长白山阔叶红松林紫椴种群结构与动态特征
2022-07-30张金峰葛树森梁金花李俊清
张金峰,葛树森,梁金花,李俊清,*
1 北京林业大学森林资源生态系统过程北京市重点实验室, 北京 100083
2 吉林省林业调查规划院, 长春 130022
种群结构与动态是种群生态学研究的核心内容[1—3]。其中,种群结构能够反映种群内个体的年龄、大小、数量及其配置情况,同时也能反映种群数量动态与变化趋势[4—6]。种群动态主要是研究种群数量在时间和空间上的变动规律[7]。数量动态指数、基于静态生命表所构建的存活曲线、生存函数以及时间序列预测模型是种群统计学的重要应用方法[8—9],不仅可以用来分析植物种群目前的生存状态及其与环境间的相互关系,还可以评估历史干扰,并对种群未来发展趋势进行预测,对植物资源的保护及利用具有重要意义[10—11]。
紫椴(Tiliaamurensis)为椴树科(Tiliaceae)椴树属(Tilia)的高大落叶乔木,是我国东北地区阔叶红松林的主要优势树种,在维持该区森林群落稳定方面发挥着重要作用[12]。该种为国家二级重点保护野生植物,也是重要的蜜源植物[13]。由于长期以来的过量采伐和利用,加之其种子可育力低,天然更新能力弱,致使野生紫椴种群数量和质量逐年下降,蓄积大幅减少[14—15]。目前,关于紫椴的研究多集种于群落结构特征[16]、种子特性[17]、形态结构[18—19]、遗传育种[20]以及流蜜情况[13]等,而对其种群结构与数量动态的研究尚未见报道。鉴于此,本文以分布于长白山北坡阔叶红松林的紫椴种群为研究对象,利用种群统计学应用方法,分析种群结构特征及其生存现状,探究种群动态发展趋势,以期为野生紫椴种群的保护提出适应性对策,进而为长白山区森林生态系统恢复和多样性保护提供科学依据。
1 材料与方法
1.1 研究区概况
