亚热带森林附生植物叶片气孔特征及其可塑性对光照变化的响应
2021-10-16普晓妍王鹏程李苏鲁志云宋钰
普晓妍 王鹏程 李苏 鲁志云 宋钰
摘 要:附生植物是熱带亚热带森林生态系统中物种多样性极高且极其脆弱敏感的生物类群之一。光照被认为是促进附生植物由陆生类群演化而来并决定其生长和分布的关键因素。然而,由于接近林冠和规范性采样的限制,附生植物与光照的关系仍亟待阐述。为揭示附生植物对光强变化的响应和适应策略,该研究以亚热带常绿阔叶林6种附生植物(林冠层木本:鼠李叶花楸、毛棉杜鹃;林冠层草本:狭瓣贝母兰、毛唇独蒜兰;树干区草本:点花黄精、距药姜)为对象,对其在4个光处理梯度下生长的叶片气孔特征及其可塑性进行了对比分析。结果表明:(1)2种附生小乔木的气孔面积(SA)、气孔密度(SD)、潜在气孔导度指数(PCI)和表皮细胞密度(ECD)均对光强改变显著响应。2种附生兰科植物的SA最大,而SD最小;附生乔木叶片SD和ECD的光响应趋势与陆生植物更相似,而附生草本则出现种间差异。(2)6种附生植物的气孔、表皮细胞特性及其表型可塑性,在草本-木本、常绿-落叶植物、林冠-树干区之间,均无明显差别。(3)附生植物气孔特性和表皮细胞平均可塑性指数均低于陆生植物。综上结果表明,亚热带常绿阔叶林中附生植物对于光环境变化的适应性相对较弱。不同的附生植物可以通过不同程度地增加叶片SD和ECD来适应高光强生境,并通过对SD和SA的双重调节以增大潜在光合能力从而应对低光胁迫。
关键词:植物生理生态,哀牢山,林冠,光环境改变,附生植物,气孔特性,表型可塑性
中图分类号:Q948
文献标识码:A
文章编号:1000-3142(2021)09-1465-11
Abstract:Epiphytes,which grow non-parasitically upon other plants,comprise one of the most diverse and vulnerable groups in subtropical and tropical forests. Light can promote the evolutionary transition of plants from terrestrial to epiphytic life-forms and determine the growth and distribution of epiphytes. Nevertheless,the research on the relation between epiphyte growth and light stress has been plagued by some technical problems such as direct canopy access and systematic sampling. In order to explore the adaptation strategies and mechanisms of epiphytes to light change,six vascular epiphytes,including two canopy trees Sorbus rhamnoides and Rhododendron moulmainense,two canopy herbs Coelogyne punctulata and Pleione hookeriana,two trunk herbs Polygonatum punctatum and Cautleya gracilis,were surveyed to study the stomatal characteristics and plasticity in leaves of vascular epiphytes and their responses to light change in evergreen broad-leaved forests in the Ailao Mountains of Yunnan Province. The results were as follows:(1)The stomatal area (SA),stomatal density (SD),potential stomatal conductance index (PCI)and epidermal cell density (ECD)of the two canopy epiphytic trees responded significantly to light change. The canopy epiphytic herbs had the largest SA and the smallest SD among all the studied epiphytes. The response of the SD and ECD of epiphytic trees to light change was consistent with those of terrestrial plants,while epiphytic herbs showed interspecific variations. (2)The stomatal and epidermal cell characteristics,and phenotype plasticity of epiphytes showed no significant differences between herbs and