葡萄根癌病病原菌中效应蛋白Tae4与其免疫蛋白Tai4的鉴定与分析
2022-01-21王鹏飞袁丽芳张庆田王海利慕茜王钦超韩金涛赵红军
王鹏飞,袁丽芳,张庆田,王海利,慕茜,王钦超,韩金涛,赵红军
(1. 山东省葡萄研究院,山东济南 250100;2. 山东省农药科学研究院,山东济南 250100;3. 山东师范大学,山东济南 250014;4. 龙口市省级农业高新技术产业开发区管委会,山东龙口 265701;5. 农业农村部华东都市农业重点实验室/山东省葡萄栽培与精深加工工程技术研究中心,山东济南 250100)
葡萄根癌病是一种细菌性病害,由葡萄根癌病病原菌引起。葡萄根癌病通常在夏季发病,对葡萄极具破坏性。根癌病病原菌可引起葡萄产量的下降,甚至导致葡萄藤的枯死[1]。最早被发现的葡萄根癌病病原菌是Agrobacterium vitis[1-2]。后来,根癌病病原菌Agrobacterium tumefaciens也从葡萄根部被分离出来,说明A. tumefaciens株系也能侵染葡萄,引发根癌病[2]。葡萄根癌病病原菌对葡萄的侵染可以分为3个步骤:第一步是致病菌侵入到葡萄的非原质体部位,汇集在葡萄的根部,再通过根部的伤口入侵[3-4];第二步,病原菌在葡萄的木质部定殖;第三步,通过葡萄木质部的液流散布到葡萄的各个器官和部位[5]。
根癌病病原菌的致病效应子VirA和VirG作为毒力蛋白可以促进根癌病病原菌对葡萄的侵染[1]。狭义上,效应子(又名效应蛋白)可以被定义为一类有利于致病菌侵染植物的蛋白质。当植物的致病菌分泌效应蛋白后,还必须将这些效应蛋白运输至植物寄主的精确亚细胞位置才能发挥功能,协助致病菌入侵植物。目前,大量的致病菌效应蛋白被报道,这些相关研究的增多使得效应子的特征被逐步分析和明确。研究显示,效应蛋白有如下特征:(1)具有信号肽;(2)无跨膜区域;(3)包含的氨基酸少于300个;(4)富含半胱氨酸;(5)具有已知的效应子结构域;(6)与已知蛋白功能无同源性等[6]。
六型分泌系统(The type VI secretion system;T6SS)是存在于根癌病病原菌等革兰氏阴性菌中的分泌系统,因为能分泌一些与致病相关的效应子,并与病原菌致病能力密切相关。然而,人们对六型分泌系统分泌的非致病效应子蛋白研究的较少。目前研究表明,六型分泌系统在细菌竞争中也发挥重要作用。有一种六型分泌系统分泌的非致病效应蛋白称为Tae4蛋白,在分泌Tae4蛋白后,会系统性地转运Tae4蛋白并注射到附近细菌的周质空间中。而这种非致病相关的Tae4效应蛋白具有酰胺酶活性,可以水解细菌的肽聚糖进而杀死附近的竞争细菌[7]。有的细菌可免受自身分泌Tae4蛋白的伤害,是因为其六型分泌系统可分泌一种Tai4,它是Tae4蛋白的免疫蛋白。目前有两种根癌病病原菌A. tumefaciens株系(ATU4346和ATU4347)被发现含有Tai4蛋白和Tae4蛋白[8]。由于这两个株系缺少完整的基因组,因此无法进行全基因组范围的Tai4蛋白和Tae4蛋白家族成员鉴定。本研究以具备完整基因组的18个A. tumefaciens和4个A. vitis株系为对象,鉴定这些菌株中含有Tai4蛋白和Tae4蛋白家族成员的确切数目,并分析该蛋白家族成员的特性。对其Tai4蛋白和Tae4蛋白的鉴定与分析可为将来研究如何借助Tae4对一些致病菌进行杀灭奠定基础。
1 材料与方法
1.1 葡萄根癌病病原菌基因组数据的获得
18个A. tumefaciens株系完整基因组以及全部蛋白序列信息下载于NCBI的A. tumefaciens基因组数据库(https://www.ncbi.nlm.nih.gov/genome/browse/#!/prokaryotes/177/),株系名称分别是:BIMB-1315G、1D1609、CFBP6623、12D1、CFBP6624、186、6N2、1D1460、EML4、CFBP7129、1D1108、6A、15955、FDAARGOS、BIM B-441、CFBP5499、CFBP5877、Q15_94。4个A.vitis株系完整基因组及其全部蛋白序列信息来源于NCBI的基因组数据库(https://www.ncbi.nlm.nih.gov/genome/browse/#!/prokaryotes/1215/),株系名称分别是:VAR06-30、VAR03-1、S4以及VAT03-9。
