柴达木大肥菇发酵过程中酚酸抗氧化活性变化
2022-09-21邓莹莹焦迎春蒋涛
邓莹莹,焦迎春,蒋涛
(青海大学农牧学院,青海 西宁 810016)
柴达木大肥菇[Agaricus bitorquis(Quél.)Sacc.]不仅富含脂肪、氨基酸、蛋白质、矿物质、膳食纤维等营养成分[1],而且含有酚酸、三萜及多糖等活性物质[2],具有极高的开发价值。酚酸类化合物是指苯环上的氢原子被酚性羟基取代的一类化合物[3],存在多种形式,主要以化学键与其他物质相结合,极少为游离态。酚酸作为一种重要的天然酚类化合物,不仅是人类从自然界直接摄入的重要酚类物质,还是多酚重要的体内代谢物,具有抗炎、抗病毒、抗癌、抗氧化[4]等生物活性,可抑制肥胖、提高免疫力、改善情绪等。目前对于柴达木大肥菇研究多集中于柴达木大肥菇的生物学特性和驯化[5]菌丝体、子实体的成分分析[1]及多糖酚酸提取工艺优化[4,6],对其酚酸类活性成分缺乏较深入的研究[2]。
生物机体内的氧自由基经过不饱和脂肪酸的过氧化可引起细胞受到损伤,进而引起细胞新陈代谢和生理反应无法正常进行,使机体加速衰老。若机体内氧化和抗氧化反应失衡则容易引起氧化应激现象,这是一种负面效应,严重时会加快机体衰老和各种疾病的产生,人体内的自由基多数为活性氧自由基(reactive oxygen species,ROS),包括含氧自由基、超氧阴离子自由基和过氧化物。活性氧的生成伴随生物的正常代谢过程,但过量的活性氧会对细胞造成氧化损伤,危害人体健康[7],因此,开发出具有低副作用天然无毒的抗氧化剂十分重要。抗氧化活性是酚酸最常见生物活性之一,酚酸以清除ROS为基础实现抗氧化活性[8],决定抗氧化活性的主要因素是酚羟基数目及其在化学结构中的位置。柴达木大肥菇在发酵过程中,可以分泌出大量酚酸,存在于发酵液和菌丝体中,分别对其提取即得胞外酚酸和胞内酚酸。陈雅楠等[4]研究发现柴达木大肥菇胞内酚酸具有较强的抗氧化活性。Leonard等[9]发现发酵通常会增加酚酸含量及其抗氧化活性;孟歌等[10]发现药食用真菌灵芝的自身生长情况、次级代谢产物分泌及还原能力等因素直接影响其抗氧化能力,表明大型食用真菌在液态发酵过程会提高其分泌出酚酸的含量,影响其抗氧化能力。因此,柴达木大肥菇做为食用真菌发酵过程中产生的酚酸也应具有类似的抗氧化活性。
综上所述,以柴达木大肥菇为研究对象,采用液体发酵的方法,测定发酵过程中的生物量、酚酸含量和酚酸的5种抗氧化指标,探究柴达木大肥菇在发酵过程中的抗氧化活性变化以及胞内、胞外酚酸抗氧化活性的差异,为柴达木大肥菇酚酸中抗氧化功能因子的开发利用奠定理论基础。
1 材料与方法
1.1 材料与试剂
柴达木大肥菇菌种:青海大学农牧学院食品工程实验室提供;三氯化铁、铁氰化钾、麦芽糖、果糖、蛋白胨、硫酸镁、氯化钙、无水乙醇、戊二醛(均为分析纯):湖北永阔科技有限公司。
1.2 仪器与设备
