调控高粱分蘖高度的基因及表达分析
2023-05-15程庆军王绘艳平俊爱张福耀
王 瑞,程庆军,王绘艳,巨 岚,平俊爱,张福耀
(山西农业大学 高粱研究所,高粱遗传与种质创新山西省重点实验室,山西 晋中 030600)
高粱(Sorghumbicolor)是全球第五大类粮食作物,是世界上数亿人口的主食[1-2],其作为C4谷物和饲草作物,可用于粮食、饲料、糖和生物能源生产[3]。高粱抗旱且耐盐碱和瘠薄土壤,被视为干旱和盐碱土壤农业区农业可持续发展的一种重要作物[4-6]。高粱也是我国的重要旱粮作物之一,目前全国种植面积约80万hm2,主要分布在东北、华北、西南等区域。
株高是非常重要的农艺性状之一,在很大程度上影响植株的总生物量和产量。有关高粱株高的研究早在20世纪50年代就已开始,Quinby等[7]研究认为,高粱株高由Dw1、Dw2、Dw3和Dw4等4个株高的基因控制。随着分子生物学的发展,调控高粱株高基因的研究逐步深入,Lin等[8]利用高粱和拟高粱(S.bicolor×S.propinquum)构建的F2群体为研究对象,在1,3,6,7,9号染色体上鉴定出6个与株高相关的QTL;Pereira等[9]用CK60×PI229828的F2群体在高粱的6,7,9,10号染色体上分别检测到4个与株高变异相关的QTL;其他研究者也先后在高粱上找到了多个控制高粱株高的QTL[10-17]。Dw3是高粱中第一个被克隆的株高基因[18],它编码一个调节高粱茎秆中生长素极性运输的ABCB1类型的生长素输出载体蛋白且同源于玉米(Zeamays)的Br2基因[19-20]。之后,Lin等[8]、Pereira等[9]和Brown等[21]把Dwl基因定位于9号染色体上。在此基础上,Hilley等[22]和Yamaguchi等[23]2016年克隆了该基因Sobic.009G229800,它编码一个高度保守蛋白,该蛋白参与细胞增殖调节。2017年,Hilley等[24]利用图位克隆的方法成功分离到Dw2基因(Sobic.006G067700),该基因同源于拟南芥(Arabidopsisthaliana)AGC蛋白酶家族中的KIPK且编码蛋白激酶。
分蘖性是高粱的重要农艺性状之一,分蘖力强的高粱品种在补偿缺苗断垄造成的基本苗不足和增加产量上有明显的优势。然而,大多数高粱品种分蘖株高高于主茎,造成品种整齐度差,尤其是给机械化收获带来重大影响。研究表明[25],分蘖高度的基因调控与Dw1、Dw2、Dw3和Dw4等4个株高基因不同,前期用分蘖与主茎株高一致的高粱品系K35-Y5与分蘖明显高于主茎的高粱恢复系1383杂交,F1自交获得F2分离群体,采用BSA和SLAF技术将高粱分蘖与主茎株高一致基因定位到Chr.9上,在关联区域发现4个非同义突变的SNP,对应到Sobic.009G197901.1、Sobic.009G213300.1和Sobic.009G221200.1等3个基因上。
