小麦抗倒伏茎秆形态指标及QTL分析
2023-05-15赵晓雪司红起马传喜
汪 涛,张 毅,赵晓雪,陈 璨,司红起,马传喜,卢 杰
(安徽农业大学,安徽 合肥 230036)
随着小麦(TriticumaestivumL.)单产的提高及水肥条件的改善,倒伏逐渐成为限制小麦高产稳产的主要因素之一。我国是世界上重要的小麦生产和消费大国,每年因倒伏而损失的粮食产量超过200万t[1],倒伏易导致收获延迟、品质及籽粒商品性变差等不利后果[2]。根据发生部位不同,小麦倒伏分为根倒伏和茎倒伏2种类型,茎倒伏是小麦生产中常见类型,由茎秆节间的弯曲或者断裂造成,主要发生在基部第2节间,且乳熟期是极易发生倒伏的时期[3]。育种实践表明,小麦品种的抗倒性与株高和茎秆强度显著相关[4],但株高的过度矮化会造成植株地上部分生物量降低、群体郁闭、病害加重,进而影响产量提升[5],当前,小麦抗倒伏育种更注重茎秆质量的改善,由矮秆抗倒向壮秆抗倒转变[6-8]。因此,研究小麦茎秆抗倒伏遗传机理,将有助于培育抗倒伏小麦品种。
研究表明,除株高外,小麦茎秆形态及结构特征与抗倒伏性相关。小麦抗倒伏性与茎秆基部节间长度呈负相关,与基部第2节间的茎秆壁厚呈显著正相关,倒伏易发生在基部第2节间,具有较短的基部第2节间、较大的茎秆粗度及壁厚的品种,其抗倒伏能力较强,反之则弱[4,9-12]。小麦茎秆中木质素和纤维素含量与抗倒伏能力相关[13],研究发现,小麦茎秆纤维素含量越高,茎秆抗压强度越高,纤维素含量低的品种茎秆更脆,茎秆强度也弱,更容易发生倒伏[2,14-15]。此外,小麦抗倒伏遗传机理的研究亦有报道,潘婷等[16]在染色体1B、3B、4A、4B和5A上定位到23个影响茎秆强度、茎秆直径和髓腔直径的QTLs,表型变异解释率在3.5%~44.0%。刘凯等[17]利用90K芯片在4B染色体上检测到9个控制小麦茎秆强度和基部第2节间壁厚的QTLs,可以解释9.40%~36.60%的表型变异。此外,通过全基因组关联分析,在自然群体的染色体1A、1B、2B、2D、3A、3B、4A、4B、5A、5B、5D、6B、7A、7B和7D上发现了37个与茎秆强度相关的标记,解释了7.76%~13.77%的表型变异。潘婷[18]利用重组自交系 (Recombinant inbred lines,RILs),在3B和5A染色体上识别到2个茎秆强度相关的QTLs。Ma等[19-22]克隆了Ta-CCR1、Ta-COMT、Ta-CAD1等3个小麦茎秆木质素合成基因。综上,小麦抗倒伏是受多因素、多基因控制的复杂性状,因此,确定适宜的抗倒伏评价指标,深入挖掘抗倒伏相关的基因或QTL,开发实用性分子标记或优异等位变异,将有助于小麦抗倒伏性的遗传改良。
本研究以茎秆强度差异显著的小麦品种安农0711和河农825构建的RILs群体为基础,测定多个环境下该群体乳熟期茎秆强度值及基部第2节间外茎、壁厚、长度、木质素、纤维素等形态结构指标,同时利用小麦55K芯片多态性SNP标记对上述指标进行基因型鉴定,旨在探究影响小麦茎秆强度的形态结构指标,鉴定茎秆强度相关QTL,为小麦抗倒伏遗传机理的研究提供参考。