鼓风干燥箱(ZXRD-B5110):上海智城分析仪器制造有限公司;数控超声波清洗仪(KQ5200DE):昆山市超声仪器有限公司;紫外可见分光光度计(UV-1780):日本岛津仪器有限公司;台式高速离心机(H/T16MM):湖南赫西仪器装备有限公司;恒温振荡培养箱(BS-1EA):常州国华电器有限公司;扫描电镜(EVO 18):北京欧波同光学技术有限公司。
1.3 方法
1.3.1 液体种子培养
无菌条件下,从斜面培养基中取出黄豆大小的菌块,接入250 mL的三角瓶中,于25℃、100 r/min恒温振荡培养箱中培养,直到出现细小均匀的菌丝球且菌液澄清透明为最佳[11]。
1.3.2 液体发酵培养
在无菌条件下,以体积分数10%的接种量接入250 mL锥形瓶中,置于25℃、100 r/min恒温振荡培养箱中培养[11]。
1.3.3 柴达木大肥菇除糖工艺
菌丝体除糖:菌丝体与蒸馏水以1∶20(g/mL)放入烧杯,70℃、45 Hz超声20 min后提取抽滤。
发酵液除糖:发酵液和无水乙醇以体积比1∶4放入烧杯,40℃、45 Hz超声30 min后提取多糖,将上清液浓缩至原体积,即为胞外酚酸[4]。
1.3.4 柴达木大肥菇胞内酚酸的提取
将除糖后的菌丝体与90%乙醇以1∶20(g/mL)料液比放入烧杯,60℃、45 Hz超声30 min后提取抽滤,滤液即为胞内酚酸。
1.3.5 菌丝体生物量的测定
根据参考文献[12]测定菌丝体干重(g)。菌丝体生物量计算公式如下。
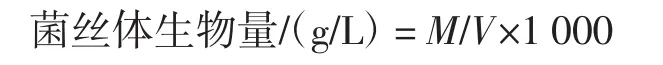
式中:M为菌丝体干重,g;V为发酵液总体积,mL。
1.3.6 菌丝体扫描电镜观察
将培养1 d~6 d的柴达木大肥菇菌丝体于2.5%戊二醛溶液中避光固定12 h。用0.1 mol/L磷酸盐缓冲液冲洗2次,然后分别用30%、50%、70%、80%、90%和100%的乙醇溶液脱水2次[13]。
1.3.7 酚酸含量测定
制备没食子酸对照品溶液,分别取 0.1、0.2、0.4、0.6、0.8 mL没食子酸对照品溶液置于25.0 mL的容量瓶中,加入1.0 mL的无水乙醇、0.3% C12H25SO4Na溶液、0.6% FeCl3溶液和 0.9 mL 0.9% K3[Fe(CN)6]溶液,并用0.1 mol/L盐酸溶液定容,静置20 min,测定720 nm处的吸光度[4],得到标准曲线方程:y=1.267 8x-0.021 5,R2=0.996 4;确定酚酸的稀释倍数,根据上述方法测得相应的吸光度后计算相应的胞内、胞外酚酸浓度。
1.3.8 抗氧化指标测定
1.3.8.1 超氧阴离子自由基清除能力测定
根据参考文献[14]的方法测定超氧阴离子自由基清除能力。清除率计算公式如下。

式中:△A空白为用无水乙醇代替样品的反应体系的吸光度;△A样品为添加样品的反应体系的吸光度;△A对照为以超纯水代替邻苯三酚的反应体系的吸光度。
1.3.8.2 铁离子还原能力的测定
