光调控香草兰花芽分化的生理机制初探
2023-06-01张何信浦同华庄辉发邢诒彰
王 怡,张何信,浦同华,庄辉发,3,邢诒彰,3,王 辉*
[1.中国热带农业科学院香料饮料研究所,海南万宁 571533;2.海南省林业科学研究院(海南省红树林研究院),海南海口 571100;3.海南省热带香辛饮料作物遗传改良与品质调控重点实验室,海南万宁571533]
香草兰(Vanilla planifoliaAndrews)又名香子兰、香荚兰,原产于热带雨林,是兰科香荚兰属的多年生热带攀援藤本香料植物,因其用途广泛,在医药、保健等行业具有极高的关注度[1]。目前,香草兰花芽分化率低是生产中亟待解决的技术问题。遮阴会降低光合速率并改变光谱质量,影响植物的光合作用和光形态建成[2]。碳水化合物是一种结构物质,同时也是能量供给源,其积累与花芽的分化密切相关,对花芽分化起着重要作用[3]。基于此,部分学者开展了遮阴影响香草兰花芽分化及碳水化合物变化的研究[4-5]。笔者在探讨光合诱导特性的基础上,深入开展遮阴对碳水化合物合成和相关酶活性的影响研究,旨在进一步探明光调控香草兰花芽分化的生理机制,为提高香草兰花芽分化率,实现稳产高产奠定理论基础。
1 材料与方法
1.1 试验地概况
试验地位于中国热带农业科学院香料饮料研究所高龙试验基地(东经110°10′,北纬18°41′)。试验地土壤为砖红壤,有机质含量为18 g·kg-1,土壤肥力中等,年平均气温24.0 ℃,年降水量2400 mm。
1.2 试验材料
试验所用香草兰品种为热引3 号香草兰,荫蔽条件下露地栽培。
1.3 试验设计
香草兰全光照条件下无法生长,无实际生产意义,故未设0%(全光照)对照。遮阴处理设50%、75%、90%(皆为遮光度)3 个水平,3 种处理均由同一种遮阳网叠加而成,各处理统一进行肥水、杂草和病虫害等管理。于2020 年12 月22 日进行换网遮阴处理,每个处理3个重复。