1 材料和方法
1.1 试验材料与试验设计
以茎秆强度值差异显著且株高相近的小麦品种安农0711和河农825构建的F9~F10120份重组自交系(RILs)群体为研究材料。
RILs群体于2019—2020年(以下称2020年)种植于合肥大杨店高新技术农业示范园(20HF:31.9 0N,117.2 0E)和淮北市濉溪县五铺农场(20HB:33.7 0N,116.7 0E),2020—2021年(以下称2021年)种植于淮北市濉溪县五铺农场(21HB)和济源市安徽隆平高科试验站(21JY:35.04N,112.65E),每份试验材料播种2行,行长2 m,行间距0.25 m,株距0.15 m,2次重复,大田常规管理。
1.2 测定项目及方法
参照肖世和等[23]描述的茎秆强度测定方法,稍做改动。花后28 d(乳熟期)测定茎秆强度,具体为每个家系选择相邻的、长势相似的5个单茎,使用YYD-1A型茎秆强度仪(浙江拓普仪器有限公司)测定供试材料茎秆强度(SS),5次重复。花后28 d随机选择5株代表株测量表型数据,用游标卡尺测量基部第2节间外径(SBD)和基部第2节间壁厚(SWT),使用20 cm直尺测量基部第2节间长度(SBL)。采用硫酸蒽酮比色法[24]测定小麦基部第2节间纤维素含量(SCC),紫外分光光度计法[25]测定基部第2节间木质素含量(SLC)。
1.3 多元线性回归分析
以茎秆强度为因变量(y),茎秆性状指标第2节间长(x1)、第2 节间粗(x2)、第2节间壁厚(x3)、第2节间纤维素含量(x4)和第2节间木质素含量(x5)为自变量,分别应用SPSS软件Linear过程“逐步法”建立多元线性回归方程。
1.4 小麦55K芯片扫描及QTL分析
利用小麦55K芯片(北京博奥晶典生物科技有限公司)对(安农0711×河农825)衍生的120个RILs家系和双亲进行扫描。利用Excel将55K芯片中在双亲安农0711和河农825间存在基因型差异的SNP进行筛选和提取,剔除双亲都缺失、单亲杂合或缺失的SNP标记,用于茎秆强度QTL分析。
QTLIciMapping v4.1软件[26](http://www.isbreeding.net/software)中混合线性模型复合区间作图法,基于2017—2021年的茎秆强度表型值进行QTL分析,将LOD阈值设为大于2.0。
1.5 数据处理
表型性状数据整理由Excel软件完成,表型数据间的相关性分析由IBM SPSS Statistics 25(www.spss.com)软件完成,Origin 2022用于图表制作。
2 结果与分析
2.1 RILs群体表型性状分析