根据参考文献[15]的方法测定铁离子还原能力。
1.3.8.3 羟自由基清除能力测定
根据参考文献[10]的方法测定羟自由基清除能力。清除率计算公式如下。
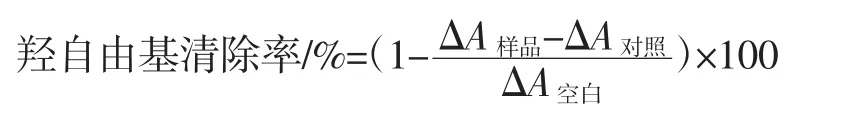
式中:△A空白为无水乙醇代替样品的反应体系的吸光度;△A样品为添加样品的反应体系的吸光度;△A对照为以蒸馏水替代H2O2的反应体系的吸光度。
1.3.8.4 2,2-联氮-二(3-乙基-苯并噻唑-6-磺酸)二铵盐阳离子 [2,2'-azinobis-(3-ethylbenzthiazoline-6-sulphonate)cation,ABTS+]自由基清除能力测定
根据参考文献[10]的方法测定ABTS+自由基清除能力。清除率计算公式如下。
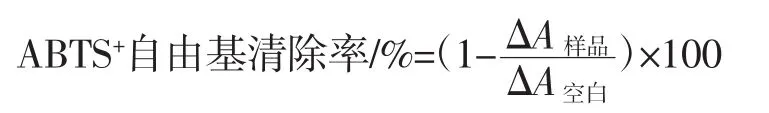
式中:△A样品为添加样品后溶液的吸光度;△A空白为未添加样品后溶液的吸光度。
1.3.8.5 1,1-二苯基-2-三硝基苯肼(1,1-diphenyl-2-picrylhydrazyl,DPPH)自由基清除能力测定
根据参考文献[16]的方法测定DPPH自由基清除能力。清除率计算公式如下。

式中:△A空白为无水乙醇替代样品的反应体系的吸光度;△A样品为添加样品后溶液的吸光度;△A对照为无水乙醇替代DPPH的反应体系的吸光度。
1.4 数据分析
所有试验数据用IBM SPSS Statistics 20分析显著性,用Origin 2018做图,显著水平设定为P=0.05,差异极显著水平设为P=0.01,相同小写字母表示差异不显著(P>0.05),不同小写字母表示差异显著(P<0.05)。
2 结果与分析
2.1 柴达木大肥菇发酵过程中菌丝体生物量变化
发酵过程中菌丝体生物量的变化见图1。发酵过程中菌丝细胞的形态变化见图2。
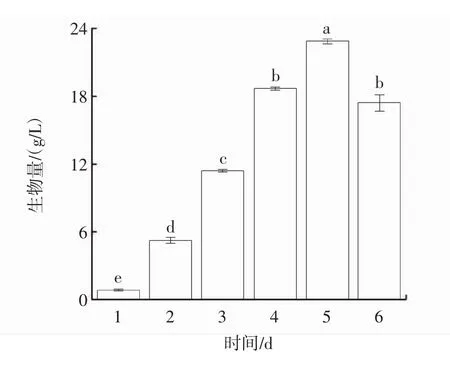
图1 发酵过程中菌丝体生物量的变化Fig.1 Variation curve of hyphae biomass
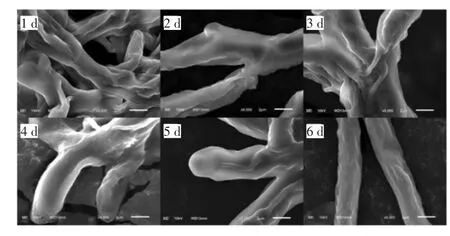
图2 发酵过程中菌丝细胞的形态变化(扫描电镜8 000×)Fig.2 Morphological changes of mycelium cells during fermentation(SEM 8 000×)
由图1可知,在培养的6 d内,菌丝体生物量呈先上升后下降的趋势,并在第5天达到峰值(22.91±0.21)g/L,较第1天提升了25.08倍。由图2可知,在发酵过程中,柴达木大肥菇菌丝球直径随着时间的延长呈先上升后趋于平稳的趋势。与鸡腿菇菌丝体生物量的试验结果趋势[17]类似,但由于真菌种类的差异,生物量达到峰值的时间不一致,随着发酵进程延续发酵液体系中溶解氧含量持续降低,菌体争夺养分、生长空间受限,菌体产量开始下降。