trees,evergreen and deciduous plants,and canopy and trunk epiphytes. (3)Their plasticity indexes of stomatal and epidermal cell characteristics were lower than those of terrestrial plants. Our results indicate that epiphytes have relatively weak adaptability to light change in subtropical forests. Epiphytes are able to adapt to high light intensity by increasing SD and ECD with different degrees,and to cope with low light stress by increasing potential photosynthetic capacity with trade-offs between SD and SA.
Key words:plant physioecology,Ailao Mountains,canopy,light change,epiphyte,stomatal characteristic,phenotype plasticity
附生植物是一类生长于其他植物体表,但并不掠夺宿主营养和水分的生物类群,其水分和养分主要从空气中获取,对环境变化极为敏感,可作为监测气候变化的重要指示生物(bio-indicator)(宋亮和刘文耀,2011;Leao et al.,2014)。其中,维管附生植物约占全球已知维管植物物种的9%(Zotz,2013),并在森林水分平衡和营养循环(Nadkarni,1984;Coxson & Nadkarni,1995),以及为其他生物提供更多样资源和生态位(Nadkarni & Matelson,1989;Montero et al.,2009;Mccracken & Forstner,2014)等方面发挥着重要作用。我国拥有世界上面积最大的亚热带常绿阔叶林,并在西南及华南地区分布着一定数量的热带森林,附生植物非常丰富(徐海清和刘文耀,2005)。然而,随着人类干扰和社会经济活动的不断扩大,亚热带常绿阔叶林植被普遍遭到严重破坏。分布于我国西南地区的热带森林和高海拔山地森林,森林片断化也日益严重,其中对环境变化异常敏感的附生植物类群受到显著的影响。但令人担忧的是,对于这一和环境变化高度相关的生物类群,目前在我国所给予的关注却远远不够。森林附生植物对环境变化的响应以及相关的生理生态学机制等方面的研究都亟待加强。
光照是植物光合作用能量的唯一且最终来源,也是影响光合作用的最根本因素(叶子飘和于强,2008)。附生植物自身对光环境改变的形态和生理适应性,是影响其分布和生长的关键因子。但是关于附生植物的生理生态学特征对光环境改变的响应机制仍未得到充分阐明(Hietz & Briones,1998;Zotz & Andrade,1998;Stuntz & Zotz,2001)。并且,附生植物自身和生境的特殊性,也常导致附生植物对光照变化的响应明显不同于陆生植物。如研究发现,凤梨科、兰科(匡美龄和张石宝,2015)、蕨类和榕属植物附生(Hao et al.,2011)和陆生类群的光响应特征普遍明显不同(张强等,2008;宋娟等,2013;Scarano et al.,2016)。因此,附生植物应对各种光环境的生理生态适应性策略和机制,亟待专门探讨和分析。
尤其值得注意的是,气孔(stomata)是植物与外界环境进行水气交换的主要途径,是植物在不断的进化中保留下的器官,具有较强的稳定性(Caglar & Tekin,1999;朱燕华,2013)。植物的光合作用和蒸腾作用是生态系统物质循环的两个最基本生理过程,均为气孔行为所控制(Hetherington & Woodward,2003;Tayloy et al.,2012)。由于植物气孔行为对环境变化反应敏感,通过对测试气孔性状参数来探究植物对环境变化的调节过程已成为近些年相关研究的热点(Uprety et al.,2002;Wang et al.,2014;Liu et al.,2018)。因此,本研究选择云南哀牢山亚热带常绿阔叶林的6种常见附生维管植物为研究对象,通过光控组下附生植物叶片的气孔和表皮细胞特征的比较研究和表型可塑性分析,探讨光环境变化对不同生活型附生植物气孔和表皮特性的影响程度的差异,旨在揭示附生植物叶片气孔和表皮特性对光强变化的适应策略和机制,为理解附生植物对生境的光变化适应提供依据,为亚热带常绿阔叶林的附生植物类群在生境遭到破坏后的生态恢复提供理论参考。
1 材料与方法
1.1 试验样地
哀牢山位于云南高原西南部、横断山脉南段,属云岭向南分支余脉,至今保存着我国最完整最大的中山濕性常绿阔叶林(吴征镒,1983)。其中,景东段徐家坝地区位于哀牢山西坡(100°44′—101°30′ E、23°36′—24°56′ N),年平均气温11 ℃,具有温暖性-温凉性气候特征。在海拔2 200~2 600 m的地区有大面积连片分布的中山湿性常绿阔叶林(也称原生木果柯林,primary Lithocarpus xylocarpus forest),约占该地区植被面积的85%以上;海拔2 600 m以上为岛状分布的山顶苔藓矮林。木果柯林群落高25~30 m,群落分层现象十分明显,以木果柯(Lithocarpus xylocarpus)、硬壳柯(L. hancei)和变色锥(Castanopsis wattii)等为群落上层的优势树种(李贵才等,2001)。