从表1可以看出,2020年合肥、2020年淮北、2021年淮北和2021年济源4个试验环境下,安农0711的茎秆强度、基部第2节间茎粗、壁厚均高于河农825,第2节间长度低于河农825; 2020年合肥和2020年淮北2个环境下第2节间纤维素含量和木质素含量均高于河农825。RILs群体的茎秆性状在各环境下均呈连续性分布(图1),茎秆强度、基部第2节间长度、茎粗和木质素含量在各环境中的偏度和偏峰绝对值均小于1,符合正态分布,基部第2节间壁厚和纤维素含量也基本符合正态分布,表明这些茎秆性状均是多基因控制的数量性状。表型数据变异系数8%~38%,在6个性状均存在超亲分离现象,且各环境下均表现一致。在这些性状中,茎秆强度和第2节间壁厚变异系数分别为26%~38%和10%~23%,说明这2个性状具有较高的遗传变异和改良潜力。
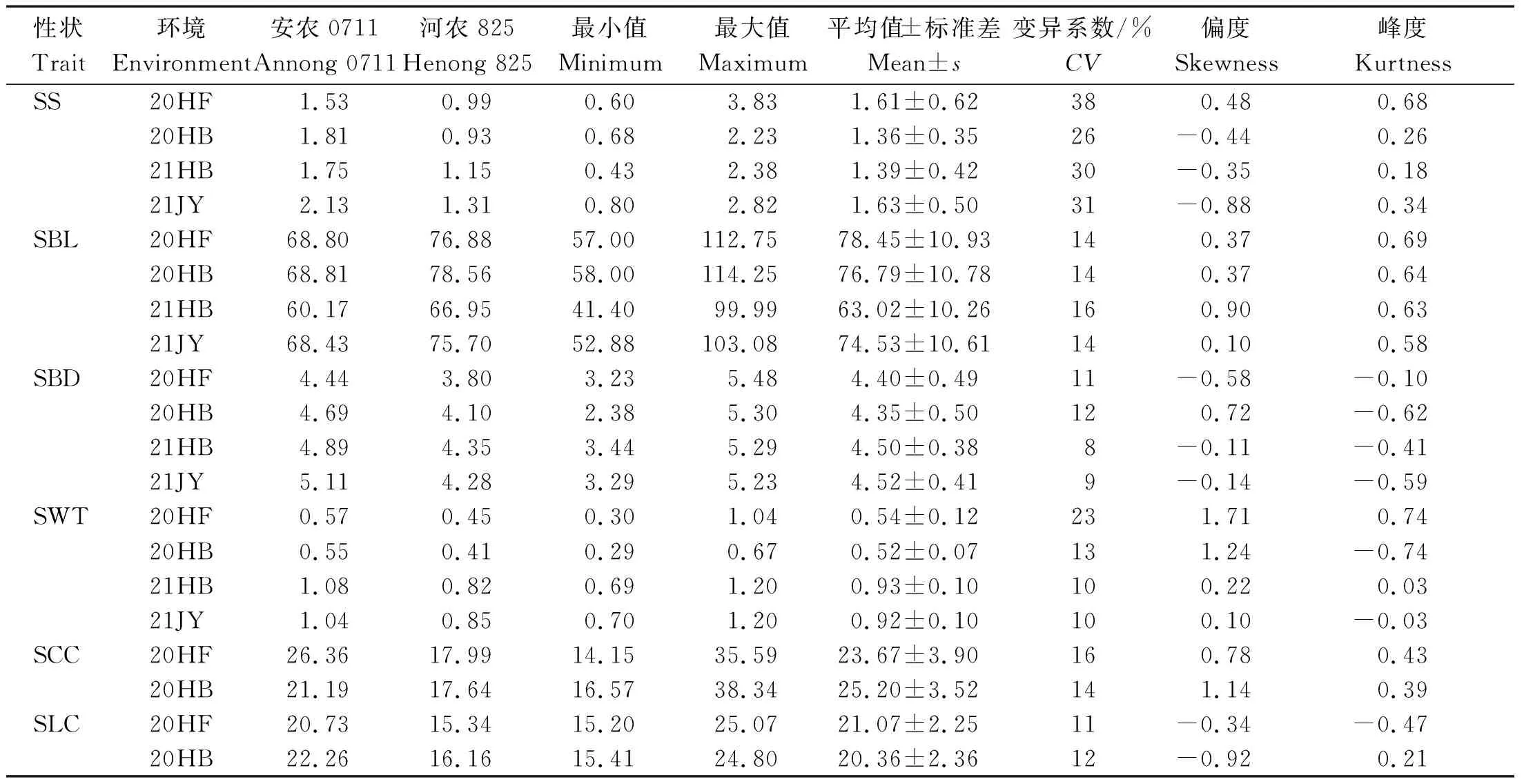
表1 安农0711/河农825 RILs群体茎秆性状统计Tab.1 Statistics of stem traits of Annong 0711/Henong 825 RILs population
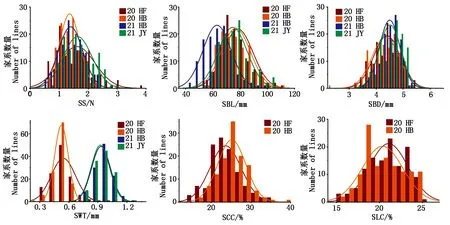
图1 RIL群体茎秆性状和茎秆强度的表型分布Fig.1 Phenotypic distribution of stem traits and stem strength in RIL populations
2.2 茎秆强度与其形态特征相关性分析
表2表明,RILs群体茎秆强度与基部第2节间长度呈负相关,在2021年淮北和2021年济源环境下达极显著水平(P<0.01);与基部第2节间茎粗和壁厚呈显著正相关(P<0.05),且2021年济源均达到极显著(P<0.01);在2020年合肥和2020年淮北2个环境下与基部第2节间纤维素含量和木质素含量均达到极显著正相关(P<0.01),说明第2节间长度越短,茎粗和壁厚越大,纤维素和木质素含量越高,越有利于增强茎秆强度,提高植株的抗倒伏性。
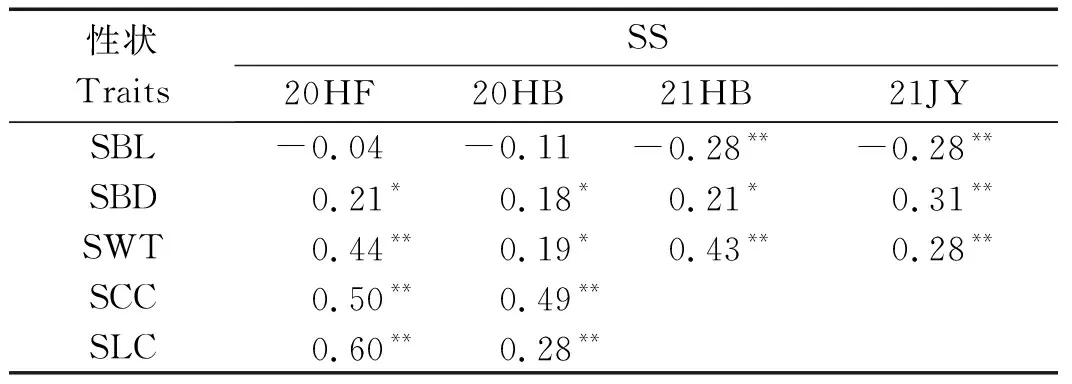
表2 RIL群体茎秆强度与茎秆性状相关性分析Tab.2 Correlation analysis of stem strength and stem traits in RIL population
2.3 茎秆性状对茎秆强度的回归分析
由表3可知,对2020年合肥环境下茎秆性状与茎秆强度进行多元回归分析,VIF值均小于5,说明自变量之间不存在多重共线性,模型构建良好。在95%显著水平下,从对小麦茎秆强度有影响的茎秆性状中逐个淘汰不显著的自变量,筛选出回归系数达到显著水平的茎秆性状入选回归方程。得到最优线性回归方程:
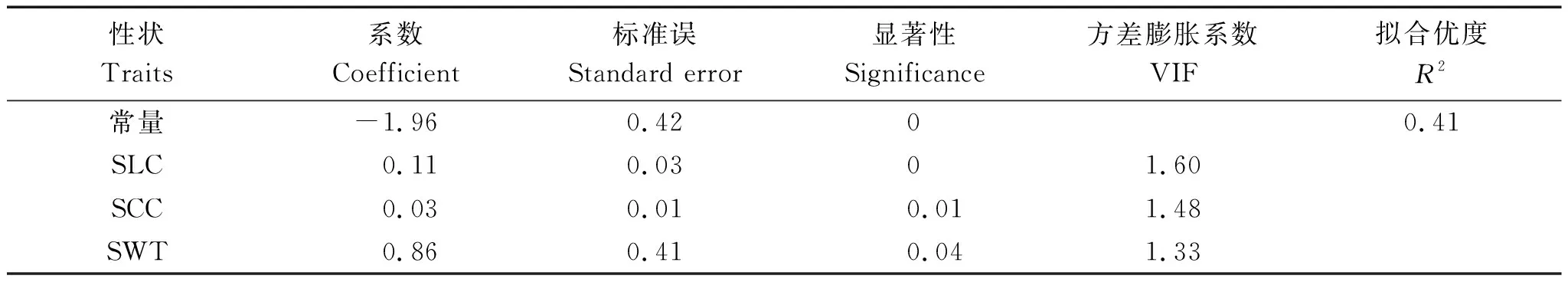
表3 茎秆性状对茎秆强度的多元回归分析结果(2020年合肥)Tab.3 Multiple regression analysis results of stem traits on stem strength(Hefei,2020)
y=-1.96+0.11x5+0.03x4+0.86x3