1.2 葡萄根癌病病原菌Tae4和Tai4蛋白家族成员鉴定
用HMMER软件,基于Tae4结构域(Tae4 domian;Pfam编号为PF14113)的隐马克夫模型(Hidden Markov Model,HMM),分别搜索18个A. tumefaciens株系和4个A. vitis株系基因组中全部Tae4蛋白,从而鉴定2种葡萄根癌病病原菌的Tae4蛋白家族成员。利用在线软件SMART(http://smart.embl-heidelberg.de/)确定蛋白是否具有完整的Tae4结构域,剔除无完整Tae4结构域的蛋白。
用HMMER软件,基于Tai4结构域(Tai4 domian;Pfam编号为PF16695)的隐马克夫模型,分别搜索18个A. tumefaciens株系和4个A. vitis株系基因组中全部Tai4蛋白,从而鉴定2种葡萄根癌病病原菌的Tai4蛋白家族成员。利用在线软件SMART(http://smart.embl-heidelberg.de/)确定蛋白是否具有完整的Tai4结构域,剔除无完整的Tai4结构域蛋白。
1.3 葡萄根癌病病原菌Tae4和Tai4蛋白家族成员亚细胞定位及信号肽预测
利用在线软件TargetP(http://www.cbs.dtu.dk/services/TargetP/)预测Tae4和Tai4蛋白家族成员的亚细胞定位;利用在线软件NLStradamus(http://www.moseslab.csb.utoronto.ca/NLStradamus/)分析核定位信号。设置参数为:Prediction Cutoff=0.6;用TMHMM2.0软件(http://www.cbs.dtu.dk/services/TMHMM/)分析跨膜结构域;利用在线软件SignalP(http://www.cbs.dtu.dk/services/SignalP/)分析信号肽。
1.4 多重序列比对与进化树分析
利用ClustalX 2.0软件对Tae4和Tai4蛋白家族成员进行蛋白多重序列比对分析;用MEGA 6.0软件构建系统发育树,方法为邻接法(Neighbor-Joining tree)。设置参数为:自举值(bootstrap value)被计算1000次。bootstrap value大于70的分支认为是1个组[9]。利用在线软件Topo-phylogeny(http://bar.utoronto.ca/Topophylogeny/)进行进化树的拓扑结构分析。
1.5 蛋白基序分析
利用在线软件MEME suite 5.3.3(https://meme-suite.org/meme/tools/meme)分析Tae4和Tai4蛋白家族成员的保守蛋白基序(motif)。设置参数为:motif发现数量=10,motif长度范围为10~200个氨基酸[9]。
1.6 蛋白三级结构分析
利用在线软件SWISS-MODEL(https://swissmodel.expasy.org/)分析蛋白质的三级结构。
1.7 蛋白互作网络分析
利用在线软件STRING(https://string-db.org/)分析Tae4和Tai4蛋白家族成员与其他蛋白的互作。数据库中A.tumefaciens蛋白互作数据来自株系CCNWGS0286。
1.8 选择压力分析
利用软件PAML包中的Code ml program(version 4.7)[10]分析Tae4蛋白编码基因在进化过程中受到的选择压力。利用PAML软件中的3个选择压力位点模型分析。利用位点模型M0(one ratio)分析整体受到的选择压力;利用位点模型M8(βandω)贝叶斯分析法(Bayesian Analysis)鉴定Tae4基因进化过程中的正向选择位点[11]。
2 结果与分析
2.1 葡萄根癌病病原菌中Tae4与Tai4的鉴定与蛋白性质分析