研究区位于吉林省东南部长白山国家级自然保护区北坡海拔1150 m以下的阔叶红松林(127°33′—128°16′E, 41°42′—42°45′N)。该区受温带大陆性山地气候影响,冬季漫长凛冽,夏季短暂多雨;年平均温度3.6℃,最热月(7月)均温为19.7℃,最冷月(1月)均温为-18.6℃;年平均降水量在600—900 mm,主要集中于7—8月份;土壤为暗棕色森林土,土层厚度20—100 cm。除紫椴外,该区乔木树种有红松(Pinuskoraiensis)、蒙古栎(Quercusmongolica)、水曲柳(Fraxinusmandshurica)、色木槭(Acermono)、春榆(Ulmusjaponica)等;灌木主要包括毛榛(Corylusmandshurica)、东北茶藨(Ribesmandschuricum)、东北溲疏(Deutziaamurensis)、刺五加(Acanthopanaxsenticosus)和卫矛(Evonymusalatus)等;草本层主要有蚊子草(Filipendulapalmate)、耧斗菜(Aquilegiaviridiflora)、狭叶荨麻(Urticaangustifolia)、荨麻叶龙头草(Meehaniaurticifolia)和猴腿蹄盖蕨(Athyriummultidentatum)等。
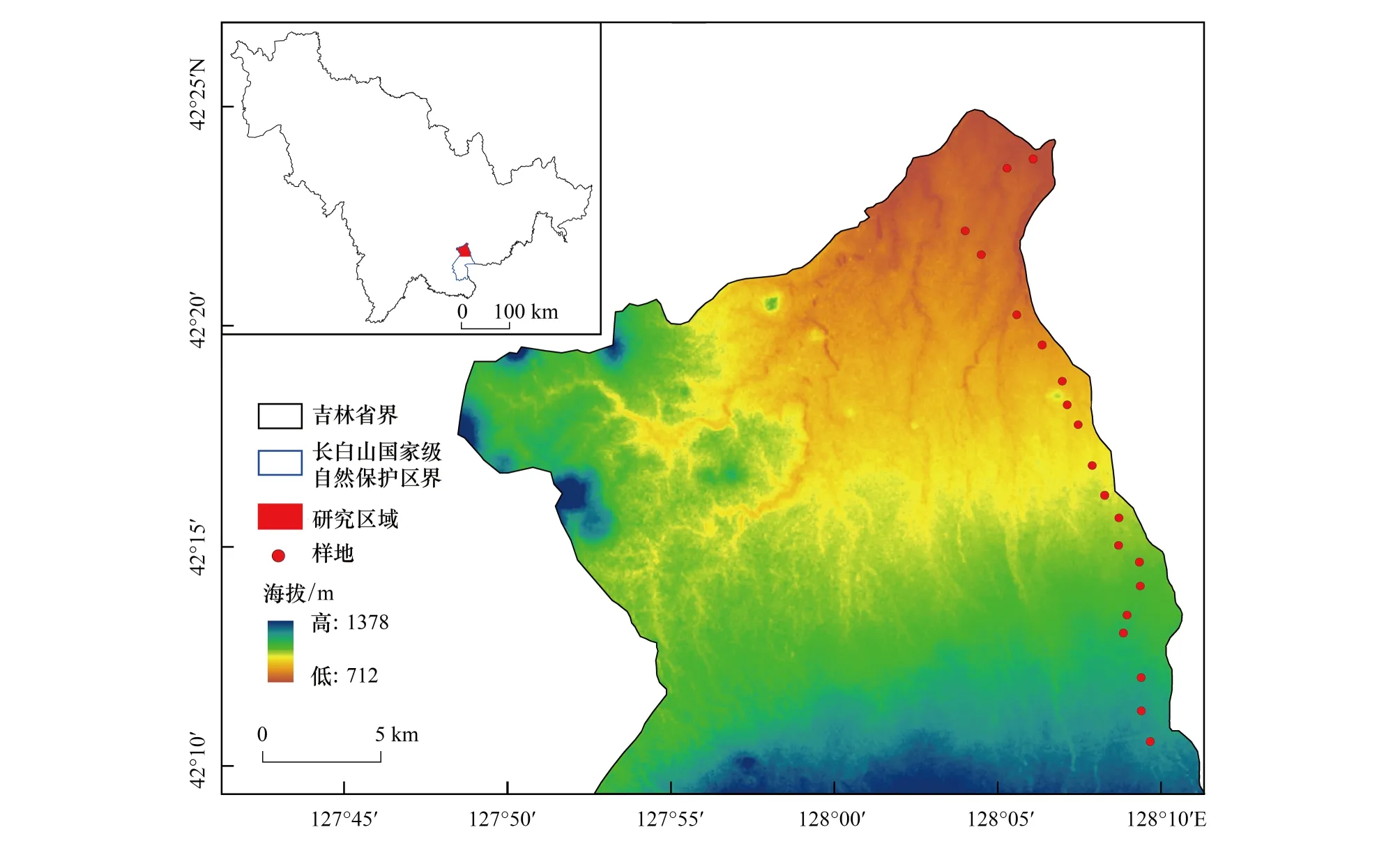
图1 长白山阔叶红松林紫椴种群样地分布图Fig.1 The sample distribution map of Tilia amurensis population in broad-leaved Korean pine forest of Changbai Mountain
1.2 研究方法
1.2.1样地设置
2020年6—8月,以长白山北坡紫椴分布的阔叶红松林(海拔730—1150 m)为研究区域,采用样地调查法,沿海拔梯度每间隔20 m设置1个样地,样地大小为30 m×30 m,共20个样地(图1),总样地面积18000 m2。对样地内DBH≥1 cm的木本植物进行每木检尺,记录胸径、冠幅、株高和数量等;同时记录每个样地的海拔、经纬度、坡度和坡向等生境因子。
1.2.2紫椴种群径级结构划分
