‘赤霞珠’与‘北红’正反交群体抗寒旱性能遗传特点
2022-01-21唐玄德王荣刘玉娟段伟梁振昌范培格
唐玄德,王荣,刘玉娟,段伟,梁振昌,范培格,3
(1. 宁夏大学农学院,银川 750021;2. 中国科学院植物研究所,北京 100093;3. 中国葡萄酒产业技术研究院,银川 750021)
近年来我国葡萄酒产业迅速发展,取得了可喜的成就[1-3]。据国际葡萄和葡萄酒组织(OIV)统计,2018年我国葡萄种植面积为87.5万 hm2,居世界第二;葡萄酒产量约为9.3亿升,居世界第十,约有6.9亿升需进口[4],因此,葡萄酒需求有很大的发展空间。我国酿酒葡萄主要分布在北方地区,以大陆性季风气候为主,冬季寒冷干旱,葡萄需要埋土防寒才能越冬。冬季埋土、春季出土不仅耗费大量劳动力,增加生产成本,同时也会造成树体的机械损伤,加剧种植地生态环境恶化[5],因此选育、推广抗寒旱的免埋土优质酿酒品种迫在眉睫。
有性杂交是果树最常见的育种方法,而果树杂交亲本在遗传上存在异质结合型,杂交性状广泛分离[6]。为了加快育种进程,提高育种效率,性状的遗传规律及亲本的选择至关重要[6]。研究认为,抗寒性遗传取决于亲本的冰点值(low-temperature exotherm,LTE)。一般LTE值越低,后代的抗寒性越强[5],且许多子代的LTE值高于亲中值,表明抗寒性存在超亲遗传现象。子代抗寒性与春季冻害等级不一致,但存在相关性。而在正反交群体中抗寒性状遗传特点的报道有限,目前仅有‘北醇’与‘康可’正反交及后代遗传规律的报道,指出二者正反交后,抗寒性不存在显著差异,也不存在细胞质遗传特点[7]。
本试验利用抗寒性具有明显差异的欧洲种‘赤霞珠’与山欧杂种‘北红’(Vitis vinifera×V. amurensis)构建的正反交群体后代为研究对象,对群体后代的冻害等级、萌芽率和LTE值等进行分析,旨在揭示群体后代的抗寒旱性状的遗传特点及正反交群体的差异,为今后葡萄抗寒旱杂交育种工作中合理选配亲本提供参考。
1 材料与方法
1.1 材料与试剂
试材为中国科学院植物研究所葡萄种质资源圃(北京)中2个群体共225个单株,‘赤霞珠×北红’杂交群体‘15-25’为正交,‘北红×赤霞珠’杂交群体‘14-2’为反交。正交为67个单株,反交为158个单株,‘赤霞珠’(埋土)与‘北红’(未埋土)为对照,2021年1月10日前后取杂交株系和对照品种树体上生长健壮枝条5条(每条5~8节),用保鲜膜包裹,放置于4 ℃的恒温冷库临时存放,用于含水量、冰点值及萌芽率调查。
1.2 仪器与设备
电子天平;数显糖度计(Digital Refractometer);风干烘箱;差热分析(different thremal analysis, DTA)系统包括电压控制冰箱和数据采集2个系统。
1.3 试验方法
1.3.1 枝条总含水量的测定
参照邹琦的烘干法[8]。取一年生5~8节生长健壮枝条,剪成10 cm的枝段,每3段一个重复,每个重复3个枝段,共9根,精确称重,记为W1;将其置于烘箱中,在105 ℃烘15 min杀青处理,然后置于80 ℃烘箱,烘至恒重,称重记为W2。
