长柄扁桃种子油脂累积及脂肪酸转化规律
2021-12-17包文泉陈俊兴包洪胜王丽丽张淑宁白玉娥
包文泉,陈俊兴,敖 敦,包洪胜,王丽丽,张淑宁,张 裕,白玉娥
(1.内蒙古农业大学 a.林学院;b.草原与资源环境学院,内蒙古 呼和浩特 010011;2.岫岩满族自治县农业农村发展中心,辽宁 鞍山 114300)
近年来,随着食用植物油需求的日益增长以及对食用植物油品质要求的提高,木本油料已越来越受人们的关注[1]。木本油料树种根系发达,适应性强,可种植于荒山、荒滩和盐碱地等非耕地,并且一次种植可收获多年[2];此外,与传统的油料作物相比,木本油料油脂品质优,富含油酸、亚油酸和亚麻酸等不饱和脂肪酸,具有极高的食用、药用和保健作用[3]。据统计,我国木本粮油植物资源丰富,大多分布在“老、山、边、穷”地区,种子含油率高于40%,可食用油料树种就有150 多种;但目前开发利用的只有核桃、文冠果、山杏等10 余种,说明我国广袤山地丘陵中尚存在大量的“绿色油库”有待进一步发掘与开发利用[2-3]。可见,加强优质木本油料树种的开发与利用,对于保障我国粮油安全和生态安全具有重要的战略意义。
长柄扁桃Amygdalus pedunculata又称野樱桃,系蔷薇科扁桃属落叶灌木,是我国重要的生态经济型树种,其抗旱、抗寒能力极强,集中分布于内蒙古、河北和陕西等地,种质资源较为丰富,是我国干旱区水土保持、生态修复等生态建设的优选树种[4-5]。同时,长柄扁桃种子含油率高,出油率高达52%,油中不饱和脂肪酸含量高达95%,可作为高品质食用植物油,其开发利用潜力巨大[6]。目前。关于长柄扁桃的研究集中于遗传多样性、抗旱抗寒性和成分提取方面[5-7],而对长柄扁桃的油脂累积特点及脂肪酸转化规律的研究极少,因而无法采取科学准确的栽培管理措施,进而严重制约了长柄扁桃这一新型优质木本粮油的产业化发展进程。本研究通过分析长柄扁桃种子发育过程中油脂累积规律及脂肪酸组分转化特点,分析油脂累积过程中各脂肪酸间的相关性,研究结果不仅可为长柄扁桃果实的适时采收提供理论依据,还对提高长柄扁桃种子油脂产量和油脂品质等具有重要的理论指导和现实意义。
1 材料与方法
1.1 试验材料
以内蒙古良种繁育中心生长情况良好的长柄扁桃良种‘蒙扁1 号’为试材,自花后15 d 至种子成熟(106 d),每7 天采一次种仁,共采集14个发育期种仁,将其常温风干,备油脂含量和脂肪酸组分的测定;将14 期种仁样品分别记为花后15、22、29、36、43、50、57、64、71、78、85、92、99、106 d 的长柄扁桃种子。
1.2 油脂和脂肪酸组分的提取与分析
分别称取花后15、22、29、36、43、50、57、64、71、78、85、92、99、106 d 的10 g 干种仁研磨,采用索氏抽提法提取油脂,重复3 次。脂肪酸组分的测定采用气相色谱仪,重复3 次,参照陈俊兴等[8]的方法进行。
1.3 统计分析