紫椴为国家二级重点保护野生植物,不宜利用解析木或生长锥进行年龄测定。因此,本文依据同一生境内,同种植物的龄级和径级对环境具有相同的反应规律这一特点[21],采用径级代替龄级的方法来分析种群动态变化。根据样地内调查情况和紫椴的生长规律,并参考董点等[18]、许梅等[22]对紫椴种群龄级结构的划分标准,将其划分为13个龄级,具体如下:0 cm<Ⅰ≤5 cm、5 cm<Ⅱ≤10 cm、10 cm<Ⅲ≤15 cm、15 cm<Ⅳ≤20 cm、20 cm<Ⅴ≤25 cm、25 cm<Ⅵ≤30 cm、30 cm<Ⅶ≤35 cm、35 cm<Ⅷ≤40 cm、40 cm<Ⅸ≤45 cm、45 cm<Ⅹ≤50 cm、50 cm<ⅩⅠ≤55 cm、55 cm<ⅩⅡ≤60 cm、XIII>60 cm。同时,将13个龄级分为4个阶段,即幼龄阶段(Ⅰ—Ⅳ龄级)、中龄阶段(Ⅴ—Ⅷ龄级)、成龄阶段(Ⅸ—ⅩⅡ龄级)和老龄阶段(XIII龄级)。
1.2.3种群动态量化
采用陈晓德[23]种群年龄结构动态量化分析法定量描述紫椴种群结构:
式中,Vn为种群内两相邻龄级间个体数量变化动态指数;Vpi为无外部干扰情况下整个种群的数量动态变化指数;Sn、Sn+1分别为第n、n+1龄级种群个体数。若考虑未来的外部干扰时,整个种群的数量动态变化指数还与各龄级内的个体数Sn以及年龄级数量(K)有关:
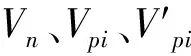
1.2.4紫椴种群静态生命表
以紫椴种群各径级的株数为依据,编制其静态生命表。生命表主要包括以下参数:1)x,龄级;2)Ax,x龄级内存活个体数。因本研究中各龄级的个体数相差较大,生命表中会出现死亡率为负值的情况,不符合静态生命表编制的假设,故采用匀滑技术[2,11]对数据进行处理,得到ax,即匀滑修正后x龄级内的存活个体数;3)lx,x龄级开始时的标准化存活个体数(一般转化为1000),lx=ax/a0×1000;4) lnlx,lx的自然对数;5)dx,从x到x+1龄级的标准化死亡数,dx=lx-lx+1;6)qx,x龄级到x+1龄级间的死亡率,qx=dx/lx×100%;7)Lx,x龄级到x+1龄级间平均存活个体数,Lx=(lx+lx+1)/2;8)Tx,x龄级到超过x龄级的个体总数,Tx=∑Lx;9)ex为生命期望,表示第x龄级的个体所能存活的平均年数期望,ex=Tx/Lx;10)Kx,各龄级内的消失率,Kx=lnlx-lnlx+1;11)Sx,种群存活率,Sx=lx+1/lx。