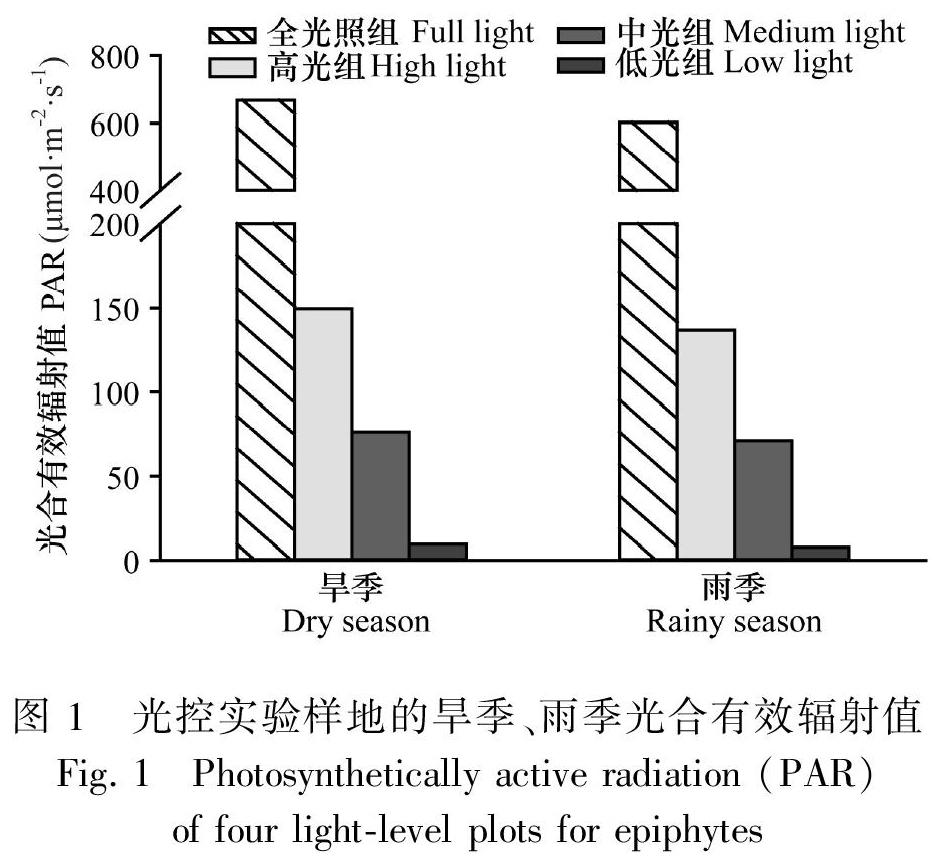
本研究选择中国科学院哀牢山生态站附近旷地,根据附生植物所分布自然生境的光照水平,在保证降水、空气湿度、温度、海拔等条件基本一致的情况下,以全光照组(不架设遮荫网)为对照,利用钢架和不同规格的遮荫网设置高光照、中光照和低光照组,开展相关试验。光处理组的划分标准参照本地区已有森林垂直光照和2015年雪灾前后森林群落林冠盖度数据(李麟辉等,2011;Song et al.,2018)。高光照组基本对应雪灾后木果柯林以及其相应栎类次生林(仅有少量附生维管植物分布)的光照水平;中光照组对应木果柯林雪灾前的冠层下平均光照;低光照对应部分原生林林冠完全郁闭时的光照。在搭建好的4个样地,分别悬挂U23-001温湿记录仪(Hobo,USA)和自计式光量子计(Odyseey,Newsland),记录大气温度、大气相对湿度和光合有效辐射(PAR)等参数。而植物生长基质含水量通过调节浇水量和频度,保持水分条件基本充足。
2016至2019年监测数据显示,4个光控样地的年均PAR为636.84、143.71、73.99、8.84 mol·m-2·s-1(图1);以全光照组作为对照,PAR的全年、雨季、旱季的比例均为全光照组∶高光组∶中光组∶低光组=100∶22∶11∶1,而样地间的年均温度(13.21~15.27 ℃)和相对大气湿度(58.77%~64.18%)差异在±0.5%以内,可认为光照是光控样地唯一的调控因子。
1.2 试验物种的选择和栽培
本研究选择云南哀牢山湿性常绿阔叶林分布较广的6种附生维管植物为研究对象,具体见表1。所有附生植物植株均采自原生常绿阔叶林内2015年雪灾导致的倒木。采集以完整成熟个体或多年生根茎为主,而毛棉杜鹃和鼠李叶花楸因成熟个体较大,不易成活,收集高度低于2 m的个体。将采集到的植株移植于培育(中等光照)遮荫棚内的花盆,花盆内苔藓/腐殖土的体积比例约为1∶1。在移植培育期间,保证所有植株土壤水分条件基本一致,及时除去非目标植物,但均不进行人工施肥。在目标植物健康生长一段时间后,于2018年1月初移植到上述4个光处理组样地生长,每个光处理组样地内每个物种不少于20盆。
1.3 测定方法
于2019年7月,每个光处理组每个物种选择6盆样本,分别采集1片当年新叶的成熟叶片,制成标本带回实验室。切取边长0.5 cm左右的片段,先用溶解有适量氯化钾的硝酸溶液浸泡样品12 h后取出,再用5%氢氧化钾溶液浸泡处理至半透明,然后用镊子和解剖针撕开叶片表皮,用毛笔刷去叶肉,表皮用1%番红染色剂染色,最后制成玻片后利用徕卡显微镜(Leica DM1000LED)拍照。用ImageJ图片分析软件测量×40倍视野下气孔长度和宽度;统计×20倍视野下气孔和表皮细胞数量,每个叶片分析6个视野。相关计算公式如下。
气孔指数(stomatal index,SI):SI=SN/(SN+ECN)。式中:SN(stomatal No.)是气孔个数;ECN(epidermal cell No.)是表皮细胞个数。
气孔面积(stomatal area,SA):SA=1/4×π×SL×SW。式中:SL(stomatal length)是气孔长度;SW(stomatal width)是气孔宽度。
气孔密度(stomatal density,SD):取6个视野中的平均值。
潜在气孔导度指数(potential conductance index,PCI):PCI=SL2×SD×10-4。 式中:SL是气孔长度;SD是气孔密度。
表皮细胞密度(epidermal cell density,ECD):取6个视野中的平均值。
气孔特性及表皮细胞密度可塑性指数按照Valladares et al.(2000)方法计算:可塑性指数=(最大值-最小值)/最大值。
1.4 统计分析
使用Sigmaplot 14软件,对不同光照组间附生植物的叶片气孔特性和表皮细胞特性参数进行单因素方差分析;同时使用SPSS 25软件,利用回归分析探讨叶片气孔特性和表皮细胞特性随PAR变化的趋势。
2 結果与分析
2.1 叶片气孔和表皮细胞特性参数在不同光强处理下的差异显著性及回归分析
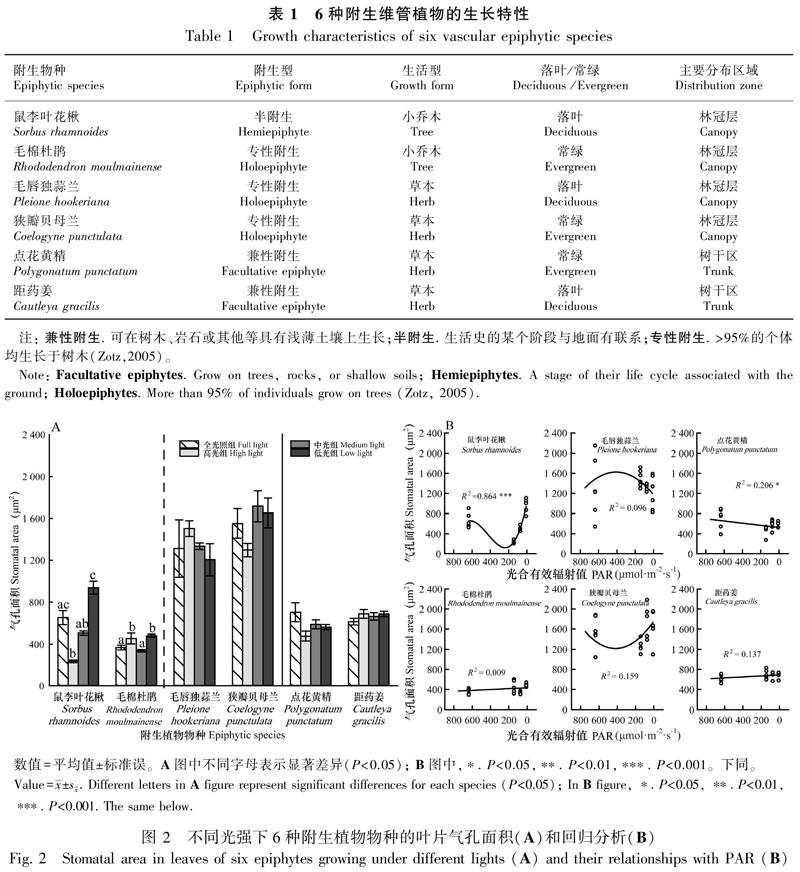
2.1.1 气孔面积(SA)在不同光强处理下的差异显著性及回归分析 结果显示,仅2种林冠层附生乔木叶片SA在不同光强下有显著差异(图2:A);相关分析进一步显示,仅2种附生植物的叶片SA与PAR显著相关(图2:B)。其中,鼠李叶花楸的叶片SA与PAR呈显著三次曲线(Cubic)相关(R2=0.864,P<0.001),高光组显著低于全光照组(P=0.015)和低光组(P<0.001);毛棉杜鹃叶片SA与PAR呈微弱负相关,但不显著,且在全光照组和中光组显著低于高光组(P=0.034)和低光组(P=0.001);毛唇独蒜兰和狭瓣贝母兰的SA随PAR降低均呈二项式分布,分别在高光组出现最大值和最小值;点花黄精的叶片SA与PAR呈显著正相关(R2=0.206,P<0.05);而距药姜SA虽随PAR下降稍微上升,但变化并不显著。2种附生兰科植物毛唇独蒜兰(P≤0.004)和狭瓣贝母兰(P<0.001)SA显著大于其他4种附生植物。
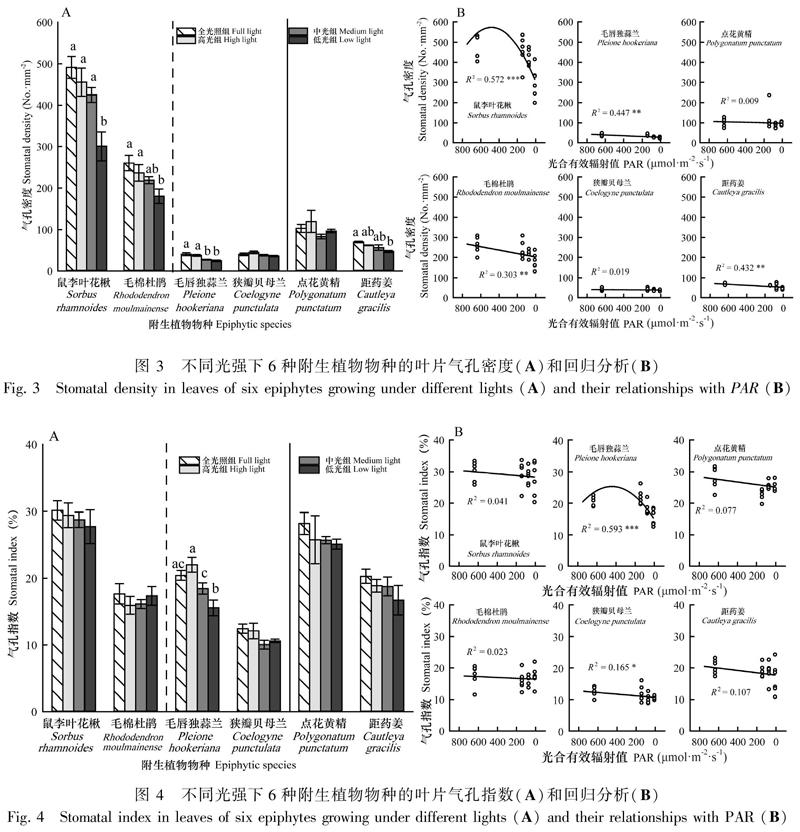