应用Excel 2019 软件对发育期种子油脂含量和脂肪酸组分含量进行统计,并使用SPSS 20.0 软件对发育期种子脂肪酸组分进行方差分析和相关性分析。
2 结果与分析
2.1 长柄扁桃种子油脂累积规律
通过测定长柄扁桃幼果期至成熟期的种子含油量,初步掌握了长柄扁桃种子发育过程中油脂累积的动态规律,所得规律如图1所示。长柄扁桃种子油脂的动态累积变化模式呈“慢—快—慢”的单S 型,根据油脂含量的变化,可将其油脂累积过程分为3 个阶段,即为油脂缓慢累积期、油脂快速累积期和油脂减缓累积期。其中,花后15~50 d 长柄扁桃种子含油量从7.10%累积增值17.23%,日平均增量约0.3%,为油脂缓慢累积期;花后50~78 d 种子油脂含量快速提高,进入油脂快速累积期,种子油脂日平均增量达1.34%,并在花后78 d 累积至最高(54.67%),而花后78 至106 d,长柄扁桃种子油脂累积速度开始下降,逐渐进入油脂减缓累积期,于78 d 降至52.90%。

图1 长柄扁桃种子油脂累积的动态变化Fig.1 Dynamic changes in the accumulation of A.pedunculata seeds
2.2 长柄扁桃种子发育过程中脂肪酸组分转化规律
对长柄扁桃种子油脂主要成分进行检测分析可知(表1),长柄扁桃种子油脂中含有棕榈酸(C16∶0)、棕榈油酸(C16∶1)、硬脂酸(C18∶0)、油酸(C18∶1)、亚油酸(C18∶2)、亚麻酸(C18∶3)和二十碳烯酸(C20∶1)等7 种脂肪酸;其中硬脂酸和棕榈酸为饱和脂肪酸(SFA),油酸、二十碳烯酸和棕榈油酸为单不饱和脂肪酸(MUFA),而亚油酸和亚麻酸为多不饱和脂肪酸(PUFA)。通过对各发育期种子脂肪酸组分的分析,发现二十碳烯酸、亚油酸、油酸、硬脂酸和棕榈酸是长柄扁桃种子发育过程中的主要脂肪酸组分,在种子发育过程中,这五种组分含量变化均存在显著差异(P<0.01),其中油酸含量具有始终增量趋势,从29.94%增至75.61%;亚油酸含量则具有先增高后降低的规律,由28.26%增高至44.42%,随后再降低至18.20%;而二十碳烯酸、硬脂酸和棕榈酸的变化趋势相似,棕榈酸从11.19%减少至3.61%,硬脂酸含量从8.96%减少至1.24%,二十碳烯酸含量从20.69%逐渐减少至消失。长柄扁桃发育期种子中的棕榈油酸和亚麻酸的动态变化相似,在油脂缓慢累积期基本未检测到棕榈油酸和亚麻酸,而从油脂快速累积期后才出现微量棕榈油酸(<0.51%)和亚麻酸(<0.34%)[5,8]。

表1 长柄扁桃发育期种子脂肪酸组分及含量†Table 1 Fatty acid components and content of A.pedunculata seeds during development
2.3 饱和脂肪酸和不饱和脂肪酸含量变化规律