1.2.5存活曲线、死亡率曲线和消失率曲线拟合
本研究以标准化存活数lx为纵坐标,以胸径表示的龄级x为横坐标,绘制紫椴种群存活曲线。根据存活曲线的变化特征,可将其分为3种类型[24]:Deevey-Ⅰ型,该类型种群中大多数个体都能活到其平均生理寿命,早期死亡率较低,当达到平均生理寿命时,短期内几乎全部死亡;Deevey-Ⅱ型,该类型种群各龄级死亡率大致相同;Deevey-Ⅲ型,该类型种群中幼龄个体死亡率较高,随后死亡率低且稳定。为了判断紫椴种群符合上述哪类存活曲线,本文参考Helt和Loucks的数学模型[25],采用指数方程Nx=N0e-bx和幂函数方程Nx=N0e-b分别检验存活曲线类型。
以龄级x为横坐标,死亡率qx、消失率Kx为纵坐标,分别绘制种群死亡率曲线和消失率曲线。
1.2.6生存分析
应用生存函数S(i)、累计死亡率函数F(i)、死亡密度函数f(ti)和危险率函数λ(ti)进行种群的生存分析:
S(i)=S1×S2×S3…×Si
F(i)=1-Si
式中,Si为存活率;hi为龄级宽度。
1.2.7种群数量时间序列预测
本文参考肖宜安等[26]的研究方法,采用时间序列分析中的一次平均推移法对紫椴种群年龄结构进行模拟和预测:
式中,n为龄级,t为需要预测的未来年限(本研究以龄级表示),Xk为当前k龄级的种群大小,Mn(1)表示种群经历未来t个龄级时间后n龄级的预测值。本研究应用此模型,以紫椴种群的实际调查数据中各龄级对应的个体数为基准年数据,预测紫椴种群各龄级经过2、3、4、5、6、7、8、9、10个龄级时间后的数量。
本研究所有统计分析在Excel 2007和SPSS 21进行,用Origin 2019软件绘图。
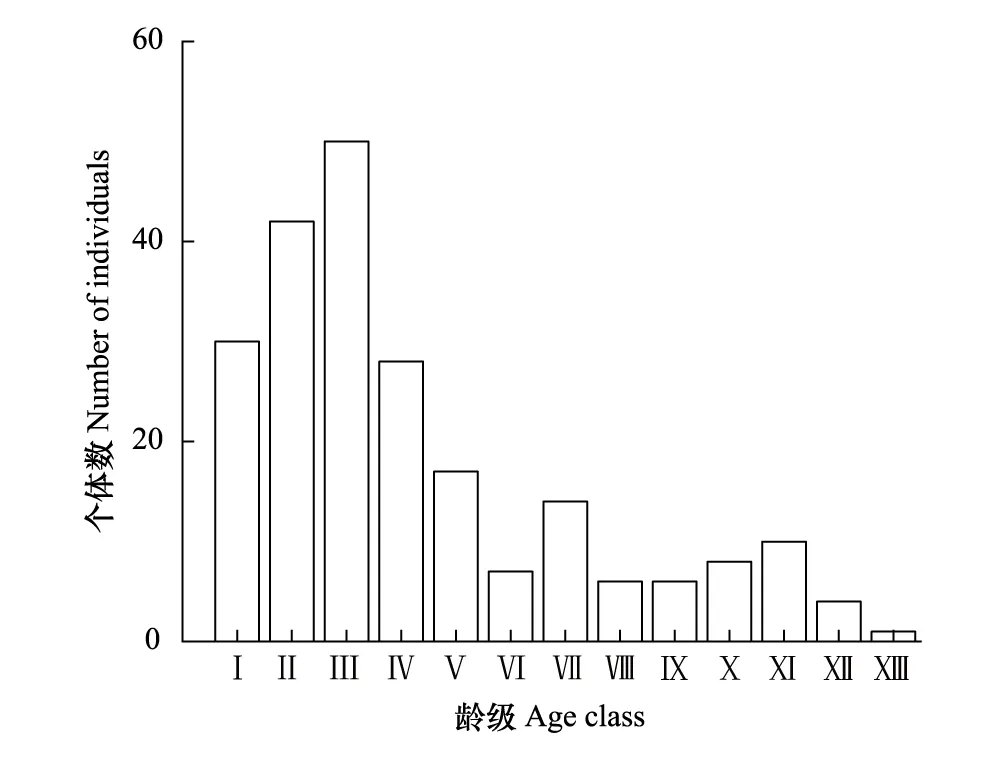