1.4 试验方法
1.4.1 取样
在花芽分化阶段,作为生长中心的芽体与距离其最近的健康叶片间有较强的库源关系,是光合产物与矿质营养的分配中心[6]。因此,选择生长良好且相对稳定的香草兰植株,在花芽分化的各个阶段,选取最接近芽体的健康叶作为芽体功能叶,并在芽体功能叶处做标记,每个重复标记40株。
从2021 年2 月底开始,每隔15 d 选择晴天采集1次标记叶片,取样时期分别为花芽特征分化期、花芽分化初期(Ⅰ);花芽分化中期、花序分化初期(Ⅱ);花序分化中期、花分化初期(Ⅲ)3 个阶段。每次采集叶片剪去先端和基部后,将叶片中部用铝箔包好在液氮中固定20 min,用冰盒带回实验室。放入-80 ℃超低温冰箱中保存待用,实验前粉碎后混匀。
1.4.2 测定
1)光诱导特性测定。在大约25 ℃的大气空气条件,使用Dual-PAM-100 测定系统(Heinz Walz,Effeltrich,德国)活体测量。2)SS、SPS 活性测定。称取0.1 g 鲜叶于2 mL 离心管中,用高通量组织研磨机粉碎,加入1 mL提取液匀浆,8000 r·min-1、4 ℃离心10 min,取上清液采用试剂盒测定。3)在开花中后期,统计标记茎段的叶芽数和花芽数,计算各重复平均花芽分化率及总芽数。
1.5 数据处理与统计方法
以可溶性糖与淀粉含量之和作为非结构性碳水化合物总量。利用SPSS 20.0软件进行方差分析和显著性检验,利用Origin 9.1软件整理数据及制作图表。
2 结果与分析
2.1 对光诱导特性的影响
如图1 所示,在光照强度增加后,光系统II 电子传递速率(ETRII)随着时间的推移逐渐增加,在240~360 s 内迅速增加到最大值的95%,其中90%遮光度处理条件下所需时间最短且ETRII 最高,但各处理差异不显著(p>0.05,下同)。