枝条总含水量(%)=[(W1-W2)/W1]×100
1.3.2 枝条自由水测定
参照潘炽瑞的方法[9]。取出10 mL型号的离心管,并放置于天平准确称重,记为W1;配制浓度65%蔗糖,分别加入离心管中,每管加5 mL,记为W2;将枝条避开芽眼用刀切成0.1 cm厚的薄片,准确称取0.5 g,3个重复,置于含有蔗糖溶液的试管中,再称重记为W3。扣紧离心管盖,置于暗处放置16~24 h,期间每隔3 h,均匀晃动,用Digital Refractometer折射仪测定各试管蔗糖溶液浓度。
枝条自由水含量(%)=[(W2-W1)×(C1-C2)/C2]/(W3-W2)×100
式中:W2-W1为糖液原质量(g);W3-W2为枝条鲜质量(0.5 g);C1为糖原液浓度(65%);C2为浸泡后糖液浓度(%)
枝条束缚水含量(%)=枝条总含水量-枝条自由水含量
束自比=束缚水含量/自由水含量
1.3.3 葡萄冬芽冰点值的测定
采用柴风梅等的方法[10]。取5~8节饱满芽,垂直芽体中轴线切下,带有0.5~1.0 mm厚的木质部组织,切面整齐。培养皿中加入1 mL蒸馏水,保证芽体湿润,电热板孔中放入吸水纸,加入150 μL蒸馏水,放入切好的芽体,每个孔中加入6~9个芽。盖上黑色海绵,保证芽体与电热模板紧密接触。固定好的电热板置于程序控制冰箱降温,降至-4 ℃保持1 h;-4~-40 ℃,降温速度为4 ℃·h-1,共需10 h,每隔15 s采集1次温度数据。植物组织低温时会集聚放出热量,因此导致温度升高,电压图会出现高峰,高峰出现时的温度为休眠芽的致死温度,即为冰点值(LTE)。
1.3.4 枝条萌芽率
萌芽率采用乔改霞的方法[11]。结合LTE试验,将取剩枝段,下截口斜剪,上截口平齐,用嫁接白色条带绑扎,逐一编号,4个重复,放置在黑色穴盘中扦插,穴盘内加满自来水,一周换两次水,下截口出现白色粘液后,清洗穴盘,重新加入自来水,直至枝条萌芽,萌芽期间加入自来水,保证盘中的穴孔满水。45 d后统计萌芽率。
萌芽率(%)=萌芽数/芽眼总数×100
1.3.5 田间冻害等级调查
春季田间植株萌芽后调查冻害等级,冻害级别及描述如下:
1级:无冻害。耐寒性极强,植株全部芽眼正常萌发,树体生长发育正常。伤流期间新剪口有伤流现象,芽眼和结果母枝未受冻,芽眼萌发整齐一致。
2级:轻微冻害。60%以上的芽眼正常萌发,伤流期间新剪口有伤流液,但伤流滴水较慢。受冻的一年生枝横截面有少量变褐,木质部轻微受冻,芽眼萌发推迟,且不一致。
3级:较严重冻害。伤流期间新剪口有伤流,但伤流滴水明显较少,整个植株部分芽眼和结果母枝受冻致死,受冻枝条形成层部分变褐,仅有部分芽眼萌发,且比正常萌发推迟,生长衰弱。
4级:严重冻害。伤流期间新剪口只有少量伤流,结果母枝和枝芽全部冻死,枝蔓形成层变黑。地上部芽眼无一萌发,只从根基部萌蘖。
5级:植株受冻害致死。伤流期中新剪口无伤流,主干基部隐芽无一萌发,整株枝蔓干枯死亡。
1.4 数据处理分析