图2 紫椴种群径级结构Fig.2 Age structure of Tilia amurensis population 根据胸径(DBD, Diameter at breast height)大小,将其划分为13个龄级(Ⅰ: 0 cm
2 结果与分析
2.1 紫椴种群分布现状及结构特征
本次共调查到紫椴个体223株,最大胸径112.34 cm。由图2可以看出,种群的龄级结构均呈不规则金字塔型,Ⅲ龄级个体数最多,为50株,占种群个体总数的22.42%;Ⅰ—Ⅳ龄级个体数之和占个体总数的67.26%,说明幼龄个体数较多;Ⅴ龄级后个体数量降低趋势明显,表明幼龄个体很难通过环境筛;成龄(Ⅸ—ⅩⅡ龄级)个体数为28株、老龄(Ⅷ龄级)个体数仅为1株,二者占种群个体总数的13.00%。
2.2 紫椴种群数量动态
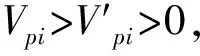
2.3 紫椴种群静态生命表及存活曲线
2.3.1静态生命表分析
由表2可以看出,紫椴种群不同龄级个体数差别大,幼龄(Ⅰ—Ⅳ龄级)个体数量较多,表明幼树数量充足,种群具有增长潜力。随龄级增大,种群的标准化存活数(lx)逐渐减少。期望寿命ex反映了各龄级内个体的平均生存能力,紫椴种群生命期望值总体呈先增加后降低的趋势。其中,Ⅴ龄级种群具最大生命期望,反映出幼龄阶段(Ⅰ—Ⅳ龄级)植株成功进入中龄阶段(Ⅴ—Ⅷ龄级)后生存能力有所增强。Ⅸ龄级后,由于受种群个体生理衰老等因素的影响,紫椴种群期望寿命随龄级增加逐渐下降,显示种群逐步到达生理寿命。

表1 紫椴种群动态变化指数
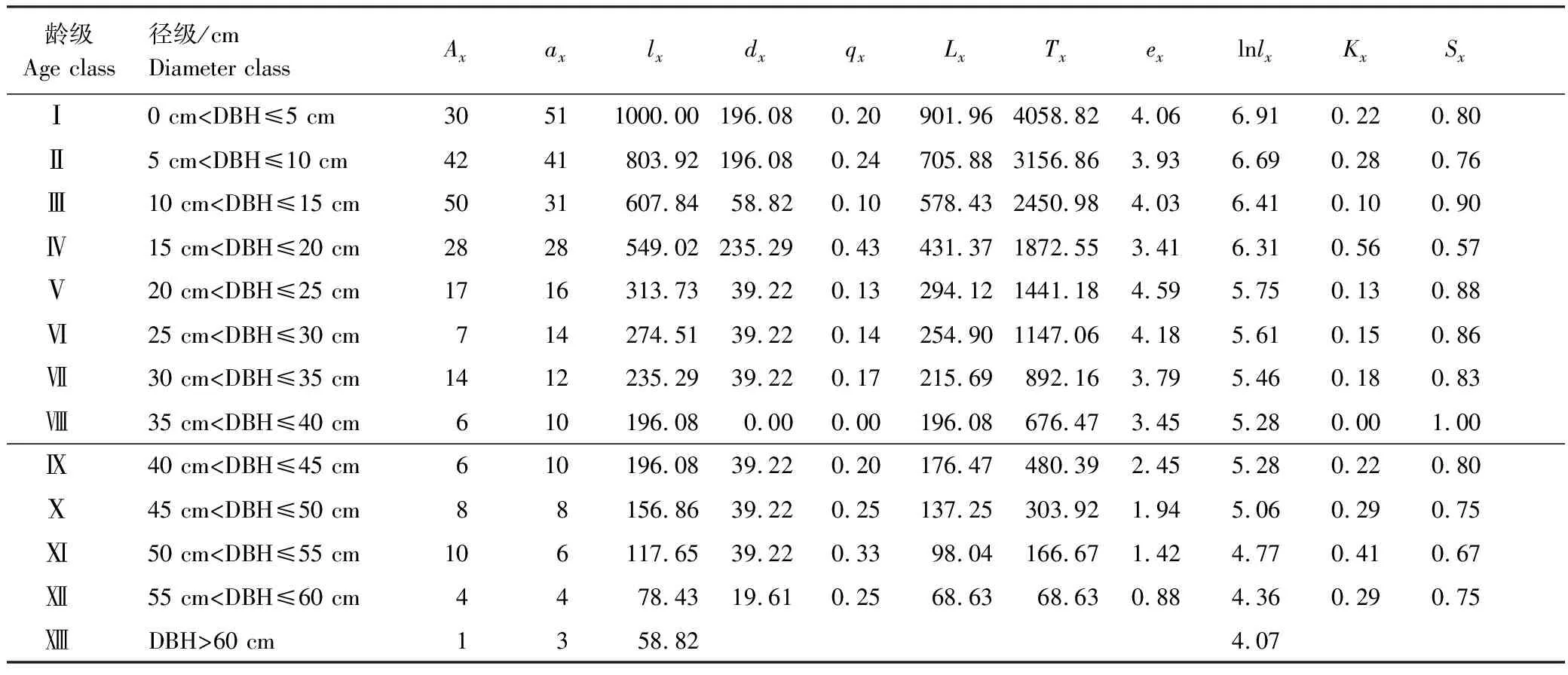
表2 紫椴种群静态生命表
2.3.2死亡率和消失率曲线分析
种群的死亡率和消失率反映了种群数量随龄级增加的动态变化。图3显示,阔叶红松林内紫椴种群死亡率(qx)和消失率(Kx)曲线变化趋势总体一致,随着龄级的增加均呈现出连续性先增后降的复杂动态变化趋势,结合表1以及种群动态量化分析的结果可知,该种群处于不稳定的状态。此外,qx和Kx均在Ⅳ龄级达最大植(0.56和0.43),说明在该龄级种群数量发生锐减。
2.3.3存活曲线分析