由图2可知,柴达木大肥菇在液体发酵过程中主要以圆球形态存在。在发酵初期,菌球较小,边缘为绒毛状,发酵液澄清透明,呈淡黄色,随着发酵的进行,菌球逐渐变大,菌丝逐渐变粗,并出现分支,发酵液逐渐浑浊、黏稠,6 d时,菌丝体老化,菌丝变细。可能是由于发酵后期时发酵液中营养物质减少,菌丝开始自溶。
2.2 柴达木大肥菇酚酸含量变化
胞内、胞外酚酸含量变化见图3。

图3 胞内、胞外酚酸含量变化Fig.3 Changes of intracellular and extracellular phenolic acid content
由图3可知,胞外酚酸含量始终高于胞内酚酸。在发酵过程中,胞内、胞外酚酸含量随着时间的延长呈先上升后平缓的趋势,并于第7天达到峰值,分别(6.73±0.05)、(10.11±0.05)g/L,较发酵第1天分别提高了 17.91倍、4.56倍,差异显著(P<0.05)。发酵前期菌丝体需一定的时间适应新的液体环境,故产生的酚酸较少。进入对数生长期后,菌丝体显著增长,酚酸含量开始增多,一直持续到稳定期,由于营养物质有限,从而导致酚酸含量增加速度变缓,最终趋于平稳。综上所述,选择发酵时间6 d作为一个发酵周期,完成对柴达木大肥菇发酵。
2.3 柴达木大肥菇发酵过程中胞内、外酚酸抗氧化活性变化
2.3.1 发酵过程中超氧阴离子自由基清除率变化
超氧阴离子自由基清除率变化见图4。
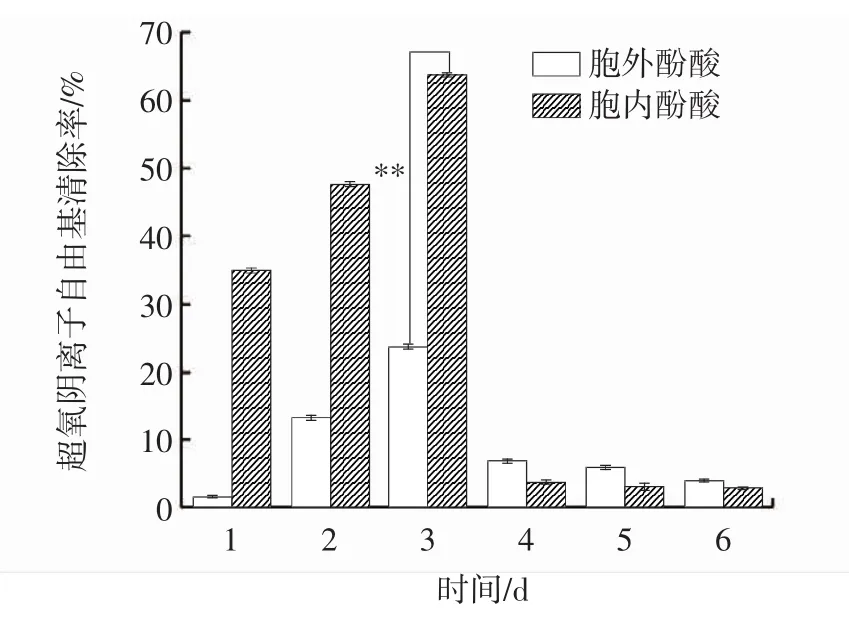
图4 发酵过程中胞内、胞外超氧阴离子自由基清除能力变化Fig.4 Changes of intracellular and extracellular superoxide anion scavenging ability during fermentation
超氧阴离子自由基(·O2-)是基态氧接受一个电子后形成的第一个氧自由基,可以转化为其他自由基如单线态氧、H2O2和羟自由基,其氧化能力很强,对组织产生氧化损伤而引发各种疾病[18]。因此,常用样品对超氧阴离子自由基清除能力来反映其抗氧化活性。由图4可知,1 d~3 d时胞内酚酸超氧阴离子自由基清除能力强于胞外酚酸,4 d~6 d时胞外酚酸超氧阴离子自由基清除能力强于胞内酚酸。在发酵过程中,胞内、胞外酚酸超氧阴离子自由基清除率随着时间的延长呈先升高后下降最后趋于平稳的趋势,并于第3天达到最大值,分别为 63.90%、23.95%,差异极显著(P<0.01),较发酵第1天分别提高了1.82倍、13.09倍。与桑黄液体培养中超氧阴离子自由基清除能力趋势类似[19]。
2.3.2 发酵过程中铁离子还原能力变化
发酵过程中胞内、胞外酚酸铁离子还原能力变化见图5。
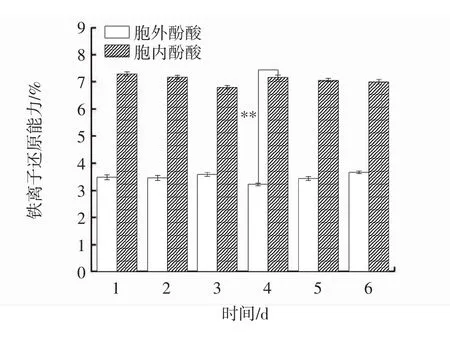