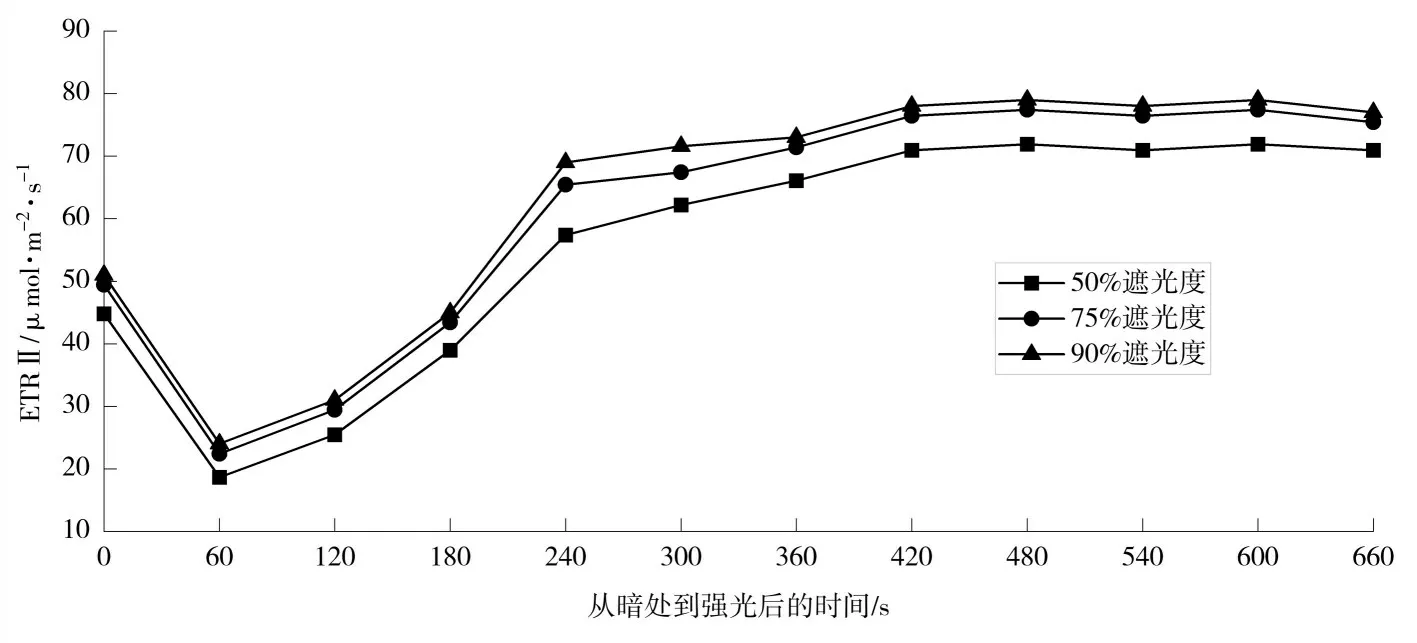
图1 不同遮阴处理下香草兰叶片光反应中心电子传递速率的变化
2.2 对可溶性糖含量的影响
香草兰花芽分化不同时期叶片中可溶性糖的含量差异显著(p<0.05,下同),可溶性糖含量在前期大量积累,中期显著降低,后期又逐渐积累。前期(Ⅰ),50%和90%遮光度处理显著增加了功能叶中可溶性糖含量;中期(Ⅱ),90%遮光度处理可溶性糖含量达3.15 mg·g-1(FW),显著高于75%遮光度处理;后期(Ⅲ),90%遮光度处理可溶性糖含量最高,且显著高于50%和75%遮光度处理(见图2)。总体上,在香草兰花芽分化期,90%遮光度处理可溶性糖含量最高,达4.00 mg·g-1(FW),50%遮光度处理次之,75%遮光度处理最低(见表1)。
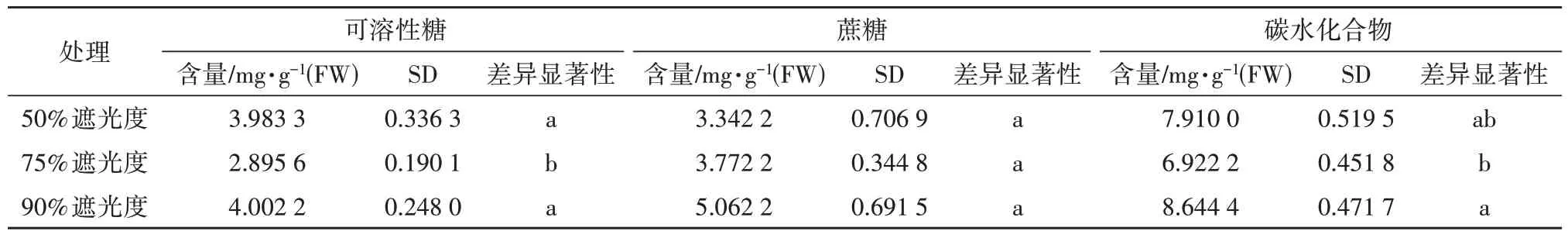
表1 不同遮阴处理下香草兰叶片糖类含量

图2 不同遮阴处理下香草兰叶片可溶性糖含量动态
2.3 对蔗糖含量的影响
如图3 所示,香草兰花芽分化前期的蔗糖含量显著高于中、后期,随着花芽孕育,蔗糖明显被消耗后又显著上升,大量积累。前期(Ⅰ),90%遮阴处理下香草兰功能叶中蔗糖含量达7.04 mg·g-1(FW),显著高于75%遮阴处理;中期(Ⅱ),75%遮阴处理蔗糖含量略高于90%遮阴处理,显著高于50%遮阴处理。如表1所示,90%遮阴处理下蔗糖总体含量最高。
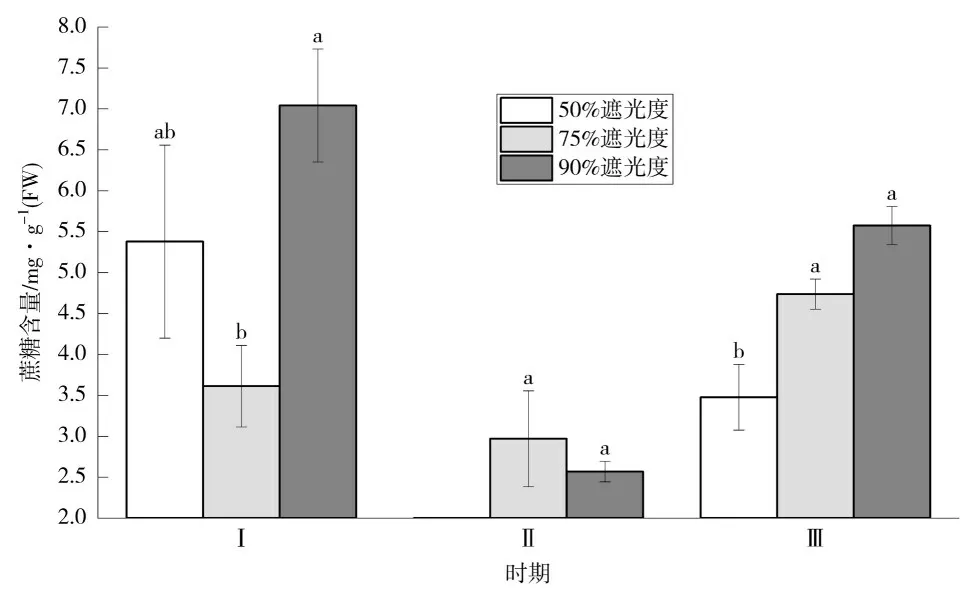
图3 不同遮阴处理下香草兰叶片蔗糖含量动态
2.4 对非结构性碳水化合物总量的影响