存活曲线能够反映种群个体在各径级的存活过程以及种群数量动态变化特征,图4显示,紫椴种群数量在Ⅰ—Ⅳ龄级下降缓慢,在Ⅳ—Ⅴ龄级急剧下降,Ⅴ龄级后下降又较为平缓,呈现出“凹-凸-凹”的变化趋势。采用2种数学模型方程对存活曲线进行检验(表3),两种模型的拟合结果均达极显著水平,但幂函数模型R2和F值均大于指数函数,表明该种群存活曲线符合Deevey-Ⅲ型,反映出幼龄期植株死亡率高。模型检验的结果与存活曲线的变化趋势基本一致。
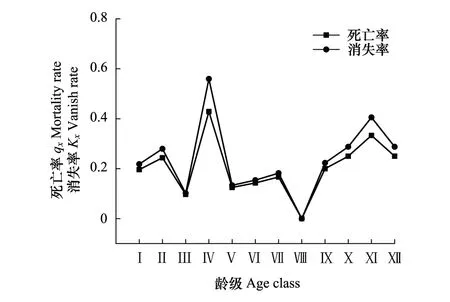
图3 紫椴种群死亡率和消失率曲线Fig.3 Mortality and vanish curves of Tilia amurensis population
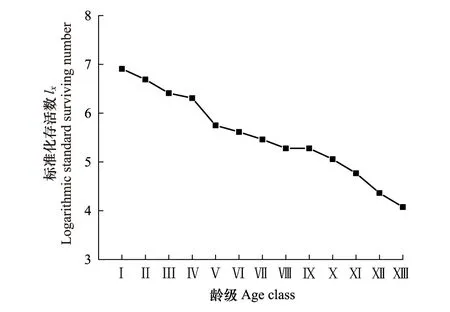
图4 紫椴种群存活曲线Fig.4 Survival curves of Tilia amurensis population

表3 紫椴种群存活曲线的检验模型
2.4 紫椴种群的生存分析
由图5可以看出,随着龄级的增加,紫椴种群生存率S(i)呈单调递减趋势,累计死亡率F(i)呈单调递增趋势,二者互补。在第Ⅲ—Ⅳ龄级间,生存率和累计死亡率相等,表明种群在该龄级阶段达到平衡状态,尔后累计死亡率大于生存率,种群进入衰退期。在Ⅰ—Ⅷ龄级之间,两者升降变化剧烈,这可能跟水热条件和生长空间有限而导致的强竞争作用有关。Ⅷ龄级后,S(i)和F(i)函数均达极限值,说明种群存活压力由竞争转向生理衰老。
紫椴种群死亡密度函数f(ti)整体呈单调递减趋势(图6),在第Ⅰ—Ⅱ龄级间急剧下降,表明其个体数量在这个龄级阶段发生锐减。在第Ⅷ龄级出现最小值(0),这说明紫椴种群开始进入生理衰老期,种群开始衰退。相较于死亡密度曲线,危险率λ(ti)曲线呈“增加-降低-增加-降低”的动态趋势,且基本保持在0.3以下,说明种群稳定性差,种群在整个生长周期都面临着死亡风险。
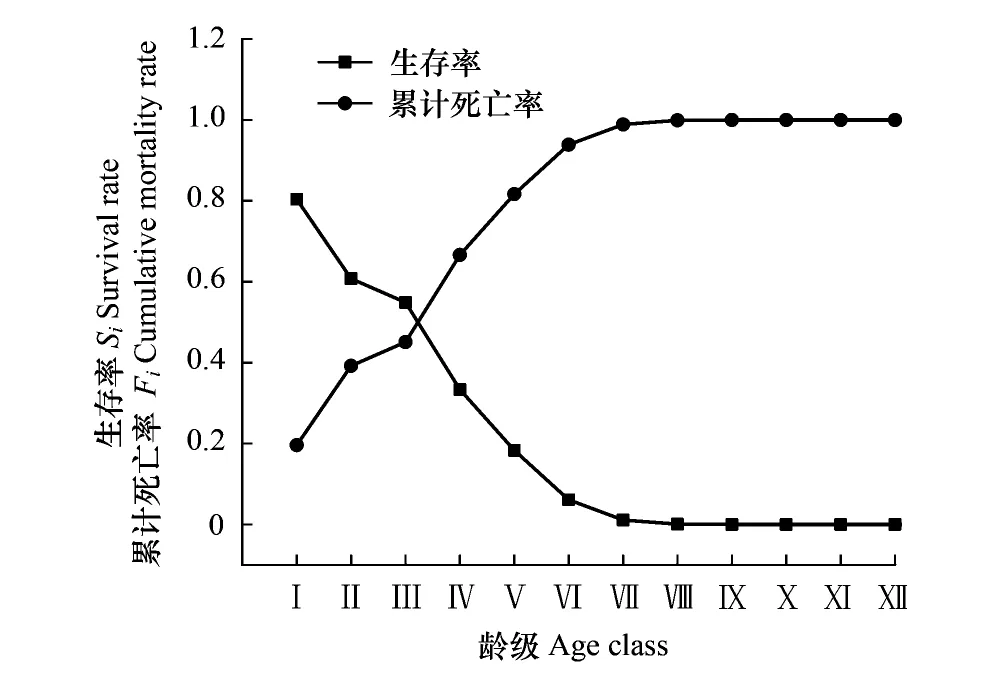
图5 紫椴种群生存率和累计死亡率曲线 Fig.5 Survival rate and cumulative mortality rate curves of Tilia amurensis population