2.1.2 气孔密度(SD)在不同光强处理下的差异显著性及回归分析 结果显示,4种附生植物叶片SD在不同光强下有显著差异(图3:A);且叶片SD均与PAR显著相关(图3:B)。鼠李叶花楸、毛棉杜鹃、毛唇独蒜兰和距药姜的叶片SD在低光组均有显著降低趋势(P≤0.008),且变化趋势与PAR显著相关(P≤0.045)。鼠李叶花楸的变化趋势明显呈二项式分布。
2.1.3 气孔指数(SI)在不同光强处理下的差异显著性及回归分析 结果显示,仅毛唇独蒜兰的叶片SI在低光组显著低于其余3个光控组(P≤0.029)(图4:A),与PAR呈显著二项式相关(R2=0.593,P<0.001)(图4:B);而狭瓣贝母兰叶片SI与PAR呈显著线性相关(R2=0.165,P<0.05)(图4:B)。
2.1.4 潜在气孔导度指数(PCI)在不同光强处理下的差异显著性及回归分析 结果显示,4种附生植物叶片PCI在不同光强下有显著差异(图5:A),相关分析进一步显示,2种附生植物PCI与PAR显著相关(图5:B)。其中,鼠李叶花楸叶片PCI于PAR呈显著三次曲线(Cubic)相关(R2=0.814,P<0.001),高光组显著低于全光照组(P=0.028)和低光组(P<0.001);狭瓣贝母兰叶片PCI与PAR呈显著二项式相关(R2=0.375,P<0.001),高光组显著低于中光组(P=0.021)和低光组(P=0.003);毛棉杜鹃叶片PCI与PAR呈微弱负相关但不显著,中光组显著低于低光组(P=0.006);毛唇独蒜兰、点花黄精和距药姜叶片PCI与PAR无显著相关,但点花黄精叶片PCI在高光组显著低于全光照组(P=0.039)和中光组(P=0.006)。2种附生兰科植物毛唇独蒜兰(P<0.001)和狭瓣贝母兰(P≤0.026)PCI显著大于其他4种附生植物。
2.1.5 表皮细胞密度(ECD)在不同光强处理下的差异显著性及回归分析 结果显示,4种附生植物ECD在不同光强下有显著差异(图6:A),相关分析进一步显示,3种附生植物ECD与PAR显著相关(图6:B)。鼠李叶花楸(R2=0.627,P<0.001)和毛棉杜鹃(R2=0.589,P<0.001)叶片ECD与PAR呈显著二项式相关,且在低光组显著低于另外3个光控组(P≤0.011);毛唇独蒜兰叶片ECD与PAR呈显著线性相关(R2=0.472,P<0.01),在中光组显著低于全光照组(P=0.001);点花黄精和狭瓣贝母兰叶片ECD与PAR呈微弱负相关,但不显著,点花黄精在中光组显著低于高光组和低光组;距药姜的叶片ECD与PAR呈微弱正相关,在4个光控组间无显著差异。
2.2 不同光强下气孔特性及表皮细胞的可塑性
附生植物不同的气孔参数,其可塑性有一定差异(图7)。6种附生植物的气孔特性和表皮细胞平均可塑性大小顺序为潜在气孔导度指数>气孔面积>气孔密度>气孔指数>表皮细胞密度。附生草本与木本、落叶植物与常绿植物以及林冠层和树干区附生植物之间,相关参数的表型可塑性均无明显差别。
3 讨论与结论
气孔是植物表皮的一个重要器官,一般由成对的保卫细胞及保卫细胞之间的孔隙组成,其孔径大小直接决定着植物的蒸腾作用和光合作用(郑玉龙等,2005)。外部环境因素(光照强度、水分条件、营养状况等)会影响气孔形状及生理特征,从而对植物蒸腾、光合和呼吸等生理活动产生影响(孙广玉和邹埼,1991;吕洪飞,2000;曹冬梅等,2002;Hetherington & Woodward,2003)。然而,目前国内外有关附生植物光响应机制的研究多集中在叶片形态、叶绿素含量、光合作用及呼吸作用等方面(Schfer & Lüttge,1988;Maxwell et al.,1994;Pires et al.,2012;Liu et al.,2018),而有关光强对附生植物叶片解剖结构如气孔和表皮特征的影响机制了解尚少。基于此,本研究通过光控试验探讨了不同光强下附生植物叶片气孔和表皮细胞特性的响应特征。
3.1 附生植物叶片气孔面积和气孔密度对光环境变化的响应
本研究结果显示,附生乔木和草本植物之间的叶片SD和SA存在较大差异,附生乔木SD大于附生草本,而SA小于后者,这与Liu et al.(2018)对热带和寒温带森林737种陆生植物叶片气孔特征的研究中发现陆生植物叶片SD呈现出乔木>灌木>草本的趋势,而气孔大小则表现为草本最大的研究结果相符合。同时,相关研究还表明植物的最大气孔导度由SD和气孔大小共同决定,二者之间存在权衡(Franks et al.,2009)。一般来说,乔木树种为获得更多水分,倾向于增加SD来提高最大光合效率以满足自身需求;而草本植物生命周期较短,更傾向于通过增大SA来快速提高光合作用效率,完成生命周期内的生长需要(Franks & Beerling,2009;Lammertsma et al.,2011;成雪峰,2013)。这很好地解释了本研究中林冠层附生兰科植物和小乔木之间叶片SA的差异。
附生植物SD对光照变化的响应虽然存在种间差异,但是部分结果与陆生植物较为一致。如林冠层附生乔木(鼠李叶花楸、毛棉杜鹃)及毛唇独蒜兰和树干区点花黄精的叶片SD对光照变化的响应趋势与陆生植物近似,均随光强减弱显著降低(何若天和吕成群,1995;孟雷等,2002;蔡志全等,2004);而林冠区的狭瓣贝母兰和树干区的距药姜叶片SD在不同光强下并无显著差异,明显不同于多数陆生植物(孟雷等,2002;贺静等,2007),表明其SD特征对光照变化并不敏感。