图5 发酵过程中胞内、胞外酚酸铁离子还原能力变化Fig.5 Changes of intracellular and extracellular phenolic acid iron ion reduction ability during fermentation
铁离子还原能力的测定,实质上是检验样品电子供应能力的过程。铁离子还原能力可以参与多种抗氧化反应,可以破坏自由基链式反应和过氧化物的降解,铁离子还原能力可用来评价样品总的抗氧化活性,且铁离子还原能力越强,酚酸抗氧化能力就越强[20]。由图5可知,胞内酚酸铁离子还原能力始终强于胞外酚酸,并在第4天达到最大差值,为3.93%,在第4天达到峰值,差异极显著(P<0.01)。在发酵过程中,胞内、胞外酚酸铁离子还原能力变化稳定,分别为7.08%、3.49%。与桦褐孔菌液体发酵培养的1 d~6 d胞内外多酚铁离子还原能力变化趋势一致[21]。
2.3.3 发酵过程中羟自由基清除率变化
发酵过程中胞内、外酚酸羟自由基清除能力变化见图6。
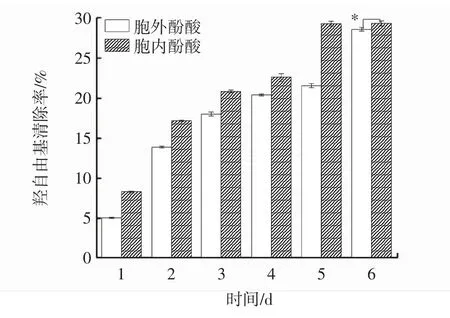
图6 发酵过程中胞内、胞外酚酸羟自由基清除能力变化Fig.6 Changes of intracellular and extracellular scavenging ability of phenolic acid hydroxyl radical during fermentation
羟自由基是一种活性氧,可以破坏细胞膜与蛋白质等生物分子发生反应,造成组织破坏和细胞死亡,这也是引起衰老和其他疾病的重要因素[22]。因此,可用羟自由基的清除能力评价样品的抗氧化能力。由图6可知,胞内酚酸羟自由基清除能力始终强于胞外酚酸。在发酵过程中,胞内、胞外酚酸羟自由基清除率随着时间的延长呈上升趋势,并于第6天达到峰值,分别为 29.35%、28.61%,差异显著(P<0.05),较发酵第 1天分别提高了2.52倍、4.61倍。与灵芝液体培养2 d~6 d的羟自由基清除能力变化趋势类似[10]。
2.3.4 发酵过程中ABTS+自由基清除率变化
发酵过程中胞内、胞外酚酸ABTS+自由基清除能力变化见图7。
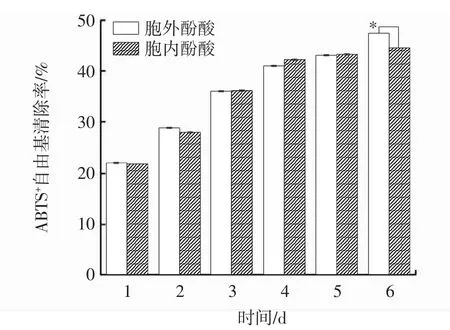
图7 发酵过程中胞内、胞外酚酸ABTS+自由基清除能力变化Fig.7 Changes of intracellular and extracellular ABTS+scavenging ability during fermentation
ABTS是一种水溶性的自由基引发剂,在清除ABTS+自由基反应中,抗氧化剂通过抑制蓝绿色的ABTS+自由基的产生来评价抗氧化性。当遇到具有抗氧化活性的物质时,ABTS+自由基会被还原,溶液颜色变浅,吸光值降低;吸光值越小,说明自由基清除剂的清除能力越强,以此可评价物质的抗氧化活性[23]。由图7可知,胞内、胞外酚酸ABTS+自由基清除能力基本相同。在发酵过程中,胞内、胞外酚酸的ABTS+自由基清除率随着时间的延长呈上升趋势,并于第6天达到峰值,分别为 44.52%、47.32%,差异显著(P<0.05),较发酵第1天分别提高了1.03倍、1.14倍。与桑黄液体发酵培养2d~6d过程中ABTS+自由基清除能力趋势一致[19]。