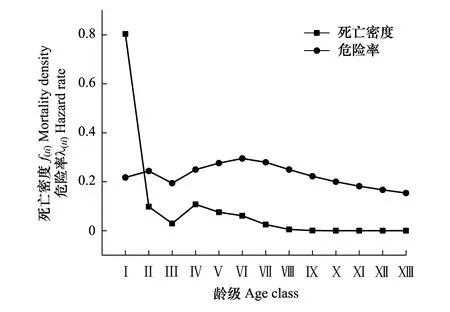
图6 紫椴种群死亡密度和危险率曲线 Fig.6 Mortality density and hazard rate of Tilia amurensis population
2.5 种群数量的时间序列分析
本文根据时间序列分析中的一次平均推移法,预测针对种群在未来2、3、4、5、6、7、8、9、10个龄级时间后的个体数量变化(表4)。结果显示,第Ⅱ龄级个体数量在经历2个龄级时间后下降14.28%;第Ⅲ龄级个体数量在3个龄级时间后下降18.00%;第 Ⅳ和Ⅴ龄级分别在3、4个龄级时间后达到种群数量的峰值,随后种群数量开始下降;上述结果共同表明尽管紫椴幼龄个体数量多,但难以过度到中龄阶段。Ⅵ、Ⅶ、Ⅷ、Ⅸ、Ⅹ、Ⅺ、Ⅻ、龄级个体数在未来总体呈现增加的趋势,说明紫椴种群未来的增长主要依靠中、成和老龄个体。然而,随龄级增加,Ⅵ—龄级植株个体数量平均增长速率依次降低(分别为19.94%、18.58%、14.80%、12.63%、11.79%、9.69%、6.93%、4.47%),这可能跟生理衰老有关。
3 讨论
3.1 紫椴种群年龄结构
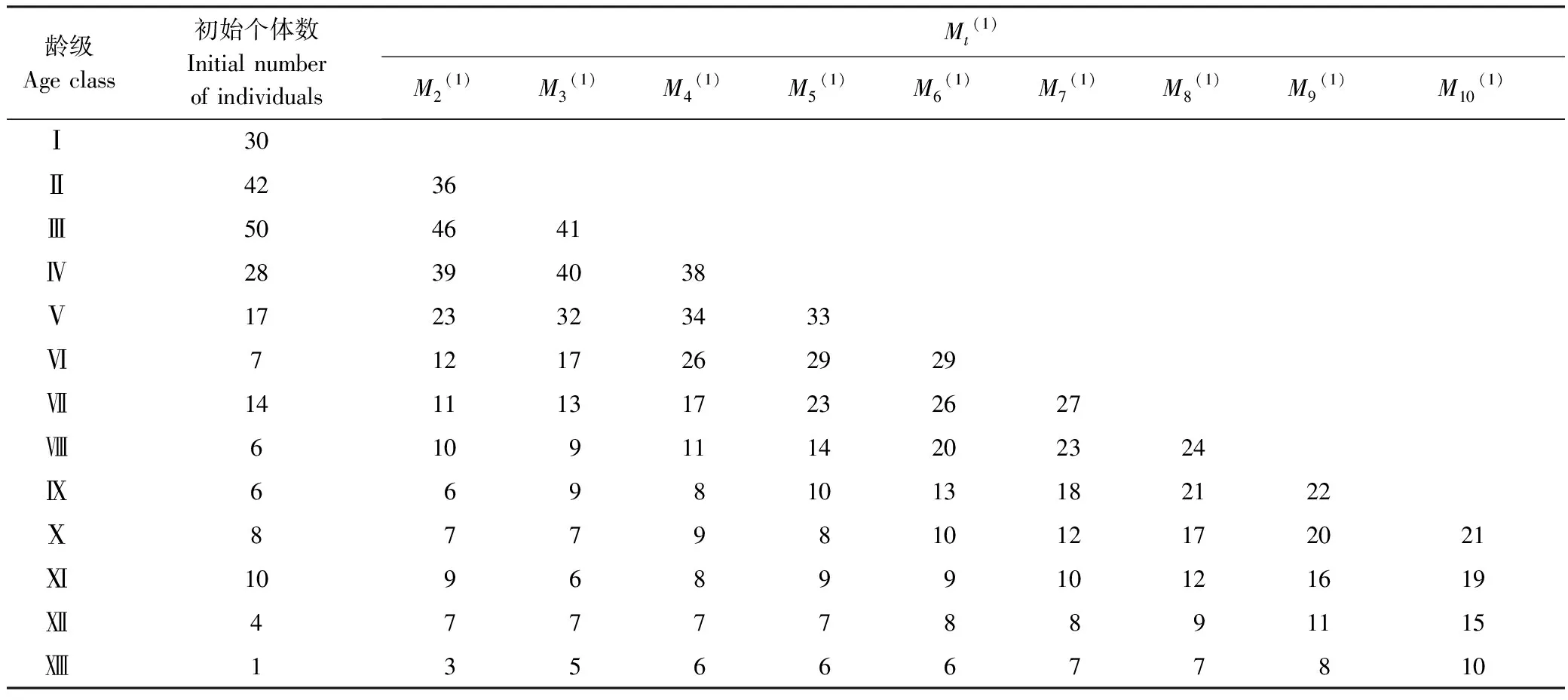
表4 紫椴种群数量动态时间序列预测
3.2 紫椴种群的动态趋势
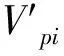
生存函数能够更直观、具体地反映种群增长与衰亡的动态过程[7]。研究发现,随龄级增加,累积死亡率函数单调上升,生存函数单调下降。其中,Ⅰ—Ⅲ龄级间种群生存率大于累计死亡率,这是因为幼龄个体数量充足,基本上能及时补充由竞争作用而导致的该龄级个体迁出或者死亡所造成的数量缺失;二者在Ⅲ—Ⅳ龄级间达到平衡状态,尔后进入衰退期,这一结果与陈科屹等[32]报道的该区红松种群在幼龄阶段的早期(Ⅰ龄级)就开始进入衰退期明显不同,这可能跟不同物种的生物学特性有关[12]。