附生植物SA对光照变化的响应同样存在种间差异。如林冠层附生乔木叶片SA随光强减弱显著增大的趋势,与蔡志全等(2004)的研究相符。而在全光照组和高光组其SA均出现显著增大,这可能是附生植物应对高光胁迫的一种适应策略,即通过同时增大气孔密度和气孔面积以增大气孔导度来达成高光时的高光合速率。4种附生草本叶片SD在不同光强下并无显著差异,与多数陆生植物并不一致(孟雷等,2002;贺静等,2007),表明附生草本的叶片SA对光照变化并不敏感。
3.2 附生植物叶片气孔指数对光环境变化的响应
本研究显示,附生植物叶片气孔指数(SI)受到光照变化的影响,随着光强减弱均呈降低趋势,且2种附生兰科叶片SI与PAR显著相关,这与早期研究认为SI主要受水分的影响,而不受光照的影响结论不相符(Ticha,1982);SI常作为气孔分化强度的指标,是表示单位叶表面上气孔频度的常用参数(Artemios & George,2002;Masle et al.,2005)。蔡志全等(2004)曾发现热带雨林3个中层种和4个冠层种树苗的叶片气孔指数随光强增大而增大。本研究除毛唇独蒜兰以外的5种附生植物均与蔡志全等(2004)研究结果近似。这或许表明降水相对充足的热带亚热带森林中某些植物类群可能普遍存在着类似的适应机制。
3.3 附生植物叶片潜在气孔导度对光环境变化的响应
潜在气孔导度指数(PCI)是反映植物潜在光合能力的重要指标(Holland et al.,2009),由气孔密度和气孔长度共同决定该指数的值,也可近似理解为单位面积的气孔面积(Tian et al.,2016)。本研究显示,附生植物PCI对光照变化的响应存在种间差异,且与陆生植物相关研究多不一致。如6种附生植物中,除点花黄精外,PCI均呈随光强减弱而增大的趋势,这与早期张大鹏(1987)对不同光强下水稻叶片SD和气孔长度在弱光环境下降低的研究结果不一致,其原因可能是附生植物的原生生境由于林冠的遮挡,光强较之水稻生长环境更弱(徐海清和刘文耀,2005),附生植物对弱光的适应和利用策略优于前者。同时,2种附生兰科PCI显著大于另外4种附生植物,这与温婧雯等(2018)发现太白山乔木、灌木和草本植物之间PCI无显著性差异的现象同样不同。这或许暗示了附生植物可能通过SD和气孔大小的双重调节来增大潜在光合能力并降低气孔导度以应对低光胁迫,弱光利用率优于部分陆生植物。
3.4 附生植物叶片表皮细胞密度对光环境变化的响应
本研究显示,附生植物叶片表皮细胞密度(ECD)与SD的光响应趋势基本一致,对光照变化的响应虽然存在种间差异,但是部分结果与陆生植物研究结果不相符。如2种林冠层附生乔木叶片ECD在全光照组表现出光抑制现象,毛唇独蒜兰的ECD随光强增加而显著增加,与Lee et al.(2015)对红叶莴苣在不同光处理下叶片ECD无显著响应的研究结果均不一致。可以看出附生乔木的叶片SD和ECD的光响应趋势与陆生植物更相似(何若天和吕成群,1995;蔡志全等,2004),而附生草本则有明显的种间差异。这表明不同生活型的附生植物通过不同程度降低叶片SD和ECD以适应低光胁迫。
3.5 附生植物叶片气孔和表皮细胞特性参数应对光环境改变的表型可塑性
表型可塑性是生物适应环境变化的重要方式。高的可塑性常常暗示植物对环境具有较高的潜在适应能力,具有更宽的生态幅和更好的环境耐受性(Strauss-Debenedetti & Bazzaz,1991;耿宇鹏等,2004)。本研究显示6种附生植物的气孔特性和表皮细胞可塑性与陆生植物并不相同。附生植物气孔特性和表皮细胞平均可塑性指数均低于前人对于陆生乔木和草本的研究(蔡志全等,2004),且附生草本与附生木本以及林冠层和树干区附生植物之间的表型可塑性并无明显差别。这与陆生植物通常在垂直分布区域内处于上层的树种比处于下层的树种在生理和形态上具有更大的表型可塑性的现象并不相符(Valladares et al.,2000;温婧雯等,2018)。这可能说明附生植物表现出对于环境变化的较弱适应性,生态幅相较于陆生植物更窄,对环境变化的耐受性更差。这也从表型可塑性的角度解释了附生种子植物通常只在原生林分布且对环境变化极端敏感的事实。
综上所述,亚热带森林系统中,附生植物可以通过不同程度地增加叶片气孔密度和表皮细胞密度来适应生境光强的增加,并通过对气孔密度和气孔面积的双重调节来增大潜在光合能力以应对低光胁迫;但不同生活型的附生植物的面对低光胁迫或高光胁迫的机制有所不同。如本研究中,林冠层的附生乔木和树干区附生草本的气孔特征的光响应规律和陆生植物基本一致(何若天和吕成群,1995;孟雷等,2002;蔡志全等,2004;Liu et al.,2018)。2种附生兰科植物气孔特征光响应机制的不同(孟雷等,2002;贺静等,2007),可能与其是否属于景天酸代谢植物有关,这仍有待进一步的研究进行解释和说明。分布于不同垂直区域内的附生植物间以及上层树种与下层树种间的表型可塑性,均无明显不同(Valladares et al.,2000;蔡志全等,2004;温婧雯等,2018),且附生植物的气孔特性和表皮细胞平均可塑性指数均低于陆生乔木和草本(蔡志全等,2004;Liu et al.,2018),这可能说明附生植物对于环境变化的适应性相对较弱,对光照变化的耐受性更差。开展研究所在的哀牢山森林系统自2015年雪灾后的自我恢复过程中(Song et al.,2017;Song et al.,2018),应该对附生植物群落的变化进行长期观测和关注,以期更好地保护相关敏感生物类群的物种多样性。