用Microsoft excel 软件进行数据整理,用SPSS 22.0软件进行数据统计分析,用Excel、Origin 2018 64Bit软件作图。参照张文娥[12]的方法,采用隶属函数计算隶属值,取平均隶属值进行抗寒性分级。共5级:0.70~1.00为高抗;0.60~0.69为抗;0.40~0.59为中抗;0.30~0.39为抵抗;0~0.29为不抗。
隶属函数公式:U(Xi)=(Xi-Xmin)/(Xmax-Xmin)。如果某一项指标与判定结果为负相关,则为反隶属函数进行定量转换。
计算公式为:U(Xi)=1-(Xi-Xmin)/(Xmax-Xmin),式中,U(Xi)为隶属函数值,Xi为无性系某项指标测定值,Xmin和Xmax为所有参数无性系中某一指标的最大值和最小值[12]。
变异系数(%)=标准差/F×100;
超低亲率(%)=平均隶属度低于较低亲本的株数/正反交群体总株数×100;
超高亲率(%)=平均隶属度高于较高亲本的株数/正反交群体总株数×100;
遗传传递力(%)=F/MP×100;

式中F为杂交后代平均值;MP为亲中值;H2为广义遗传力;VG为遗传方差;MS为杂交后代组间方差,Mse为杂交后代组内方差。
2 结果与分析
2.1 正反交群体后代枝条含水量及萌芽率
由图1可以看出,亲本及正反交群体后代枝条总含水量保持稳定,大致呈一条水平线,分布在39%~51%。而正反交群体后代束缚水、自由水含量在群体中有差异,随着单株自由水含量的上升,其束缚水含量呈下降趋势。正反交后代束缚水含量均值差异明显,分别为20.5%、19.3%,且束缚水含量介于亲本之间,偏向抗寒性强的亲本‘北红’。

图1 亲本及后代水含量分布情况Figure 1 Water content distribution of parent and offspring
亲本‘赤霞珠’和‘北红’的总含水量、自由水含量没有显著差异,总含水量分别为44%、42%,自由水含量分别为25%、23%。但‘赤霞珠’的束缚水含量为14.6%,‘北红’的束缚水含量为20.5%,差异明显。‘赤霞珠’和‘北红’束自比也存在显著差异。
正交群体‘15-25’后代的束缚水含量高于‘北红’的单株有27株,占比约40%。反交群体‘14-2’后代的束缚水含量高于‘北红’的单株有34株,占比约22%。正交群体‘15-25’后代的束自比高于‘北红’的单株有21株,占比约31%。反交群体‘14-2’后代的束自比高于‘北红’的单株有34株,占比约22%。
后代株系的萌芽率不管是正交群体还是反交群体的均为100%。
2.2 正反交后代抗寒性遗传分析
亲本‘赤霞珠’LTE值为-16.20 ℃,属于低抗;‘北红’LTE值为-21.70 ℃,属高抗,两者的LTE值存在显著差异,分属不同的抗寒性等级。由表1可以看出,两个品种构建的正反交群体后代LTE值均呈不同程度的连续分布,分别为-24.40~-10.89、-24.68~-12.64 ℃。两个群体LTE值分布范围均大于强亲本,具有超亲遗传特点。正反交群体后代LTE均值分别为-21.01、-21.52 ℃,存在显著差异,整体趋向抗寒性较强的亲本‘北红’。变异系数分别为12.95%和9.52%,正交群体‘15-25’的变异系数大于反交群体‘14-2’,在同一环境条件下,正交群体的变异性高于反交群体。广义遗传力与遗传传递力没有显著差异,广义遗传力为0时,表明表型变异由环境影响造成;广义遗传力为1时,表型变异由基因控制,而正交群体后代在相同的环境条件下广义遗传力更多受基因影响。