2.3.5 发酵过程中DPPH自由基清除率变化
发酵过程中胞内、胞外酚酸DPPH自由基清除能力变化见图8。
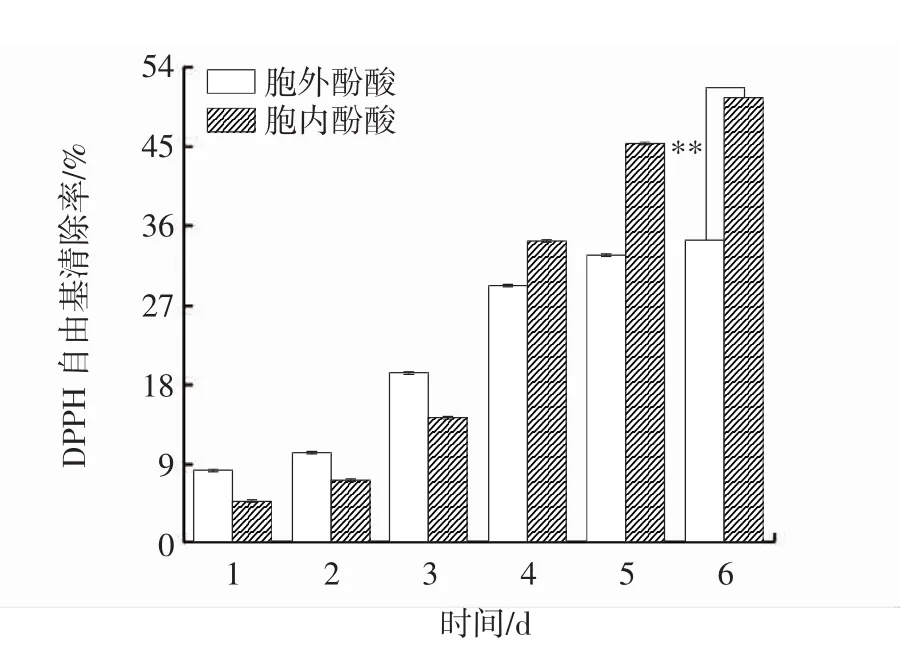
图8 发酵过程中胞内、胞外酚酸DPPH自由基清除能力变化Fig.8 Changes of DPPH scavenging capacity of intracellular and extracellular phenolic acids during fermentation
DPPH是一种稳定的顺磁化合物,若自由基清除剂存在时,则DPPH接受一个电子或氢原子,单电子被配对而形成稳定的DPPH-H化合物,使其乙醇溶液从深紫色变为黄色,变色程度与配对电子数(自由基清除活性)成正比[24]。因此通过DPPH的清除率可反映出酚酸的抗氧化活性。由图8可知,在发酵前期,胞外酚酸DPPH自由基清除能力大于胞内酚酸,在发酵后期,情况相反。在发酵过程中,胞内、胞外酚酸DPPH自由基清除能力随着时间的延长呈上升趋势,并于第6天达到峰值,分别为50.54%、34.39%,差异极显著(P<0.01),较发酵第1天分别提高了10.52倍、3.17倍。与栎生桑黄液体发酵过程中DPPH自由基清除能力变化趋势一致[25]。
3 结论
本文以发酵过程中的柴达木大肥菇为研究对象,利用超声波辅助法提取胞内、胞外酚酸,采用ABTS+自由基、DPPH自由基、羟自由基、超氧阴离子自由基清除能力及铁离子还原能力5个抗氧化体系,综合分析柴达木大肥菇在发酵过程中胞内、胞外酚酸抗氧化活性变化。结果显示:在发酵过程中菌丝体生物量呈现先上升后下降的趋势,在第5天时菌丝体生物量达到最大,较第1天提升25.08倍;酚酸含量变化呈现先上升后趋于平稳的趋势,胞外酚酸含量高于胞内酚酸含量,在发酵后胞外酚酸含量提升了4.56倍,胞内酚酸含量提升了17.91倍;柴达木大肥菇发酵过程中胞内、胞外酚酸具有良好的抗氧化活性,且胞内酚酸的抗氧化能力高于胞外酚酸的抗氧化能力。所得柴达木大肥菇胞内酚酸对ABTS+自由基、DPPH自由基、超氧阴离子自由基、羟自由基的最高清除率及最高铁离子还原能力分别为44.52%、50.54%、63.90%、29.35%、7.17%,分别是最低值的2.03倍、11.52倍、20.61倍、3.52倍、1.05倍,胞外酚酸对4种自由基的最高清除率及铁离子还原能力分别为47.32%、34.39%、23.95%、28.61%、3.67%,分别是最低值的2.14倍、4.17倍、14.09倍、5.61倍、1.13倍。柴达木大肥菇发酵过程中胞内、胞外酚酸体现了良好的抗氧化能力,可以作为天然抗氧化剂进一步开发利用。