不足的是,本文仅从附生植物叶片气孔特性的变化方面探讨了光照强度改变对附生植物物种分布和个体生长的影响。尤其值得注意的是,针对个别附生植物如鼠李叶花楸的气孔面积和潜在气孔导度与PAR进行拟合时,二项式模型均出现最低值为负的现象,明显不符合植物生理生态学常理;而改用Cubic模型进行拟合时,R2改变极小。这或许暗示了附生植物对高光至低光改变过程中的适应过程和机制,可能更加复杂和多变。未来仍需开展更广泛的研究,综合深入探讨森林系统附生植物的光响应机制。
致谢 感谢中国科学院哀牢山亚热带森林生态系统研究站对本研究野外试验的支持,感谢中国科学院西双版纳热带植物园宋亮老师提供切片实验器材支持。
参考文献:
ARTEMIOS MB,GEORGE K,2002. Comparative effects of drought stress on leaf anatomy of two Olive cultivars[J]. Plant Sci,163(2):375-379.
CAGLAR S,TEKIN H,1999. The stomata density of Pistachio cultivars on different pistacia rootstocks[J]. Turk J Agric For,23(EK5):1029-1032.
CAI ZQ,QI X,CAO KF,et al.,2004. Response of stomatal characteristics and its plasticity to different light intensities in leaves of seven tropical woody seedlings[J]. Chin J Appl Ecol,15(2):201-204.[蔡志全,齊欣,曹坤芳,等,2004. 七种热带雨林树苗叶片气孔特征及其可塑性对不同光照强度的响应[J]. 应用生态学报,15(2):201-204.]
CAO DM,KANG LF,WANG YS,et al.,2002. Effect of K spray on stomatal characteristics and photosynthetic rate of young apple trees under external application of root[J]. J Shanxi Agric Sci,30(1):57-60.[曹冬梅,康黎芳,王云山,等,2002. 根外施钾对苹果幼树气孔特性及光合速率的影响[J]. 山西农业科学,30(1):57-60.]
CHENG XF,2013. Overview of plant stomata and their movement mechanism[J]. Biol Teach,38(12):7-9.[成雪峰,2013. 植物气孔及其运动机理概述[J]. 生物学教学,38(12):7-9.]
COXSON DS,NADKARNI NM,1995. Ecological roles of epiphytes in nutrient cycles of forest ecosystems[M]//LOWMAN MD,NADKARNI NM. Forest Canopies. San Diego:Academic Press:495-543.
FRANKS PJ,BEERLING DJ,2009. Maximum leaf conductance driven by CO2 effects on stomatal size and density over geologic time[J]. Proc Natl Acad Sci USA,106(25):10343-10347.
FRANKS PJ,DRAKE PL,BEERLING DJ,2009. Plasticity in maximum stomatal conductance constrained by negative correlation between stomatal size and density:An analysis using Eucalyptus globulus[J]. Plant Cell Environ,32(12):1737-1748.
GENG YP,ZHANG WJ,LI B,et al.,2004. Phenotypic plasticity and invasiveness of alien plants[J]. Biodivers Sci,12(4): 447-455.[耿宇鹏,张文驹,李博,等,2004. 表型可塑性与外来植物的入侵能力[J]. 生物多样性,12(4): 447-455.]
HAO GY,WANG AY,LIU ZH,et al.,2011. Differentiation in light energy dissipation between hemiepiphytic and non-hemiepiphytic Ficus species with contrasting xylem hydraulic conductivity[J]. Tree Physiol,31(6):626-636.
HE J,HU JY,YANG DS,et al.,2007. Comparative study on stomatal density of Fagus pashanica seedlings in different habitats[J]. J Mianyang Norm Univ,26(11): 71-73.[贺静,胡进耀,杨冬生,等,2007. 不同生境的巴山水青冈幼苗气孔密度比较研究[J]. 绵阳师范学院学报,26(11): 71-73.]