为了进一步解析相关基因在调控分蘖高度中发挥的作用,本研究选取分蘖与主茎株高一致和不一致的极端材料组成自然群体作为研究对象,对4个SNP位点进行验证,然后运用实时荧光定量PCR(Quantitative Real-time PCR,qRT-PCR)技术对相关基因组织特异性的表达进行分析,以期确定高粱分蘖株高基因的调控模式,为适宜机械化生产高粱品种选育提供一定的理论基础。
1 材料和方法
1.1 试验地概况
试验设在山西农业大学高粱研究所榆次试验基地。该地位于北纬37°5′,东经112°5′,海拔780 m,年平均气温9.8 ℃,年降雨量418~483 mm,年日照时数2 662 h。2021年高粱生长季即4—9月平均气温分别为11.9(4月),18.4(5月),22.8(6月),24.3(7月),22.7(8月),18.7 ℃(9月),与常年气温相近。田间顺序排列2行区种植,行长5 m,行距45 cm,留苗密度9.0万株/hm2,生育期间管理同大田。开花后调查分蘖与主茎株高的一致性:分蘖株高与主茎株高同等或株高差≤5 cm,记为株高一致;分蘖株高与主茎株高差> 5 cm记为不一致[26]。
1.2 试验材料
调控高粱分蘖高度基因研究选用32份高粱主要亲本系为试验材料,其中15份为分蘖与主茎同等高度,株高整齐一致;17份为分蘖高于主茎,株高不一致(表1)。
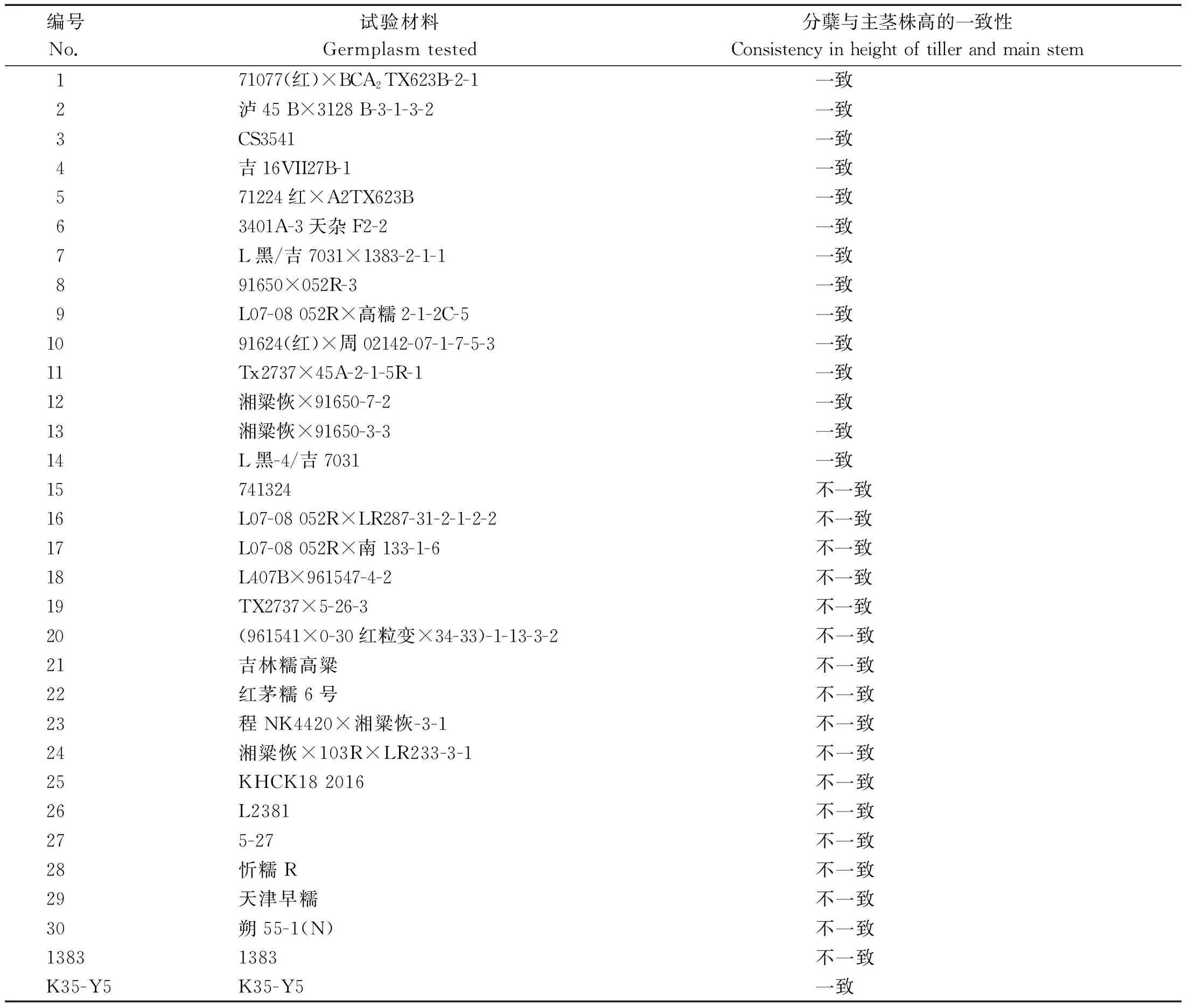
表1 试验材料及分蘖性状表型Tab.1 Germplasm tested and the tillering phenotypes
基因表达分析研究选用分蘖与主茎同等高度,株高整齐一致的品种K35-Y5与分蘖明显高于主茎的品种1383这2个品种为试验材料。
1.3 试验方法
1.3.1 调控分蘖高度基因研究 DNA提取。采用CTAB法提取32份高粱基因组DNA,提取的DNA使用超微量核酸蛋白测定仪(scandrop100)测定其纯度及浓度。
目标片段PCR扩增、测序和序列比对。将32份高粱材料分别用4个SNP引物进行PCR扩增(表2),反应产物琼脂糖凝胶电泳分离,将单一扩增产物送北京擎科生物科技公司测序,利用DNAMAN v7.0软件进行序列比对分析。

表2 引物序列和信息Tab.2 Sequence and information of the primers
1.3.2 基因表达分析 RNA提取。利用TRIzol法分别提取K35-Y5和1383不同时期的叶、根和茎中总RNA,提取的总RNA使用超微量核酸蛋白测定仪(scandrop100)测定其纯度及浓度。
反转录合成cDNA。分别以K35-Y5和1383在枝梗分化期、主茎孕穗期、分蘖孕穗期、主茎开花期、分蘖开花期的主茎和分蘖的叶、根和茎共50份组织的总RNA为模板,使用Aidlab公司反转录试剂盒(TUREscript 1st Stand cDNA SYNTHESIS Kit)进行反转录,500 ng总RNA建立20 μL体系,程序设置为42 ℃ 40 min,65 ℃ 10 min,反应后得到cDNA样品,-80 ℃保存备用。
