.00 mm,极显著高于pH 值为11.0、12.0和3.0 时的菌落直径。可见,紫果西番莲顶枯病病原菌暹罗炭疽菌菌丝在pH 值为3.0~12.0 均能生长,以pH 值为5.0~9.0 为菌丝生长的较佳范围(图1)。

图1 病原菌菌丝生长、产孢及孢子萌发对不同pH 的适应性
病原菌在pH 值为2.0 时不能产孢;在pH 值为7.0 和8.0 时产孢量较多,产孢量分别为53.72×104个/mL 和54.32×104个/mL,二者间差异不显著,但均显著高于其他pH 值条件下的产孢量;在pH 值为9.0 时产孢量也较多,且显著高于除pH 值为7.0和8.0 外其他pH 值条件下的产孢量。说明紫果西番莲顶枯病病原菌暹罗炭疽菌在pH 值为3.0~12.0均能产孢,而pH 值7.0 和8.0 为较佳产孢量范围(图1)。
病原菌孢子在pH 值为2.0 时萌发受到严重抑制,孢子萌发率为0.07%。在pH 值为5.0~10.0 时孢子萌发率较高,为90.20%~93.13%,处理间差异不显著,但均显著高于其他pH 值条件下的孢子萌发率。在pH 值为11.0 时孢子萌发率也较高,且显著高于pH 值为12.0、4.0、3.0 和2.0 条件下的孢子萌发率。说明紫果西番莲顶枯病病原菌暹罗炭疽菌孢子在pH 值为2.0~12.0 时均能萌发,而pH 值为5.0~10.0 为较佳孢子萌发范围(图1)。
2.1.2 病原菌菌丝生长、产孢及孢子萌发对不同温度的适应性
暹罗炭疽菌菌丝在温度为4℃、40℃时不生长,8℃时生长严重受阻,12~36℃时均能生长,8~28℃时生长速度与温度度数成正比,28~36℃时生长速度与温度度数成反比。28℃时生长较快,菌落直径为88.00 mm,极显著高于其他温度条件下的菌落直径;24℃时菌落直径为82.00 mm,极显著高于除28℃外其他温度条件下的菌落直径。可见,紫果西番莲顶枯病病原菌暹罗炭疽菌菌丝在温度为12~36℃均能生长,以温度28℃为菌丝生长较佳温度(图2)。

图2 病原菌菌丝生长、产孢及孢子萌发对不同温度的适应性
病原菌在温度为8℃、40℃时不产孢,12~36℃时均能产孢,28℃时病原菌产孢量较多,产孢量为53.72×104个/mL,极显著高于其他温度条件下的产孢量;32℃、24℃、36℃时产孢量分别为24.84×104个/mL、20.40 ×104个/mL、13.75×104个/mL;32℃与24℃时的产胞量差异不显著,与36℃时产孢量有显著性差异;24℃与36℃时产孢量无显著性差异,三者均显著高于除28℃外其他温度条件下的产孢量。可见,紫果西番莲顶枯病病原菌暹罗炭疽菌在温度为12~36℃时均能产孢,较佳产孢温度为28℃(图2)。