HE RT,L CQ,1995. Effects of light on stomatal density of canopy leaves of several broad-leaved trees[J]. J Guangxi Agric Univ,14(4):311-316.[何若天,吕成群,1995. 若干阔叶树树冠各层叶气孔密度及光照条件对气孔密度的影响[J]. 广西农业大学学报,14(4):311-316.]
HETHERINGTON AM,WOODWARD FI,2003. The role of stomata in sensing and driving environmental change[J]. Nature,424(6951):901-908.
HIETZ P,BRIONES O,1998. Correlation between water relations and within-canopy distribution of epiphytic ferns in a Mexican cloud forest[J]. Oecologia,114(3):305-316.
HOLLAND N,RICHARDSON AD,GOODALE UM,et al.,2009. Stomatal length correlates with elevation of growth in four temperate species[J]. J Sustain For,28(12):63-73.
KUANG ML,ZHANG SB,2015. Physiological response to high light in Cymbidium tracyanum and C. sinense[J]. Plant Divers Resour,37(1):55-62.[匡美齡,张石宝,2015. 西藏虎头兰和墨兰对强光的生理响应[J]. 植物分类与资源学报,37(1):55-62.]
LAMMERTSMA EI,BOER HJ,DEKKER SC,et al.,2011. Global CO2 rise leads to reduced maximum stomatal conductance in Florida vegetation[J]. Proc Natl Acad Sci USA,108(10):4035-4040.
LEAO TCC,FONSECA CR,PERES CA,et al.,2014. Predicting extinction risk of Brazilian Atlantic forest angiosperms[J]. Conserv Biol,28(5):1349-1359.
LEE MJ,PARK SY,OH MM,et al.,2015. Growth and cell division of lettuce plants under various ratios of red to far-red light-emitting diodes[J]. Hort Environ Biotechnol,56(2): 186-194.
LI GC,HAN XG,HUANG JH,et al.,2001. Dry-season dynamics of soil inorganic nitrogen pools in primary Lithocarpus xylocarpus forest and degraded vegetations in Ailao Mountain,Yunnan Province[J]. Acta Phytoecol,25(2): 210-217.[李贵才,韩兴国,黄建辉,等,2001. 哀牢山木果柯林及其退化植被下土壤无机氮库的干季动态特征[J]. 植物生态学报,25(2):210-217.]
LI LH,ZHANG YP,YOU GY,et al.,2011. Spatiotemporal distribution pattern of photosynthetically active radiation in subtropical evergreen broadleaved forest in Ailaoshan Mountains of Southwest China[J]. Chin J Ecol,30(11):2394-2399.[李麟輝,张一平,游广永,等,2011. 哀牢山亚热带常绿阔叶林光合有效辐射的时空分布[J]. 生态学杂志,30(11):2394-2399.]
LIU CC,HE NP,ZHANG JH,et al.,2018. Variation of stomatal traits from cold temperate to tropical forests and association with water use efficiency[J]. Funct Ecol,32(1): 20-28.
L HF,2000. Comparative study on the stomatic distribution on plants Setereasea purpurea Boom.,Zebrina pendula Schnizl. and Commelina communis Linn.[J]. Chin Bull Bot,17(4):375-380.[吕洪飞,2000. 紫竹梅、吊竹梅和鸭跖草气孔分布与比较[J]. 植物学通报,17(4):375-380.]
MASLE J,GILMORE SR,FARQUHAR GD,2005. The ERECTA gene regulates plant transpiration efficiency in Arabidopsis[J]. Nature,436 (7052):866-870.
MAXWELL DP,FALK S,TRICK CC,et al.,1994. Growth at low temperature mimics high-light acclimation in Chlorella vulgaris[J]. Plant Physiol,105(2):535-543.
MENG L,CHEN WF,LI LX,et al.,2002. Influence of low light on stomatal characters in rice leaves[J]. J Shenyang Agric Univ,32(2):87-89.[孟雷,陈温福,李磊鑫,等,2002. 减弱光照强度对水稻叶片气孔性状的影响[J]. 沈阳农业大学学报,32(2):87-89.]
MCCRACKEN SF,FORSTNER MR,2014. Oil road effects on the anuran community of a high canopy tank bromeliad (Aechmea zebrina)in the upper Amazon basin,Ecuador[J]. PLoS ONE,9:e85470.
MONTERO A,ESTEVEZ J,PADILLA R,et al.,2009. Analytical performance of some methods for the determination of trace elements in lichens used as air quality assessment[J]. J Radioanal Nucl Chem,281(3):569-575.
NADKARNI NM,1984. Epiphyte biomass and nutrient capital of a Neotropical elfin forest[J].Biotropica,16(4):249-256.
NADKARNI NM,MATELSON TJ,1989. Bird use of epiphyte resources in Neotropical trees[J]. The Condor,91(4):891-907.
PIRES MV,ALMEIDA ALEX-ALAN FDE,ABREU PP,et al.,2012. Does shading explain variation in morphophysiological traits of tropical epiphytic orchids grown in artificial conditions?[J]. Acta Physiol Plant,34(6):2155-2164.
SCARANO FR,MATTOS EADE,FRANCO AC,et al.,2016. Features of CAM-cycling expressed in the dry season by terrestrial and epiphytic plants of Clusia arrudae Planchon & Triana in two rupestrian savannas of southeastern Brazil in comparison to the C3-species Eremanthus glomerulatus Less[J]. Trees,30(3):913-922.
SCHFER C,LTTGE U,1988. Effects of high irradiances on photosynthesis,growth and crassulacean acid metabolism in the epiphyte Kalancho uniflora[J]. Oecologia,75(4):567-574.
SONG J,LI RH,ZHU SD,et al.,2013. Leaf functional traits of ferns from different habitats in monsoon evergreen broad-leaved forest in Dinghushan Mountain[J]. J Trop Subtrop Bot,21(6):489-495.[宋娟,李榮华,朱师丹,等,2013. 鼎湖山季风常绿阔叶林不同生境蕨类植物的叶片功能性状研究[J]. 热带亚热带植物学报,21(6):489-495.]