实时荧光定量 PCR。选择高粱Actin基因作为内参基因,以高粱总RNA反转录得到的cDNA模板进行RT-qPCR扩增。其中,所用内参及目的基因引物序列如表3所示。PCR反应体系为:cDNA 1 μL,2× SYBR® Green Supermix(1×)5 μL,上、下游引物(200 nmol/L)各0.5 μL,ddH2O 3.0 μL。PCR 反应条件为:95 ℃预变性3 min;95 ℃变性10 s,退火30 s(表3),39个循环。
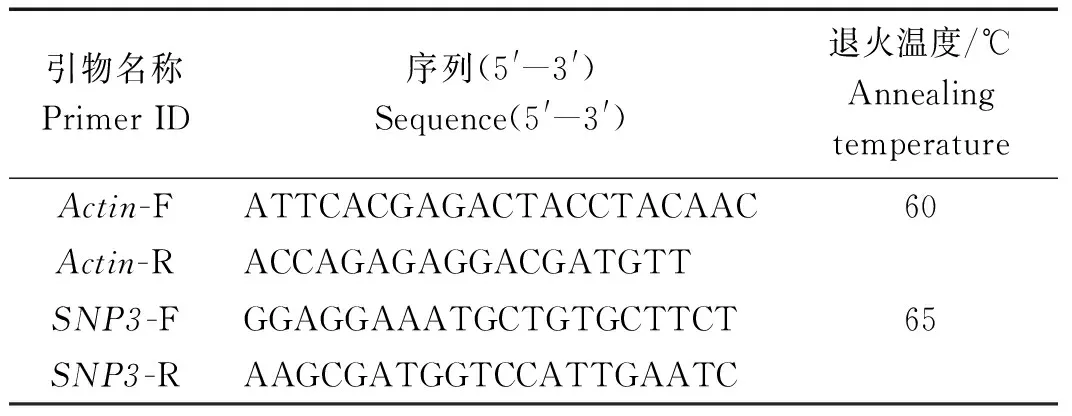
表3 引物信息Tab.3 Information of the primers
1.4 数据分析
每个cDNA样品重复3次试验,实时荧光定量扩增所得Ct值,以Actin为内参,利用2-ΔΔCt计算各个样品的基因相对表达量[26-27]。
2 结果与分析
2.1 基因测序比对与功能分析
山西农业大学高粱研究所分子育种研究室前期用分蘖与主茎株高一致的高粱品系K35-Y5与分蘖明显高于主茎的高粱恢复系1383杂交获得F2分离群体,选择高粱基因组Sorbi1.25作为参考基因组(ftp://ftp.ensemblgenomes.org/pub/plants/release-25/fasta/sorghum_bicolor/),采用BSA和SLAF技术将高粱分蘖与主茎株高一致的基因定位到Chr9上,在关联区域发现4个非同义突变的SNP。本研究选择已经测序完成的高粱基因组v3.1.1作为参考基因组(https://phytozome.jgi.doe.gov/pz/portal.html#!bulk?org=Org_Sbicolor),对原数据进行对比分析,结果显示,包含原4个SNP突变位点,分别对应到基因组v3.1.1的位置(表4)。

表4 SNP标记信息Tab.4 Information of SNP
将32份高粱材料分别用4个SNP引物进行验证,利用DNAMAN 7.0序列分析软件将测序结果进行序列比对分析发现,引物SNP1、SNP2、SNP4测序结果未能对应参试品种表型,只有引物SNP3的测序结果与表型对应,株高整齐一致的品种No.1~14和K35-Y5在第55 892 216 bp处均为T,分蘖高于主茎的品种No.15~30和1383在第55 892 216 bp处为G,SNP3测序结果与田间表型完全一致(图1),证明SNP3为调控高粱分蘖高度位点,其位点所属基因为Sobic.009G213300.1.v3.2,命名为SbTH。

图1 引物SNP3测序部分序列比对分析结果Fig.1 Comparative analysis result of SNP3 sequences
利用Psipred(http://bioinf.cs.ucl.ac.uk/psipred/)对SbTH进行二级结构分析发现,由于一个碱基位点突变,引起甲硫氨酸(Met)变成精氨酸(Arg)(图2),二级结构中的α-螺旋(Helix)结构区域延伸了一个氨基酸(图3);利用pfam(http://pfam.xfam.org/)数据库对该蛋白进行保守结构域分析,结果显示,该突变氨基酸位于水解酶_4(Hydrolase_4)保守区域内。