表1 正反交群体LTE值统计Table 1 Statistics of orthogonal group LTE value
由表2可以看出,最大比例的群体后代均是介于亲本之间的类型,正交和反交群体LTE值高于强亲本值的比例分别为43.47%和41.25%,弱于弱亲本值的比例分别为8.69%和3.12%,抗寒性弱的亲本做母本后代中会出现更高比例抗性更弱的株系。

表2 正反交后代抗寒性偏向统计Table 2 Statistics of the anti-cold bias in offspring with positive and negative cross%
由图2可以看出,正交群体呈偏右峰单向正态分布,反交群体呈经典型正态分布。正反交群体后代LTE均呈离散分布。正交群体LTE集中分布区间为-23.8~-17.7 ℃,反交群体LTE集中分布在-23~-20 ℃,正交群体相对反交群体LTE更分散。
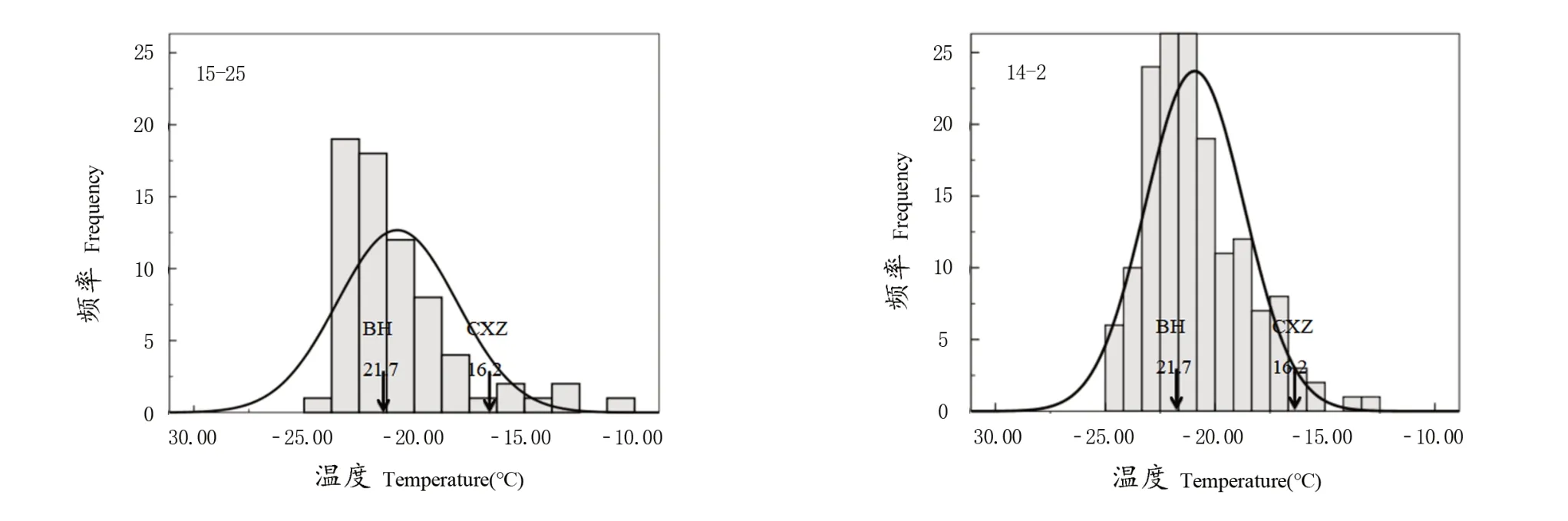
图2 正反交群体后代LTE值频率直方图Figure 2 Frequency histogram of orthogonal group offspring
2.3 群体田间冻害等级分析
由表3正反交群体后代冻害等级显著性分布看,两个群体后代最高比例等级均为1级,即无冻害植株,分别占比为50.70%和68.44%,反交群体高出近18个百分点。两个群体间后代3级和5级所占比例差异不大,而4级所占比例差异较大,正交群体比例高13.64个百分点。1、2级植株表现轻微冻害,60%以上的芽眼正常萌发,生产中对产量影响不大;反交群体1级(无冻害)和2级株系的比例高于正交20个百分点,说明反交群体‘14-2’后代具有更大比例的实际抗寒性能明显优于正交群体的株系。

表3 正反交群体冻害等级的分布Table 3 Distribution of sprouting grade in positive and negative groups%
2.4 含水量、LTE值、冻害等级相关性
根据群体后代株系含量差异的束自比与LTE值的相关性分析可看出(图3),两个群体的LTE值与束自比均具有相关性,一般束自比值高,LTE值低,抗寒性强。正交群体‘15-25’单株LTE值与束自比相关性为0.62,反交群体的相关性为0.74,但均未达到显著相关。

图3 束自比、LTE值相关性曲线Figure 3 Bound-free ratio and LTE value correlation curve