SONG L,LIU WY,2011. Epiphytic plants:Their responses to global change and roles in bioindication[J]. Chin J Ecol,30(1):145-154.[宋亮,刘文耀,2011. 附生植物对全球变化的响应及其生物指示作用[J]. 生态学杂志,30(1): 145-154.]
SONG QH,FEI XH,ZHANG YP,et al.,2017. Snow damage strongly reduces the strength of the carbon sink in a primary subtropical evergreen broadleaved forest[J]. Environ Res Lett,12(10):104014.
SONG XY,HOGAN JA,LIN L,et al.,2018. Canopy openness and topographic habitat drive tree seedling recruitment after snow damage in an old-growth subtropical forest[J]. For Ecol Manag,429:493-502.
STRAUSS-DEBENEDETTI S,BAZZAZ FA,1991. Plasticity and acclimation to light in tropical Moraceae of different sucessional positions[J]. Oecologia,87(3):377-387.
STUNTZ S,ZOTZ G,2001. Photosynthesis in vascular epiphytes:a survey of 27 species of diverse taxonomic origin[J]. Flora,196(2):132-141.
SUN GY,ZOU Q,1991. Responses of photosynthetic rate and stomatal conductance to water stress in Soybean leaves[J]. Acta Bot Sin,33(1):43-49.[孙广玉,邹埼,1991. 大豆光合速率和气孔导度对水分胁迫的响应[J]. 植物学报,33(1):43-49.]
TAYLOY SH,FRANKS PJ,HULME SP,et al.,2012. Photosynthetic pathway and ecological adaptation explain stomatal trait diversity amongst grasses[J]. New Phytol,193(2):387-396.
TIAN M,YU GR,HE NP,et al.,2016. Leaf morphological and anatomical traits from tropical to temperate coniferous forests:mechanisms and influencing factors[J]. Sci Rep-uk,6:19703.
TICHA I,1982. Photosynthetic characteristics during ontogenesis of leaves VII. Stomatal density and sizes[J]. Photosynthetica,16(3):375-381.
UPRETY DC,DWIVEDI N,MOHAN JV,2002. Effect of elevated carbon dioxide concentration on the stomatal parameters of rice cultivars[J]. Photosynthetica,40(2):315-319.
VALLADARES F,WRIGHT SJ,LASSO E,et al.,2000. Plastic phenotypic response to light of 16 congeneric shrubs from a Panamanian rainforest[J]. Ecology,81(7):1925-1936.
WANG RL,YU GR,HE NP,et al.,2014. Elevation-related variation in leaf stomatal traits as a function of plant functional type:evidence from Changbai Mountain,China[J]. PLoS ONE,9(12):e115395.
WEN JW,CHEN HX,TENG YP,et al.,2018. Variation of leaf stomatal traits in Quercus species along the altitudinal gradient in Taibai Mountain,China[J]. Acta Ecol Sin,38(18): 6712-6721.[溫婧雯,陈昊轩,滕一平,等,2018. 太白山栎属树种气孔特征沿海拔梯度的变化规律[J]. 生态学报,38(18):6712-6721.]
WU ZY,1983. Research of forest ecosystem on Ailao Mountains[M]Kunming:Yunnan Science and Technology Press:17-35.[吴征镒,1983. 云南哀牢山森林生态系统研究[M]昆明:云南科学技术出版社:17-35.]
XU HQ,LIU WY,2005. Species diversity and distribution of epiphytes in the montane moist evergreen broad-leaved forest in Ailao Mountain,Yunnan[J]. Biodivers Sci,13(2):137-147.[徐海清,刘文耀,2005. 云南哀牢山山地湿性常绿阔叶林附生植物的多样性和分布[J]. 生物多样性,13(2): 137-147.]
YE ZP,YU Q,2008. Photosynthetic response to irradiance and CO2 concentration for flag leaves of winter wheat[J]. J Yangzhou Univ (Agric Life Sci Ed),29(3):33-37.[叶子飘,于强,2008. 冬小麦旗叶光合速率对光强度和CO2浓度的响应[J]. 扬州大学学报(农业与生命科学版),29(3): 33-37.]
ZHANG DP,1987. The study of rice leaf stomata Ⅰ,stomata density and length[J]. J Fujian Agron J,18(1):45-51.[张大鹏,1987. 水稻叶片气孔的研究 Ⅰ、气孔密度与长度[J]. 福建农学院学报,18(1):45-51.]
ZHANG Q,CHEN JW,CHEN YJ,et al.,2008. Photosynthetic induction in two fern species with different eco-types in Xishuangbanna tropical rainforest[J].Chin Bull Bot,25(6): 673-679.[张强,陈军文,陈亚军,等,2008. 西双版纳热带雨林中两种生态型蕨类植物的光合特性比较研究[J]. 植物学通报,25(6):673-679.]
ZHENG YL,JIANG CL,FENG YL,et al.,2005. Stomatal generation in plants[J]. Plant Physiol Commun,41(6):847-850.[郑玉龙,姜春玲,冯玉龙,等,2005. 植物的气孔发生[J]. 植物生理学通讯,41(6):847-850.]
ZHU YH,2013. Variation pattern of leaf traits of (Quercus variabilis)in East Asia and its response to environmental changes[D]. Shanghai:Shanghai Jiao Tong University:11-13.[朱燕华,2013. 东亚地区栓皮栎(Quercus variabilis)叶片性状的变异格局及其对环境变化的响应[D]. 上海:上海交通大学:11-13.]
ZOTZ G,2005. Vascular epiphytes in the temperate zones:A review[J]. Plant Ecol,176(2):173-193.
ZOTZ G,ANDRADE JOSE-LUIS,1998. Water relations of two co-occurring epiphytic bromeliads[J]. J Plant Physiol,152(4-5): 545-554.
ZOTZ G,2013. The systematic distribution of vascular epiphytes—A critical update[J]. Bot J Linn Soc,171(3):453-481.
(责任编辑 何永艳)