图2 SbTH基因的突变位点Fig.2 Mutation site of SbTH
图3 SbTH基因的二级结构Fig.3 Secondary structure of SbTH
2.2 SbTH基因在高粱不同组织的表达分析
2.2.1SbTH基因在高粱叶片中的表达 通过qRT-PCR分析K35-Y5和1383叶的基因相对表达量,在K35-Y5叶中,SbTH基因在主茎和分蘖的相对表达量呈现先上升再下降的趋势,在主茎孕穗期的表达量最高,虽表达量不同,但变化趋势基本一致;在1383叶中,SbTH基因的表达提前,主茎和分蘖的相对表达量均在枝梗分化期最高,随后呈现下降的趋势,变化趋势也基本一致(图4)。

图4 SbTH在K35-Y5和1383叶中表达趋势Fig.4 SbTH expression trend in K35-Y5 and 1383 leaves
2.2.2SbTH基因在高粱根系中的表达 分析K35-Y5和1383根的基因相对表达量,结果显示(图5),K35-Y5基因相对表达量呈现先上升再下降的趋势,主茎孕穗期开始极速上升,在分蘖孕穗期表达量最高,到开花期又极速下降;1383基因相对表达量相差不是很大,先上升,在分蘖孕穗期表达量增高,随后下降再上升,在分蘖开花期达到最高。在K35-Y5与1383根中,SbTH基因相对表达量不同,但趋势基本一致。
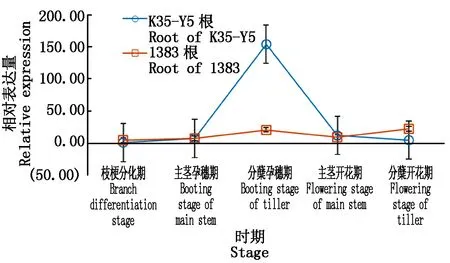
图5 SbTH在K35-Y5和1383根中表达趋势Fig.5 SbTH expression trend in K35-Y5 and 1383 roots
2.2.3SbTH基因在高粱茎秆中的表达 通过qRT-PCR分析K35-Y5和1383茎的基因相对表达量,结果显示(图6),K35-Y5主茎与分蘖基因相对表达量在主茎开花期时有显著差异,主茎相对表达量高,分蘖相对表达量低;1383主茎和分蘖基因相对表达量在各时期的变化无显著差异,主茎的基因相对表达量呈现先上升再下降的趋势,在主茎开花期达最高,分蘖的表达量逐渐升高,在分蘖开花期达最高。1383主茎和K35-Y5主茎基因相对表达量在各时期的变化趋势一致;分蘖的相对表达量在主茎开花期时有显著差异,K35-Y5分蘖相对表达量低,1383分蘖相对表达量高。

图6 SbTH在K35-Y5和1383茎中表达趋势Fig.6 SbTH expression trend in K35-Y5 and 1383 stems