表明尽管二者同为阔叶红松林优势种,但红松幼龄个体对生存空间和资源条件的要求更为苛刻,种内竞争更为激烈;相反,小径级的紫椴个体需要的资源较少,竞争相对较弱[12]。尽管如此,紫椴由低龄级向中、高龄级生长的过程中对空间、光照、水分、养分的需求不断增大,有限的生存空间和资源条件成为紫椴种群在Ⅲ—Ⅳ龄级后进入衰退期的主要因素[3,5—6]。Ⅷ龄级后,紫椴种群累积死亡率和生存率曲线变化趋势较为平缓,表明此时种群的竞争压力减小并已成功占据一定的空间和环境资源,导致其死亡的主要原因转为生理衰老。死亡密度函数结果显示,Ⅰ—Ⅱ龄级间种群个体数量发生锐减,这与种群生命表分析的结果以及存活曲线趋于Deevey-Ⅲ型相对应。
从时间序列预测分析上来看,在未来2—10个龄级时间之后,紫椴种群Ⅱ—Ⅴ龄级个体数逐渐降低,说明幼龄个体难以过度到中龄阶段;Ⅵ—龄级个体数在未来逐渐增加,反映出幼龄个体一旦进入中、成和老龄阶段,则具有较强的生存能力,推测未来种群以大树、老树占主导地位,整个紫椴种群向高龄级个体集中。另外,本研究中,老龄个体数量虽然少但在自然环境中却占有较大空间。因此,从种群整体发展角度来看,少数老龄个体的消亡可促使幼龄个体进入上层空间,有利于种群更新和繁衍[6]。
3.3 紫椴种群的保护对策
根据本文的研究结果,建议对长白山紫椴种群的保护采取以下保护措施:1) 对幼龄死亡率高的紫椴种群展开实生苗繁育工作并种植回归原生境,但需人工疏苗减小幼苗密度,提高幼苗质量和存活率;同时,为防止远交衰退所带来的紫椴种群遗传没化,实生苗的种子必须从原生种群采集[33]。2) 针对该种群不稳定且在干扰情况下增长趋势不明显这一特点,应当采取封育保护措施,避免对其栖息地造成破坏。在封育保护的同时进行科学管理,适当疏伐非目标树种和病弱的紫椴个体。3) 实行就地保护,在紫椴现有分布区附近,选择相似生境作为保护点,人工育种后把苗木移栽到保护点进行人工种群的恢复。4) 目前已建立的长白山国家级自然保护区对该区物种保护起到了一定作用,但人为干扰因素仍较多,例如采集食用菌、药用植物等,加之近年来长白山地区旅游业逐渐兴起,应更加注重其保护情况,建议加强对保护区的管理力度,杜绝人为破坏。