病原菌孢子在温度为8℃时不能萌发;12~40℃时均能萌发;20~36℃时病原菌孢子萌发率较高,萌发率为91.52%~97.08%,极显著高于其他温度条件下的萌发率;16℃时病原菌孢子萌发率为73.44%,极显著高于温度为12℃和40℃时的孢子萌发率。说明紫果西番莲顶枯病病原菌暹罗炭疽菌孢子在温度为12~40℃时均能萌发,20~36℃为较佳孢子萌发范围(图2)。
2.1.3 病原菌菌丝生长、产孢对不同光照的适应性
从表2 看出,暹罗炭疽菌菌丝生长对光照要求不严格。5 种光照处理均能生长,完全黑暗对菌丝生长有利,光照对菌丝生长有一定抑制作用。其中,菌丝在连续黑暗条件下生长较快,菌落直径为84.50 mm,显著高于其他光照条件下的菌落直径。12 h 亮12 h 暗(150 lx )、12 h 亮12 h 暗(200 lx )、12 h 亮12 h 暗(300 lx )及连续光照(300 lx )条件下菌落直径没有显著差异(表2)。

表2 病原菌菌丝生长、产孢对不同光照的适应性
病原菌在5 种光照处理条件下均能产孢,完全黑暗对病原菌产孢有利,光照对病原菌产孢有一定抑制作用。其中,病原菌在连续黑暗条件下产孢量较大,产孢量为37.66×104个/mL,极显著高于其他光照条件下的产孢量。12 h 亮12 h 暗(150 lx)、12 h 亮12 h 暗(200 lx )、12 h 亮12 h 暗(300 lx )及连续光照(300 lx )条件下,病原菌产孢量没有显著差异(表2)。
综上所述,最适合暹罗炭疽菌菌丝生长及产孢的光照条件为连续黑暗。
2.1.4 病原菌菌丝生长、产孢对不同碳源的适应性
暹罗炭疽菌在供试的9 种碳源上均能生长,但菌丝生长速度在不同碳源间存在差异。其中,以葡萄糖为碳源时菌丝生长最快,菌丝生长密厚,菌落直径达78.67 mm,显著高于其他碳源的菌落直径。菌丝生长较快的碳源依次为山梨醇、麦芽糖、甘露醇,其菌丝较稀疏,菌落直径在69.50~70.83 mm,相互间无显著差异。以D-木糖和甘油为碳源时菌丝生长较慢,菌落直径均低于61.00 mm,相互间无显著差异,但均与以山梨醇、麦芽糖、甘露醇为碳源的菌落直径存在显著或极显著差异。说明葡萄糖是最适宜暹罗炭疽菌菌丝生长的碳源(表3)。

表3 病原菌菌丝生长、产孢对不同碳源的适应性
暹罗炭疽菌产孢量最多碳源依次为乳糖、D-木糖、山梨醇,产孢量分别为46.45×104个/mL、37.36×104个/mL、29.89×104个/mL。乳糖与D-木糖产孢量没有显著差异,与山梨醇产孢量有显著差异。D-木糖与山梨醇产孢量没有显著差异,三者均极显著高于其他碳源条件下的产孢量。产孢量较多的碳源依次为淀粉、甘露醇,产孢量分别为19.19×104个/mL、13.53×104个/mL,相互间无显著差异。以麦芽糖为碳源时产孢量最少,与淀粉、甘露醇为碳源产孢量差异显著。说明乳糖、D-木糖、山梨醇是最适合暹罗炭疽菌产孢的碳源(表3)。