茎秆高度由节数、节间长和穗柄长构成,那么SbTH基因是调控了节数与节间长度还是穗柄长,通过分别比较K35-Y5与1383主茎和分蘖的节间长度、节数与穗柄长,结果发现,K35-Y5主茎株高105 cm,分蘖株高104 cm;1383主茎株高145 cm,分蘖株高176 cm;K35-Y5分蘖数2~3个,1383分蘖数0.6~1.0个。无论是株高整齐一致的K35-Y5还是分蘖高于主茎的1383,穗柄长度基本一致,株高整齐一致的K35-Y5分蘖和主茎节数均为7个,节间长基本一致(图7-A、B),分蘖高于主茎的1383主茎节数13节,分蘖节数12节,分蘖比主茎少1节,但节间长度明显长于主茎(图7-C、D)。因此,分蘖高于主茎的原因主要是分蘖节间比主茎增长,从这一结果看,SbTH基因基本不影响穗柄长度,而是与节间的伸长有关。另外,1383分蘖比主茎少1节,说明其分蘖时期偏晚,晚分蘖是否会造成分蘖高于主茎仍需进一步研究。

A、C.K35-Y5和1383全株;B、D.K35-Y5和1383节间与穗柄长(标尺为5 cm)。A,C.Whole plant of K35-Y5 and 1383;B,D.Internodes and panicle handle of K35-Y5 and 1383 (Ruler is 5 cm).图7 K35-Y5和1383主茎和分蘖Fig.7 The length of main stem and tillers of K35-Y5 and 1383
3 结论与讨论
高粱的4个株高基因有3个已完成了克隆,Dwl基因(Sobic.009G229800)编码参与细胞增殖调节的高度保守蛋白质[22-23],Dw2基因(Sobic.006G067700)编码蛋白激酶[24],Dw3基因同源于玉米的Br2基因,编码ABCB1生长素转运体[19-20]。本研究验证了SNP3位点调控分蘖高度,所属基因为Sobic.009G213300.1.v3.2,命名为SbTH,位于水解酶_4保守区域,有别于其他株高基因。Johnson[28]和Bononi等[29]研究认为,α/β水解酶参与脂质信号、代谢和调节,包括生长和休眠等。但SbTH是如何调控高粱分蘖生长尚需深入研究。
从SbTH基因在不同组织的表达量来看,不同发育时期表达量有巨大差异,枝梗分化期和主茎孕穗期时叶片中表达量高,分蘖孕穗期时根中表达量高,到主茎开花期和分蘖开花期时茎中表达量高,SbTH基因的表达峰值依次在叶、根和茎中高表达,可以看出,SbTH基因影响高粱植株的生长并且基因的表达是从叶移动到根、再移动到茎,这与高粱生长的源库平衡基本一致[30]。
从SbTH基因在叶、根、茎的表达量来看,在叶和根中虽然表达量有差异,但趋势基本一致;在茎中,分蘖高于主茎的1383表达量和趋势也基本一致,只有K35-Y5(分蘖高度与主茎一致)在主茎开花期时呈反向表达,主茎表达量达到最高,同期的分蘖表达量降到最低。分析认为,SbTH在K35-Y5分蘖中表达量低,控制了分蘖的过度生长,形成主茎与分蘖高度一致的株型,SbTH在1383分蘖中表达量高,促进分蘖的生长,使得分蘖比主茎高,因此,SbTH基因在主茎开花期差异表达是影响分蘖株高生长的关键。