该方程中,基部第2节间壁厚(x3)回归系数最大,为0.86,表明基部第2节间壁厚对小麦茎秆强度的贡献最大;基部第2节间纤维素含量(x4)回归系数为0.03,表明基部第2节间纤维素含量越高,小麦茎秆强度越大;基部第2节间木质素含量(x5)回归系数为0.11,表明基部第2节间木质素含量对茎秆强度有较大影响。3个茎秆性状共解释41%的茎秆强度变化。
由表4可知,对2020年淮北环境下茎秆性状与茎秆强度多元进行回归分析,VIF值均小于5,说明自变量之间不存在多重共线性,模型构建良好。在95%显著水平下,从对小麦茎秆强度有影响的茎秆性状中逐个淘汰不显著的自变量,筛选出回归系数达到显著水平的茎秆性状入选回归方程。得到最优线性回归方程:y=0.13+0.05x4。

表4 茎秆性状对茎秆强度的多元回归分析结果(2020年淮北)Tab.4 Multiple regression analysis results of stem traits on stem strength(Huaibei,2020)
该方程中,基部第2节间纤维素含量(x4)回归系数为0.05,解释24%的茎秆强度变化,表明基部第2节间纤维素含量越高,小麦茎秆强度越大。
2.4 小麦茎秆性状QTL鉴定
如表5所示,基于小麦55K芯片筛选得到的11 135个二态性SNP标记和4个环境下茎秆性状表型值,共检测到19个与茎秆强度和茎秆性状相关的QTLs,分布在1A、1D、2B、2D、4D、5A、5B、5D和7B染色体上,能够解释7.67%~65.33%的表型贡献率。其中,在1D、2B、2D、4D、5A、5B、5D和7B染色体上共检测到8个控制小麦茎秆强度的QTLs,解释表型贡献率为7.67%~61.46%,增效基因均来自安农0711,Qss.ahau-5B、Qss.ahau-5D和Qss.ahau-7B均能在2个环境中检测到,分别解释8.57%~9.99%,10.35%~13.29%和7.67%~7.86%的表型变异率。在2021年济源环境检测到的Qss.ahau-5A解释61.46%的表型变异率,可增加小麦茎秆强度0.39N。检测到2个控制小麦基部第2节间长度的QTLs,位于1D和5D染色体,解释表型贡献率7.96%~15.48%,增效基因均来自河农825。Qsbl.ahau-5D可在2020年合肥和2021年淮北2个环境检测到,解释12.10%~15.48%的表型变异率,可降低第2节间长度3.58~4.32 mm。在1A和1D染色体上检测到2个控制基部第2节间茎粗的QTLs,增效基因均来自安农0711,可增加第2节间茎粗0.15~0.23 mm。Qsbd.ahau-1D与调控茎秆强度的Qss.ahau-1D位置相同,解释9.42%的表型变异率。检测到1个控制基部第2节间壁厚的QTLs,位于1D染色体上,增效基因来源于安农0711,该位点与调控第2节间长度的Qsbl.ahau-1D区间相同,解释10.76%的表型变异率。在1D和4D染色体上检测到2个控制基部第2节间纤维素含量的QTLs,解释9.51%~10.76%的表型变异率,增效基因均来自安农0711。检测到4个控制基部第2节间木质素含量的QTLs,分布在1A、5A、5D和7B染色体上,解释10.82%~65.33%的表型变异率,加性效应均来自安农0711,Qslc.ahau-5D在2020年合肥和2020年淮北2个环境均被检测到,解释10.82%~14.23%的表型变异率。Qslc.ahau-5A和Qss.ahau-5A位置相同,表型变异率分别为65.33%和61.46%。