目前,NCBI公布的根癌农杆菌基因组数据包含18个A. tumefaciens和4个A. vitis的完整基因组。在18个A. tumefaciens株系的基因组中鉴定了14个Tae4基因,在4个A.vitis株系的基因组中鉴定了1个Tae4基因和1个Tai4基因。其中,A. tumefaciens物种中只有BIMB-1315G、1D1609、CFBP6623、12D1、CFBP6624、186、6N2、1D1460、EML4、CFBP7129、1D1108、6A、15955、FDAARGOS株系中发现各有1个Tae4基因,其他株系中未发现,且这18个株系基因组中均未发现存在Tai4基因。A. vitis物种中只有VAR06-30株系基因组中被发现1个Tae4基因和1个Tai4基因。并且发现,14个A. tumefaciensTae4基因都存在线形染色体(linear chromosome),而1个A. vitisTae4基因和1个A. vitisTai4基因存在A. vitisVAR06-30株系的环状染色体(circular chromosome)。
本文发现的Tai4基因编码蛋白都含有Tae4结构域(Pfam编号为PF14113),其中的A. tumefaciensTae4蛋白都含有EAGR_box结构域(Pfam编号为PF16713)。本文发现的唯一A. vitisTai4蛋白含有1个Tai4结构域(Pfam编号为PF16695)和1个Paramyxo_PCT结构域。此外,用于预测结构域的SMART软件还检测到A.vitisTai4蛋白含有一段长度为22个氨基酸的信号肽,序列为“MMYRLLLLGLVGLCVNSQIAAA”。
通过检测15个Tae4蛋白和1个Tai蛋白的性质发现,15个Tae4蛋白都各自包含166个氨基酸,其半胱氨酸均分别占各自氨基酸含量的1.8%;Tai4蛋白包含151个氨基酸,其半胱氨酸占自身氨基酸含量的2%。A. vitisTae4蛋白的等电点是8.96,分子量是18 446.84;A. vitisTai4蛋白的等电点是6.71,分子量是16 749.02。A. tumefaciens株系CFBP6623和CFBP6624的Tae4蛋白的等电点是8.81,其他12个都是9.05;A. tumefaciens的14个株系Tae4蛋白的分子量在18 214.44到18 353.57之间。15个Tae4蛋白与其他效应子一样不含有跨膜结构域,A. vitisTai4蛋白也不含有跨膜结构域。经预测,15个Tae4蛋白与A. vitisTai4蛋白都是GPI锚定蛋白,定位于线粒体。先前的研究显示,促进致病菌侵染植物的效应子都含有信号肽[6]。本研究结果显示,A. vitisTae4蛋白被预测含有信号肽的可能性略高于14个A. tumefaciensTae4蛋白。在线软件SignalP分析结果也显示,A. vitisTai4蛋白含有信号肽。
疏水性分析结果显示,A. vitisTae4蛋白疏水氨基酸和亲水氨基酸的分布与A. tumefaciensTae4蛋白略有差异。而本次鉴定出的1个Tai4蛋白和15个Tae4蛋白既不属于亲水蛋白质也不属于疏水蛋白质(图1)。

图1 蛋白的疏水性分析结果Figure 1 Hydrophobicity analysis of protein
2.2 蛋白序列比对与进化树分析
将14个A. tumefaciensTae4蛋白利用ClustalX 2.0软件进行比对发现,A. tumefaciensTae4蛋白比较保守,14个A. tumefaciensTae4蛋白之间的相似度较高。相对而言,A. tumefaciens株系CFBP6623和CFBP6624的Tae4蛋白相对于其他12个蛋白在4个氨基酸位点上有差异,与其他Tae4蛋白差别最大(图2 A)。A. tumefaciens株系FDAARGOS_1048的Tae4基因的起始密码子是TCA,终止密码子是CAT,而其他A. tumefaciensTae4蛋白的起始密码子都是ATG,终止密码子都是TGA。A. tumefaciens株系FDAARGOS_1048的Tae4蛋白序列并未因起始密码子和终止密码子的区别而有显著差异,其蛋白序列与大部分A. tumefaciensTae4蛋白相似度高,极为保守(图2 A)。而将14个A. tumefaciensTae4蛋白和A. vitisTae4蛋白利用ClustalX 2.0软件进行比对发现,A. tumefaciensTae4蛋白和A. vitisTae4蛋白相似度不高,而A. vitisTae4蛋白和A. vitisTai4蛋白的相似度也不高(图2 B)。