不同发育期长柄扁桃种子饱和脂肪酸与不饱和脂肪酸的动态变化分析结果显示(图2),在种子发育前期(15~78 d),饱和脂肪酸SFA、单不饱和脂肪酸MUFA 和多不饱和脂肪酸PUFA的含量都存在较大的变化,其中PUFA 的变化与MUFA 变化相反,如MUFA 含量先出现微量减少再出现迅速提高,由50.62%降低至45.76%,再增高至75.99%,动态变化差异显著;而PUFA含量先出现增高再出现降低,从28.26%增高至44.42%,再降低至18.12%,动态变化差异显著;SFA 含量则先出现降低再逐渐稳定,由20.15%降低至4.91%。此外,进入油脂减缓累积期后长柄扁桃种子中SFA、MUFA 和PUFA 的含量均趋于稳定,其含量无显著变化[8]。

图2 长柄扁桃发育期种子饱和脂肪酸、单不饱和脂肪酸和多不饱和脂肪酸的动态变化Fig.2 Dynamic changes of saturated fatty acids and unsaturated fatty acids in A.pedunculatus seeds
2.4 长柄扁桃种子发育过程中脂肪酸组分间的相关性分析
发育期种子脂肪酸组分间的相关性分析表明(表2),在种子发育过程中油酸与亚麻酸、油酸与棕榈油酸、亚麻酸与棕榈油酸间均存在显著正相关,其间的相关性系数分别为0.980、0.772 和0.751;硬脂酸与棕榈酸、二十碳烯酸与棕榈酸、二十碳烯酸与硬脂酸间同样存在显著正相关,其相关性系数分别为0.977、0.951 和0.933;而油酸和亚麻酸分别与二十碳烯酸、棕榈酸、亚油酸、硬脂酸存在显著负相关,并且相关性系数均大于0.800[8]。
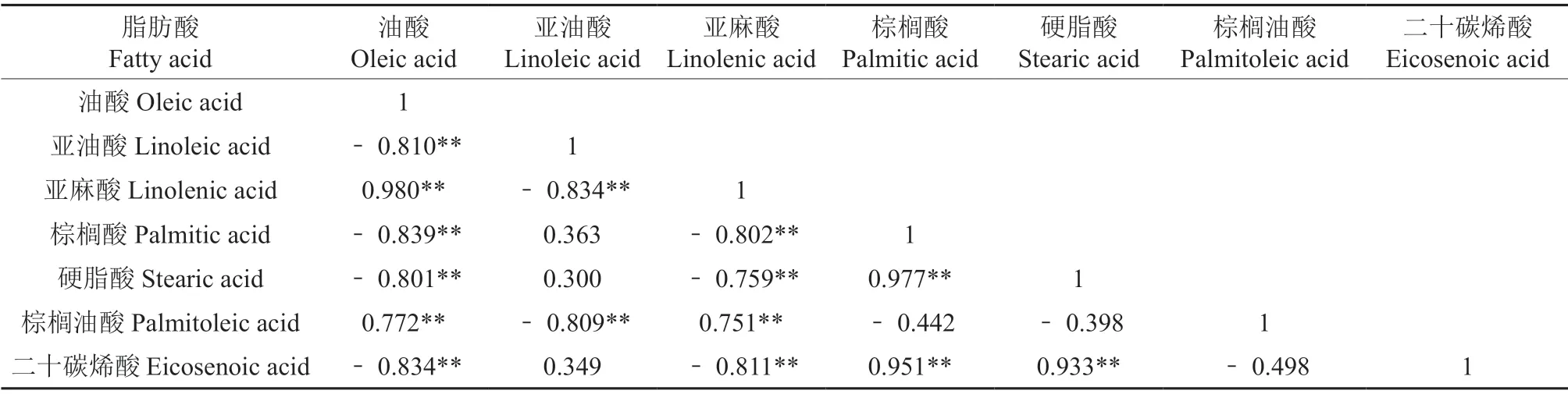
表2 长柄扁桃发育期种子中脂肪酸组分间的相关性Table 2 Correlation between fatty acid components in seeds with A.pedunculatus during development
3 结论与讨论
含油率是衡量木本油料树种开发利用价值的重要性状。随着种子发育,木本油料树种的含油率也发生动态变化,不同树种或同一树种种子发育期不同,其种子含油量存在显著差异,但木本油料树种种子油脂累积过程通常可分为3 个阶段,大多呈“S”型累积模式[9-10]。对长柄扁桃发育期种子油脂累积的动态变化研究表明,长柄扁桃种子油脂累积变化模式呈典型的单S 型,可分为油脂缓慢累积期、油脂快速累积期和油脂减缓累积期。这与木本油料树种野山杏[11]、文冠果[12]、麻疯树[13]、黄连木[14]、白檀[15]等的种子油脂累积规律的研究结果相似,符合绝大多数木本油料果实或种子的油脂累积规律。长柄扁桃种子发育初期(15~50 d)油脂累积缓慢,累积增量小于0.3%,这与种子发育初期林木个体有限的营养物质和较弱的同化能力有关外,还与甘油三酯合成初期,关键调控酶与蛋白正处于启动阶段有关[11]。由于自身同化能力和相关酶活性的提高,自花后50 d开始长柄扁桃种子油脂累积速度提高,平均日增油量高达1.34%,并于花后78d 油脂含量达最大值。因此,在该阶段应加强地面管理和树体管理,以期提高长柄扁桃种子油脂产量和质量,并初步确定采收时间。此外,从花后78d 开始,长柄扁桃种子含油量变得逐渐稳定,这与山杏、文冠果等其他木本油料树种相似[11-12],这可能是种子发育后期含水量显著降低导致油脂合成和转化相关酶活性的降低,进而抑制其脂肪酸的合成[16]。