如图4所示,前期(Ⅰ),各处理条件下叶片非结构性碳水化合物大量积累,50%遮阴处理最高,达9.72 mg·g-1(FW),比75%遮阴处理提高了20.15%;中期(Ⅱ),50%遮阴处理非结构性碳水化合物总量最低;后期(Ⅲ),非结构性碳水化合物总量在90%遮阴处理下最高,显著高于50%和75%遮阴处理。总体上,香草兰花芽分化期间芽体功能叶中非结构性碳水化合物含量依次为90%>50%>75%,90%遮阴处理显著高于75%遮阴处理(见表1)。
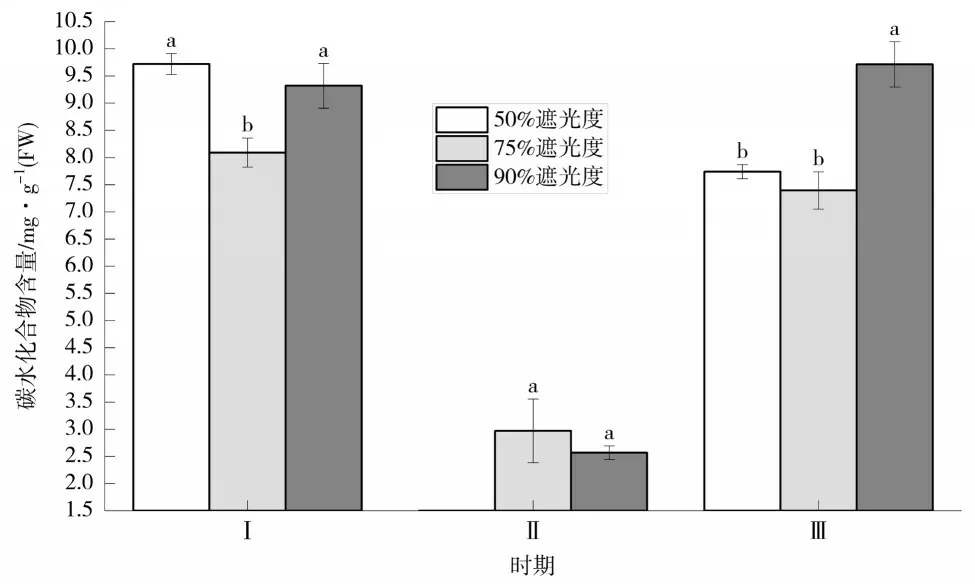
图4 不同遮阴处理下香草兰叶片碳水化合物总量动态
2.5 对蔗糖合成酶(SS)活性及蔗糖磷酸合成酶(SPS)活性的影响
如表2 所示,各处理SS 活性动态变化趋于一致,在花芽分化后期急剧提高,前期以50%遮光度处理下SS 活性最高;中期以90% 遮光度处理最高,为817.36µg·g-1·min-1(FW);分化后期,50%遮光度处理最高,比75%和90%遮光度处理分别高18.41%和11.17%。如表3 所示,75%和90%遮光度处理下,芽体功能叶中SPS 活性动态一致,在前期达最大值,随花芽生长发育又逐渐降低;而50%遮光度处理下,在分化中期达到最大值,且高于其他处理,而后急剧降低。
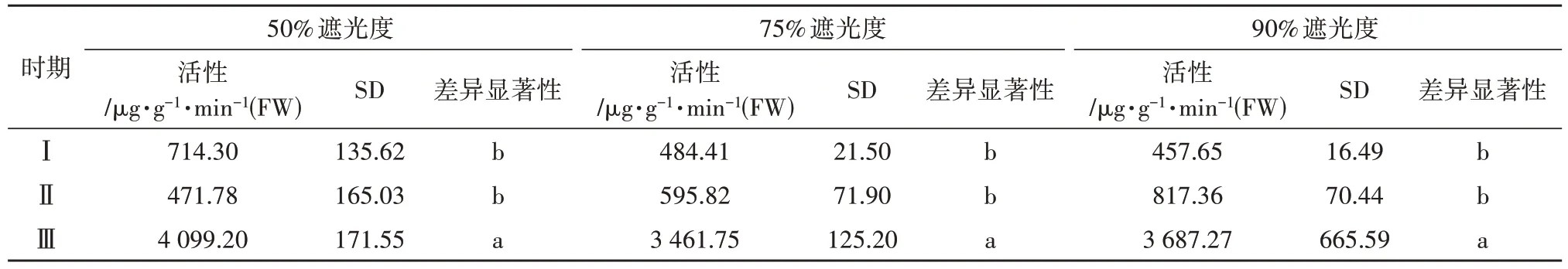
表2 不同时期和遮阴处理下香草兰叶片蔗糖合成酶活性

表3 不同时期和遮阴处理下香草兰叶片蔗糖磷酸合成酶活性
2.6 对花芽分化的影响
如表4 所示,50%遮光度处理下香草兰总芽数显著高于75%和90%遮光度处理;50%遮光度处理下花芽分化率达62.20%,比75%和90%遮光度处理分别高出24.58和60.53个百分点。
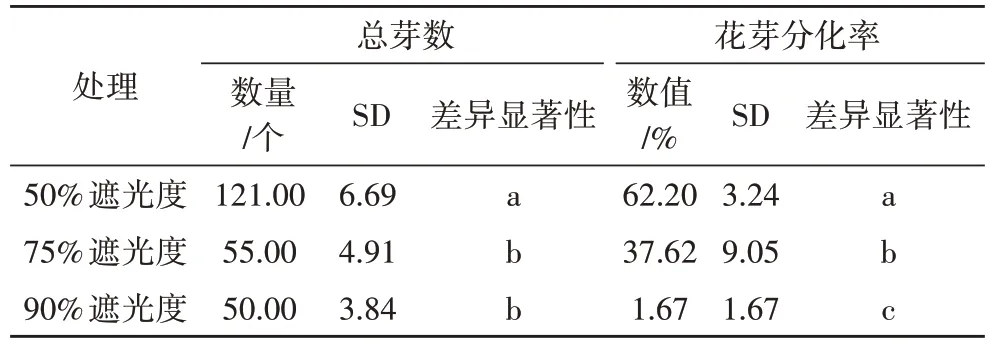