表5 RILs群体茎秆强度相关的QTLTab.5 QTLs related to stem strength in RILs population
3 结论与讨论
在小麦抗倒伏研究中,茎秆强度、倒伏指数、茎秆弹性、茎秆折断力及自然倒伏的级别与面积等指标均可用于评价其抗倒伏性,其中,茎秆强度指标因与小麦的抗倒伏性呈正相关且检测容易、重复性好,应用较为广泛[27]。研究表明,小麦茎秆强度与节间长度、茎粗、壁厚等多种茎秆性状相关[28]。前人报道小麦基部节间茎粗和壁厚与茎秆强度呈极显著正相关[6,29],本研究发现,在4个环境下,茎秆强度与基部第2节间茎粗和壁厚呈显著或极显著正相关,与基部第2节间长度呈负相关。表明提高小麦基部第2节间茎粗和壁厚,有助于提高茎秆强度,对增强小麦抗倒伏能力具有重要意义。此外,本研究结果显示,2020年合肥和淮北2个环境下茎秆强度与基部第2节间纤维素含量和木质素含量均达到了极显著水平,这与陈晓光[30]的研究结果较为一致,表明基部第2节间纤维素含量和木质素含量是影响小麦茎秆强度的重要指标。多元回归分析方程表明,基部第2节间纤维素含量在2020年合肥和淮北环境下均是重要的因变量,综上可见,基部第2节间纤维素含量是影响小麦茎秆强度的关键指标。
本研究基于11 135个多态性SNPs标记和4个环境下的茎秆强度、第2节间长度、茎粗和壁厚以及2个环境下的第2节间纤维素和木质素含量表型值,共检测到19个与茎秆性状相关的QTLs位点,分布在9条染色体上,这说明小麦茎秆性状是多基因控制的数量性状,受环境影响较大。本研究在2D染色体上定位的Qss.ahau-2D位于标记AX-94774565和AX-108995193之间,解释18.59%的表型贡献率,郭会君[31]在2D上的附近区域也定位到1个控制茎秆强度的QTL,解释12.00%的表型贡献率,说明该区域可能存在控制茎秆强度的主效QTL。郭会君[31]在3B和5B上定位到调控基部第2节间茎粗的QTL,潘婷[18]在1B和3B上定位到控制茎粗的QTL,本研究检测到2个调控基部第2节间茎粗的QTL位点位于1A和1D染色体上,不同于前人研究结果。目前,调控茎秆基部第2节间壁厚的QTL分布在2A、2D、3B、4A、4B、4A、5D和6A染色体上[18,32],而本研究检测到调控基部第2节间壁厚的QTL位于1D染色体上,不同于前人的研究结果,可能是一个新的位点。本研究中,在1D染色体上,与标记AX-110771095和AX-109431570连锁的QTL位点同时控制基部第2节间长度、壁厚及纤维素含量3个性状,解释表型贡献率为7.96%~10.76%,可作为小麦抗倒伏研究的候选位点。