图2 蛋白比对结果Figure 2 Alignment of proteins
利用MEGA 6.0软件构建系统发育树显示,14个A. tumefaciensTae4蛋白可以被分为2组。一组包含A. tumefaciens株系CFBP6623和CFBP6624的Tae4蛋白,另一组包含其他12个蛋白。A. vitisTae4蛋白不属于这两个组,独立成为一个孤儿蛋白(图3)。利用在线软件Topo-phylogeny分析MEGA 6.0软件构建系统发育树的拓扑结构分析显示,该15个Tae4蛋白可以被分为3组:一组包含A. tumefaciens株系CFBP6623和CFBP6624的Tae4蛋白,另一组包含其他12个蛋白,第三组包含A. vitisTae4蛋白。进化树的分析结果与ClustalX 2.0软件比对结果高度一致。

图3 15个Tae4蛋白的进化树分析结果Figure 3 The evolutionary tree of 15 Tae4 proteins
2.3 葡萄根癌病病原菌中Tae4与Tai4蛋白结构及互作分析
利用在线软件MEME suite 5.3.3分析了15个Tae4蛋白和1个Tai4蛋白的保守基序(motif)。结果显示,14个A. tumefaciensTae4蛋白中可以鉴定出2个保守基序,其中motif1长达71个氨基酸,motif2包含19个氨基酸。15个根癌病病原菌Tae4蛋白间可以鉴定出10个保守基序,每个保守基序都含有10个氨基酸。这10个保守基序在A. tumefaciensTae4蛋白和A. vitisTae4蛋白中排序不同。
利用在线软件SWISS-MODEL预测了15个Tae4蛋白和1个Tai4蛋白的三级结构(图4)。由图4可见,A. tumefaciensTae4蛋白和A. vitisTae4蛋白的三级结构有所区别。A. tumefaciensTae4蛋白结构与SWISS-MODEL蛋白三级结构数据库中的模板6ijf.1.C具有93.87%的相似度。该模板的蛋白三级结构来源于A. tumefaciensATU4346和 ATU4347株系中的Tae4蛋白的晶体结构[8]。A. vitisTae4蛋白结构与SWISS-MODEL蛋白三级结构数据库中的模板蛋白4jur.1.A具有41.14%的相似度,且最接近这个模板蛋白。该模板蛋白三级结构来源于阴沟肠杆菌和鼠伤寒沙门氏菌的Tae4蛋白晶体结构[12]。此外,利用在线软件STRING分析两种葡萄根癌病病原菌Tae4和Tai4蛋白家族成员在各自物种中与其他蛋白的互作,结果显示,A. tumefaciensTae4可与10个蛋白互作,包括溶血素协同调节蛋白(Hemolysin-coregulated protein,HCP)、六型分泌系统蛋白VasG等(图5)。而除了A. vitisTae4蛋白和A. vitisTai4蛋白二者能互作外,没有预测到这两个蛋白在A. vitis物种中能与其他蛋白互作。分析基于STRING数据库中A. tumefaciens株系CCNWGS0286版本的数据。

图4 蛋白的三级结构预测结果Figure 4 The 3D structure of the protein
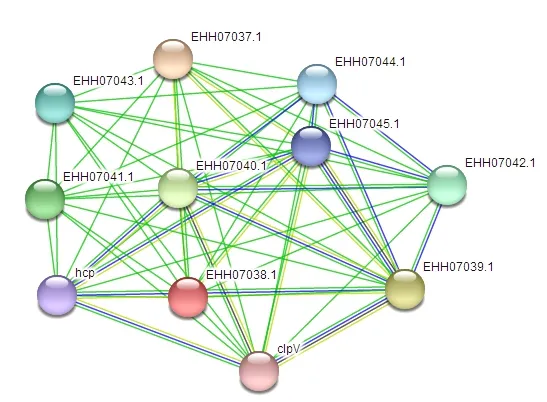
图5 STRING分析A. tumefaciens Tae4蛋白与物种中其他蛋白互作的结果Figure 5 STRING analysis of A. tumefaciens Tae4 proteins with other proteins in species
2.4 两种葡萄根癌病病原菌中Tae4蛋白编码基因的选择压力
将14个A. tumefaciensTae4蛋白的编码基因作为一个整体进行选择压力分析,PAML软件的M0模型分析结果显示,A. tumefaciensTae4蛋白编码基因整体遭受纯化选择(dN/dS=0.3719)。用PAML软件只在14个A. tumefaciensTae4蛋白的编码基因中发现了1个显著的正向选择位点(149F)。将14个A. tumefaciensTae4蛋白和1个A. vitisTae4蛋白的编码基因作为一个整体进行选择压力分析,PAML软件的M0模型分析结果显示,这15个Tae4蛋白的编码基因整体遭受正向选择(dN/dS=2.39292)。PAML软件M8模型用贝叶斯分析法在这15个Tae4蛋白的编码基因中发现了27个显著的正向选择位点。