表4 不同遮阴处理下香草兰叶片糖类含量
3 结论与讨论
在花芽萌发和花器官形成中,高含量的可溶性糖可促进花芽分化。同样,香草兰花芽分化期间,需要大量可溶性糖和淀粉维持花芽生长发育,花芽发育的速度也由可溶性糖和淀粉含量的高低决定。光照对植物体内碳水化合物积累有所影响,遮阴有利于蔗糖积累,从而为植物体生长发育提供充足物质基础,促进其成花。本研究表明,遮阴处理影响了香草兰功能叶中的碳水化合物积累和相关代谢酶活性,其均与花芽分化表现出一定的相关性。在50%和90%遮光度处理下,香草兰叶片非结构性碳水化合物总量均高于75%,可更好地为花芽分化提供能量,促进成花;但75%遮光度下花芽分化率显著高于90%,具体原因需进一步探讨。
蔗糖的合成与输出是植物成熟叶片的主要功能,SS 和SPS 协同调控植物体内碳水化合物的积累和分配。目前,大部分研究显示SPS 活性与蔗糖含量存在正相关关系,而与淀粉含量呈负相关;SS活性对蔗糖积累有正效应,与淀粉含量负相关[7]。本实验中,SS与蔗糖含量正向相关,影响了植株的蔗糖积累,与相对淀粉含量正相关,调节碳同化产物在蔗糖和淀粉之间的分配。
综上,香草兰作为景天酸代谢(CAM)植物,在光照强度增加后,ETRII 在240~360 s 内迅速增加到最大值的95%,这说明其表现出快速的光合诱导,90%遮光度处理条件下苹果酸相对更充足,会快速脱羧苹果酸;50%和90%遮光度下能提高香草兰功能叶中的碳水化合物,但90%遮光度在一定程度上抑制了香草兰花芽分化,从而降低其花芽分化率。碳水化合物的积累虽为花芽分化所需,适宜的碳水化合物对植物花芽分化有重要作用,但高含量的碳水化合物则不一定导致成花,这与孙敏研究结果一致[8]。在75%遮光度下香草兰总芽数及花芽分化率相对较高,有利于香草兰产量提高和植株发育。