本研究中,从SbTH基因在1383与K35-Y5这2个材料的表达来看,在叶片中表达量出现较大差异,表达时期也明显不同,1383在枝梗分化期表达量最高,且主茎表达量比分蘖高;K35-Y5中SbTH基因的表达延后至主茎孕穗期,且分蘖的表达量比主茎高,这可能是由于该基因在1383中表达时期较早,主茎叶片正值发育时期,主茎比分蘖有更强的活力,导致SbTH基因主茎比分蘖表达量高,而K35-Y5表达量峰值在主茎孕穗期,此时,主茎的叶片数已经固定,叶面积生长达到最大,而分蘖的叶还处于生长时期,从而导致分蘖的表达量比主茎高。SbTH基因高表达会促进高粱植株的生长发育,K35-Y5分蘖茎在主茎开花期SbTH基因表达量低,抑制了茎的生长,形成表型整齐一致的株型。 Kim等[31]研究认为,高分蘖杂交种比低分蘖杂交种具有更高的早期分蘖出现频率。本研究中,K35-Y5比1383分蘖力强,K35-Y5比1383的分蘖时间早,1383的分蘖时期偏晚导致1383分蘖比主茎少1节,晚分蘖是否会造成分蘖高于主茎仍需进一步研究。
Chen等[32]从BTx623 EMS诱变群体中鉴定了一个NAB1(non-dormantaxillarybud1)基因突变体,nab1突变体增加了分蘖,降低了株高。利用nab1突变体与ShangZhuang扫帚高粱构建的F2群体将NAB1定位于6号染色体Sobic.006G170300基因上。NAB1编码一个类胡萝卜素裂解双加氧酶7(CCD7),主要表达在节和分蘖基部。Govindarajulu等[33]用拟高粱和高粱Tx7000(S.propinquum×S.bicolorTx7000)种间杂交衍生的重组自交系(RIL)群体,将QTL分析与转录组学相结合解析分蘖的遗传基础,发现6 189个基因在分蘖伸长过程中有差异表达,如DRM1等,表明这些基因是分蘖伸长的关键调控因子。本研究结果表明,SbTH基因在高粱不同组织中均有表达,只有在茎中K35-Y5(分蘖高度与主茎一致)主茎开花期时呈反向表达,主茎表达量达到最高,同期的分蘖表达量降到最低,分析认为,SbTH在K35-Y5分蘖中表达量低,控制了分蘖的过度生长,SbTH基因在高粱分蘖生长发育过程中发挥重要作用。
Quinby等[7]在4个确定的Dw1、Dw2、Dw3、Dw4的基因座上鉴定了等位基因变异,认为这些基因座通过改变茎节间长度来调节植株高度。Yamaguchi等[23]研究认为,Dw1降低了节间的细胞增殖活性,Dw1和Dw3的协同效应有助于提高抗倒伏性和机械收获。 Brown等[21]对378个高粱自交系使用关联作图描述dw3突变的表型效应,表明dw3突变与节间长度缩短和顶端伸长有关。Li等[34]研究认为,Dw3影响旗叶以下的茎高度。本研究发现,调控高粱分蘖高度的基因SbTH在主茎开花期差异表达是影响分蘖节间伸长的关键,这对提高品种整齐度、适宜机械化生产具有重要意义。
本研究发现,调控高粱分蘖高度的基因SbTH,所属基因为Sobic.009G213300.1.v3.2,位于水解酶_4保守区域。SbTH基因在主茎开花期差异表达是影响分蘖株高生长的关键,分蘖茎中表达量低,控制了分蘖节间的过度生长,形成株高整齐一致的株型。该基因的发现为研究高粱分蘖高度调控分子机制奠定了基础,对提高高粱品种整齐度、选育适宜机械化生产高粱品种具有重要的意义。