3 讨论和结论
本研究分析了基因组被测序的18个A. tumefaciens和4个A. vitis株系的Tae4蛋白和Tai4蛋白及其编码基因。结果显示,18个A. tumefaciens株系基因组不是全部都含有Tae4基因,并且都不含有Tai4基因。然而,之前的研究显示,有的A. tumefaciens株系基因组含有Tai4基因,例如ATU4346和ATU4347株系[8]。本研究的4个A. vitis株系中只有VAR06-30株系的基因组含有Tae4基因和Tai4基因。这说明,对于A. tumefaciens和A. vitis,Tae4蛋白和Tai4蛋白虽然可以做为一种攻击其他病原菌的“武器”及自身免疫这种“武器”的防具[7],但这两种蛋白的编码基因并不是该两种葡萄根癌病病原菌生长发育所必须的基因。也说明A. tumefaciens和A. vitis不同株系基因组之间Tae4和Tai4两个基因位点的变异程度较高。有的株系基因组Tae4或Tai4基因缺失可能是由于基因组发生了结构变异(Structural Variation,SV)或插入缺失(Indel)。有的A. tumefaciens株系基因组缺失了Tai4基因,说明可能存在其他机制协助这些A. tumefaciens株系抵御自身Tae4蛋白的伤害。另一种可能是这些A. tumefaciens株系的Tae4蛋白具有较小的伤害作用。而这些A. tumefaciens株系是否能抵御其他属种的Tae4也需要进一步研究。在A. tumefaciens株系中,Tae4基因也存在碱基突变。例如A. tumefaciens株系CFBP6623和CFBP6624相对于其他株系在Tae4基因上存在较多的非同义突变,A. tumefaciens株系FDAARGOS_1048的Tae4基因的起始密码子和终止密码子相对于其他株系的Tae4基因存在碱基突变,但都属于同义突变,并没有引起氨基酸的改变。
对于A. tumefaciens和A. vitis的Tae4蛋白性质的分析显示,这些葡萄根癌病病原菌的Tae4蛋白都包含少于200个氨基酸,也不含跨膜结构域,这与之前报道的致病效应子类似[6]。A. tumefaciens和A. vitis的Tae4蛋白共有10个保守基序,然而这些保守基序在这两个物种中的排列顺序不同。这种保守基序排列顺序的差别可能导致了这两个物种中Tae4蛋白较显著的序列差异。尽管A. tumefaciens和A. vitis的Tae4蛋白都含有Tae4结构域,并且含有较多的保守基序,但是这些保守基序排列顺序的差异可能导致功能上的差异,本试验得到的蛋白互作分析结果也证明了这种差异可能存在。结果显示,A. tumefaciensTae4可与溶血素协同调节蛋白、六型分泌系统蛋白VasG等互作,而A. vitis的Tae4蛋白则不能。而选择压力分析显示,A. tumefaciens株系的Tae4基因间遭受纯化选择,蛋白之间没有新功能的产生。而A. tumefaciens和A. vitis的Tae4基因间遭受正向选择,且鉴定出许多正向选择位点,正向选择可能会导致功能分歧或新功能的产生[13]。因此,推测不同葡萄根癌病病原菌间的Tae4基因功能可能存在差异。A. tumefaciens和A. vitis的Tae4蛋白三级结构的差异也较明显。A. vitisTae4蛋白的三级结构与阴沟肠杆菌和鼠伤寒沙门氏菌的Tae4蛋白晶体结构更为相似。进化树的结果也证明了A. tumefaciens和A. vitis中Tae4蛋白的差异较大。
综上所述,该研究认为,不是全部A. tumefaciens株系基因组含有Tae4基因,部分A. tumefaciens株系不含Tai4基因。说明对于A. tumefaciens和A. vitis,Tae4蛋白和Tai4蛋白虽然可以做为一种攻击其他病原菌的“武器”及自身免疫这种“武器”的防具,但这两种蛋白的编码基因并非生长发育所必须的基因。A. tumefaciens和A. vitis的Tae4蛋白的氨基酸序列差异较大,不同葡萄根癌病病原菌间的Tae4基因功能可能存在差异。