2.1.5 病原菌菌丝生长、产孢对不同氮源的适应性
暹罗炭疽菌在供试的9 种氮源上均能生长,但菌丝生长速度在不同氮源间存在差异。其中,以酵母粉、牛肉浸膏为氮源时菌丝生长最快,菌落直径达89.13 mm 和87.75 mm,酵母粉菌丝密厚,牛肉浸膏菌丝稀松,两者间没有显著差异,但均显著高于其他氮源的菌落直径。菌丝生长较快的氮源依次为甘氨酸、蛋白胨,甘氨酸菌丝密厚,蛋白胨菌丝稀松,菌落直径分别为73.88 mm 和72.00 mm, 相互间无显著差异。以L-丙氨酸、尿素、L-赖 氨酸为氮源时菌丝生长较慢,菌落直径均低于65.50 mm,相互间无显著差异,但均与以甘氨酸、 蛋白胨为氮源的菌落直径存在显著或极显著差异。 说明酵母粉、牛肉浸膏是最适宜暹罗炭疽菌菌丝生 长的氮源(表4)。暹罗炭疽菌在供试的9 种氮源上除尿素外均能产孢,尿素可抑制孢子产生。产孢量最多的氮源为牛肉浸膏,产孢量为252.03×104个/mL,极显著高于其他氮源条件下的产孢量。产孢量较多的氮源为蛋白胨、酵母粉,产孢量分别为55.94×104个/mL、47.66×104个/mL,相互间无显著差异,但极显著高于除牛肉浸膏为氮源外的其他氮源条件下的产孢量。以甘氨酸、L-精氨酸、L-丙氨酸为氮源时产孢量最少,相互间产孢量没有显著性差异。说明牛肉浸膏是最适合暹罗炭疽菌产孢的氮源(表4)。

表4 病原菌菌丝生长、产孢对不同氮源的适应性
2.1.6 病原菌菌丝生长、产孢对不同培养基的适应性
暹罗炭疽菌在6 种培养基中均能生长。其中,PDA 培养基菌丝生长最快,菌落直径达75.38 mm,极显著高于其他培养基的菌落直径。其次为PSA 培养基,其菌落直径极显著高于除PDA 培养基外的其他培养基。WA 培养基菌丝生长最慢,其菌落直径均极显著小于其他培养基。说明PDA 培养基是最适宜暹罗炭疽菌菌丝生长的培养基(表5)。

表5 病原菌菌丝生长、产孢对不同培养基的适应性
暹罗炭疽菌在PDA、PSA 培养基上产孢量最多,分别为46.25×104个/mL、44.02×104个/mL,两者间没有显著差异,但均显著高于其他培养基的产孢量。CMA、OA、百香果汁液、WA 培养基产孢量较少,均低于8.00×104个/mL,处理间没有显著差异。说明PDA、PSA 培养基是最适宜暹罗炭疽产孢的培养基(表5)。
2.2 室内杀菌剂毒力测定结果
以同种杀菌剂不同浓度的对数值为横坐标,同种杀菌剂不同浓度平均相对抑制率(%)的机率值为纵坐标建立毒力回归直线,算出毒力回归方程、相关系数(r)和半效应浓度(EC50),计算结果详见表6。
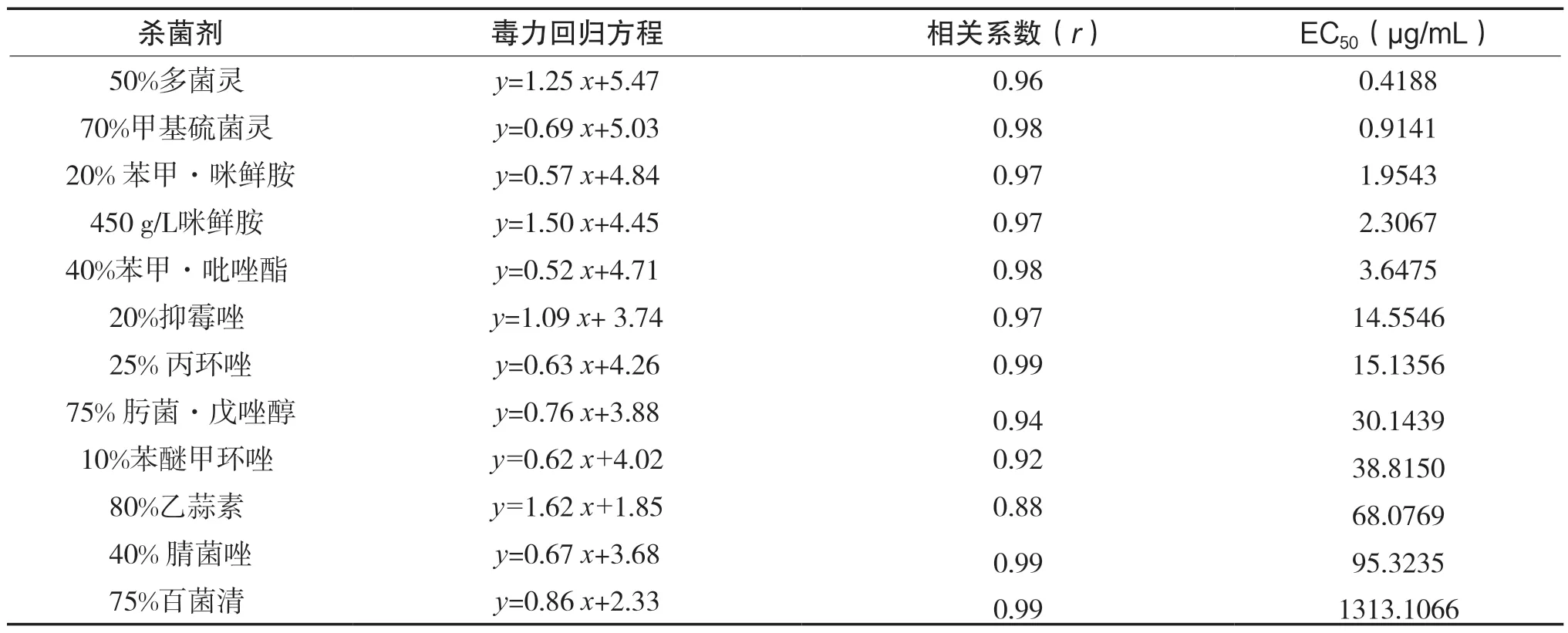
表6 12 种杀菌剂对紫果西番莲顶枯病病原菌的室内毒力测定结果