由图4可以看出,正交群体‘15-25’冻害等级对应的LTE中位数依次为-21.43、-21.82、-17.62、-21.62、-21.59 ℃,反交群体‘14-2’冻害等级对应的LTE值中位数依次为-21.35、-21.95、-21.85、-21.29、-20.58 ℃。由这两组数据可以看出,冻害等级与LTE值没有直接相关性,冻害等级与冬季枝条含水量也无直接相关性。

图4 不同冻害等级植株的冬芽LTE值分布Figure 4 Distribution of LTE of plants with different freezing grades
3 讨论与结论
利用高抗性的野生资源与优质的栽培品种杂交是选育具有优质高抗品种的重要方法[1,14,21-23]。本试验亲本‘北红’是山葡萄F1代,可抵御-21.83 ℃低温[13],在北方露地栽培可安全越冬。‘赤霞珠’是优良的欧亚种品种,但不抗寒,利用两个品种杂交期望选育出既有‘北红’的高抗寒性能,又具有‘赤霞珠’优良品质性状的新优品种。从群体后代的LTE值看,具有超亲遗传特点,而且两个群体株系在春季萌芽后田间表现出较高比例的无冻害植株比例,利用抗寒的山葡萄血统后代(F1)做亲本完全可以选育出抗寒性能强的株系,这与Wang等的结果一致[7]。
另外,有性杂交育种中父母本的配置至关重要。本试验利用抗寒性具有明显差异的‘赤霞珠’与‘北红’构建正反交群体,两个群体后代LTE均值不存在显著差异,但群体后代LTE值整体趋向抗寒性较强的亲本‘北红’,这与郭海林等[24]在结缕草属植物抗寒性试验中得出的遗传现象一致,也与王荣、柴风梅等[5,7,13]的研究结果一致。但试验发现,后代LTE值弱于弱亲本值的占比正交群体大于反交群体,因此抗寒性弱的亲本做母本(正交群体15-25)后代中会出现更高比例抗性更弱的株系。从群体后代冻害等级分布结果也得出,正交群体株系4级(地上部无一萌芽)所占比例相较反交群体高13.64个百分点,反交群体后代中1级(无冻害)和2级株系的比例高于正交群体20个百分点,说明选用抗寒性能强的亲本做母本,获得抗寒旱后代的株系概率更大。
试验结果发现,经过低温驯化后的1月份取样测得的枝条束自比与冬芽LTE值具有一定相关性,但未达到显著相关,而冬芽LTE值与春季田间冻害等级没有直接相关性。Wang等[7]利用7个杂交群体进行的试验也得出同样的结果。北京冬末春初,气温升高,但气候干旱,且常伴有大风天气,推测可能是气温回升后枝条中水分的变化影响最终的萌芽,具体不同类型基因型在越冬期间水分变化的差异及调控机制有待深入研究。
4 展望
有性杂交育种是抗寒旱优质育种的重要途径之一,明确不同正反交育种抗寒旱遗传特点,一方面可以解决我国北方葡萄埋土防寒根本问题,减少生产成本,提高市场竞争力。选用抗寒旱性能强的品种与品质优良的品种杂交选育适宜我国立地条件的新优品种,不仅从生产源头上解决北方葡萄冬季防寒埋土所带来的成本增高、生态恶化等问题,也可以丰富酿酒葡萄种质资源,实现品种与产品品质的多样化。另一方面,深入进行杂交群体后代及不同抗性种质资源抗寒旱调控机制研究,挖掘抗寒性状相关基因并加以验证,可为新品种的分子辅助筛选及定向育种提供新的技术手段和理论依据。