三酰甘油是植物种子油脂的主要成分[17-18]。木本油料树种种子三酰甘油的脂肪酸以16~18个碳脂肪酸为主,如油酸、亚油酸、棕榈酸和亚麻酸[18]。长柄扁桃发育期种子油脂中共检测出7种脂肪酸,链长分别为16、18 和20 个碳,这与许新桥等[1]研究毛乌素沙地长柄扁桃种子脂肪酸组分变异的研究结果一致,但其各脂肪酸组分含量存在差异,这可能是不同材料间遗传背景差异所致[11]。同一树种间脂肪酸组分含量存在较大差异,这一结果同样也存在于麻疯树[20]和文冠果[21]等木本油料中。
木本油料树种种子油脂累积特点、脂肪酸组分转化规律及其相关性研究可为油脂提质增产提供科学依据。长柄扁桃种子发育过程中,C18脂肪酸含量最高,占总脂肪酸的90%以上,其中,油酸构成其主要单不饱和脂肪酸,而亚油酸构成其主要多不饱和脂肪酸,并且二者的动态累计模式变化表现完全相反,符合油酸是亚油酸生物合成的前体物质这一结论[11]。棕榈酸、硬脂酸和二十碳烯酸是长柄扁桃种子的主要饱和脂肪酸(SFA),其含量分别由11.19% 降至3.61%、8.96% 降至1.24%和20.69%降至0。食用植物油中,SFA、MUFA 和PUFA 的比例是决定其营养价值和稳定性的重要指标[22]。通常认为高品质油应含有较高比例的MUFA、较低比例的PUFA 和SFA,长柄扁桃种子油脂中SFA∶MUFA∶PUFA 比例为4.85%∶76.11%∶18.53%,属于营养价值高且稳定强的高品质油脂。长柄扁桃种子SFA、MUFA 和PUFA 的含量均在油脂含量最大期(78 d)达稳定,可见,根据油脂累积和脂肪酸转化特点制定采收期,可获取高品质种子油。同时,本研究发现长柄扁桃种子油脂累积过程中油酸、亚麻酸、棕榈油酸三者间呈显著正相关,棕榈酸、硬脂酸和二十碳烯酸三者间同样呈显著正相关,而油酸和亚麻酸分别与二十碳烯酸、亚油酸、硬脂酸、棕榈酸呈显著负相关;表明后期如对某一脂肪酸组分进行选择时,可根据脂肪酸组分间的相关性,定向培育或改良高品质油用长柄扁桃种质。
综上所述,长柄扁桃种子油脂累积及脂肪酸组分转化是个复杂的生理生化过程,了解其油脂累积及脂肪酸转化特点,采取相应的管理措施,对获得高产、高品质的长柄扁桃种子至关重要。但油脂累积及脂肪酸转化还与生境、气候等环境因子有关,并且某一种质油脂累积规律并不能完全代表长柄扁桃种子油脂累积及脂肪酸转化规律[23-24]。因此,后期应进一步开展长柄扁桃不同种源和种质的油脂累积及脂肪酸转化相关的研究,找出其相似点与差异,结合栽培管理,为油用长柄扁桃育种的配套措施提供参考。