从表6 可知,12 种供试药剂的质量浓度与相对抑菌率的相关系数都在 0.88 以上,说明两者间呈正相关。EC50值可用来比较杀菌剂对供试菌株的毒力,EC50值越小表明药剂的毒力越强。12 种供试杀菌剂对顶枯病病原菌生物活性有差异,其中50%多菌灵毒力最强,EC50为0.4188 μg/mL;其次为70%甲基硫菌灵、20%苯甲·咪鲜胺、450 g/L 咪鲜胺、40%苯甲·吡唑酯,其EC50分别为0.9141 μg/ mL、1.9543 μg/mL、2.3067 μg/mL、3.6475 μg/mL;说明这几种杀菌剂均为紫果西番莲顶枯病田间防治参考用药。毒力中等的杀菌剂有20%抑霉唑、25%丙环唑、75%肟菌·戊唑醇、10%苯醚甲环唑,其EC50分别14.5546 μg/mL、15.1356 μg/mL、30.1439 μg/mL、38.8150 μg/mL。毒力较差的杀菌剂为80%乙蒜素、40%腈菌唑,其EC50为68.0769、95.3235 μg/mL;75%百菌清的毒力最差,其EC50为1313.1066 μg/ mL。
3 讨论与结论
本研究结果表明,不同培养条件对菌丝生长和产孢量、萌发率均有显著影响,且菌落颜色、厚度以及菌丝的生长状况明显不同。病原菌在pH 值为3.0~12.0 均能生长,最适生长pH 值为5.0~9.0。在pH 值为3.0~12.0 均能产孢,最适产孢量pH 值为7.0~8.0。菌丝在8~36℃均能生长,最佳生长温度为28℃。在12~36℃均能产孢,最佳产孢温度为28℃。孢子在pH 值为2.0~12.0 均能萌发,最佳萌发pH 值为5.0~10.0。孢子在12~40℃均能萌发,最佳萌发温度为20~36℃。其中,适宜菌丝生长的pH和温度与罗敦文[21]研究菠萝蜜蒂腐病病菌和刘在哲等[20]研究红肉苹果炭疽病病原暹罗炭疽菌适宜的pH 和温度基本一致。本研究中,在5 种光照条件下该病原菌均能生长并产孢,其中完全黑暗对菌丝生长和产孢最有利,与刘在哲等[16]研究认为红肉苹果炭疽病病原暹罗炭疽菌菌丝生长适宜光照条件为持续光照或光暗交替、王芳等[17]研究认为青星美人黑腐病病原菌最适菌丝生长光暗交替的观点有差异,而完全黑暗对菌丝生长和产孢最有利的研究结果与王芳等[17]的研究结果一致。该病原菌菌丝生长最适碳源为葡萄糖,产孢量最多碳源依次为乳糖、D-木糖和山梨醇。菌丝生长最适氮源为酵母粉和牛肉浸膏,产孢量最多氮源为牛肉浸膏,与罗敦文[21]对菠萝蜜蒂腐病菌菌丝生长的研究结果基本一致,但与王芳等[17]筛选获得青星美人黑腐病病原菌菌丝生长和产孢的最适碳源不同。该病原菌菌丝生长最适培养基为PDA,最适产孢培养基为PDA 和PSA,与罗敦文[21]对菠萝蜜蒂腐病菌菌丝生长的研究结果一致,与刘在哲等[16]对红肉苹果炭疽病病原暹罗炭疽菌菌丝的研究结果存在差异。本研究中紫果西番莲顶枯病病原菌在光照条件、碳氮源、培养基方面与前人研究结果存在的差异,可能与病原菌寄主、供试材料及试验环境等不同有关。
杀菌剂的毒力作用包括杀菌作用 (fungicidal action) 和抑/静菌作用 (fungistatic action),杀菌剂抑制病原菌生长发育的毒力测定可以在室内离体下进行,常用于抑制病原菌生长发育的化合物活性筛选,杀菌剂离体毒力测定方法,即用药剂直接处理病原菌或病原菌在含药基质上培养,测定药剂对病原菌生长发育的影响,在室内的离体毒力测定,能够在较小的空间内测定和比较多种药剂的毒力,测定条件容易控制,结果能够反映化合物本身的绝对毒力大小, 重复性好和可比性强,所以常常被指定为大多数杀菌剂的毒力测定方法[29]。在暹罗炭疽菌杀菌剂防治研究中,宋慧云等[3]对多菌灵、嘧菌酯、溴菌腈、苯醚甲环唑、吡唑醚菌酯、氨基寡糖素、嘧霉胺、福美双等8 种农药防效进行评价,认为多菌灵、吡唑醚菌酯和福美双可作为防治多穗柯炭疽病的化学杀菌剂。徐丹丹等[20]对苯醚甲环唑、咪鲜胺、吡唑醚菌酯、甲基硫菌灵等4 种农药防效进行评价,认为咪鲜胺、甲基硫菌灵可作为防治咖啡炭疽病首选杀菌剂。王志华等[22]对多菌灵、苯醚甲环唑、三唑酮、醚菌酯等4 种农药防效进行评价,认为多菌灵可湿性粉剂对金森女贞炭疽病抑制作用最强。本研究发现,苯并咪唑衍生物类杀菌剂(50%多菌灵)对紫果西番莲顶枯病病原菌毒力强度最强;咪唑类杀菌剂(70%甲基硫菌灵、450 g/L 咪鲜胺、20%抑霉唑)对病原菌毒力强度较强,与前人研究的结果相似;三唑类杀菌剂(25%丙环唑、10%苯醚甲环唑)对病原菌毒力强度中等;有机硫类杀菌剂(80%乙蒜素)、吡咯类杀菌剂(40%腈菌唑)对病原菌毒力强度较差;取代苯类杀菌剂(75%百菌清)对病原菌的毒力强度最差。本研究还发现,复配杀菌剂的毒力强度强于单剂杀菌剂,其20%苯甲·咪鲜胺微乳剂(含苯醚甲环唑5%和咪鲜胺唑15%)的EC50为1.9543 μg/mL,均低于单剂的10%苯醚甲环唑或450 g/L 咪鲜胺的EC5(02.3067、38.8150 μg/mL);40%苯甲·吡唑酯(苯醚甲环唑15%,吡唑醚菌酯25%)的EC50为3.6475 μg/mL 明显低于单剂的10%苯醚甲环唑(38.8150 μg/mL)。
本研究对紫果西番莲顶枯病病原菌的生物学特性进行分析,并用12 种杀菌剂对其进行室内毒力测定,结果可为紫果西番莲顶枯病防治提供技术参考。由于杀菌剂毒力测定试验是在室内进行,田间防治效果还与病原菌孢子[11]、寄主植物生长环境[13]、杀菌剂特性[30]、施药时期、气候、药械及其他不可预见因素等有关[31]。因此,田间防治效果还需进一步验证。
