粒肥施用时间对双季稻乳熟期亚细胞镉分布的影响
2021-08-06张玉盛张小毅肖欢敖和军向焱赟田伟肖峰
张玉盛,张小毅,肖欢,敖和军*,向焱赟,田伟,肖峰
(1.湖南农业大学农学院,长沙 410128;2.南方粮油作物协同创新中心,长沙 410128)
重金属镉(Cadmium,Cd)具有高毒性和迁移性,易被水稻吸收积累,通过食物链途径进入人体,危害人体健康。开展水稻镉低积累调控技术研究,对镉污染稻田水稻安全生产具有十分重要的意义。近几十年来,科研工作者就如何减少水稻镉积累开展了大量研究,并取得了可喜的成果。研究发现,营养调控[1−2]、土壤镉钝化[3]、生物修复[4]、品种筛选[5]等措施可降低水稻镉吸收和积累。其中,营养调控被认为是最经济、有效的降低作物镉积累的方法[6−7]。氮肥是农业生产中使用最多的肥料,氮在植物生长和发育过程中起着重要的作用,是植物主要营养物质和限制作物生长的最常见因子。氮肥调控作物镉的吸收、转运和积累,表现为:影响土壤理化性质,进而影响重金属迁移性和生物有效性;氮进入作物体内后,从分子、生化、细胞和整株(生物量和产量)等水平机制来平衡重金属毒性[8−9]。氮素营养对作物镉吸收表现为促进还是抑制作用与氮肥形态和剂量有关[10−11]。赵晶等[10]研究发现,施用氮肥较无肥处理均增加小麦对镉的吸收,不同氮肥处理之间存在显著差异,其中NH4Cl 对小麦吸收镉的促进作用最强,其次是NH4NO3和高量尿素处理,低量尿素处理和(NH4)2SO4处理小麦吸收镉最少。甲卡拉铁等[11]研究发现,不同氮肥对水稻增产效应呈现为尿素>(NH4)2SO4>NH4Cl>NH4NO3,相比于尿素、(NH4)2SO4和NH4NO3,NH4Cl能显著增加水稻镉含量,并促进镉由秸秆向籽粒中转移;适量的尿素能显著降低籽粒镉含量,而不施尿素和高量尿素显著提高籽粒镉含量。杨锚[12]研究发现,NH4Cl 处理、(NH4)2SO4处理相比尿素处理显著增加水稻单株镉积累量,而Ca(NO3)2处理降低水稻单株镉积累量,但与尿素处理间差异不显著。
水稻生育后期(抽穗以后)是籽粒镉积累的关键时期[13]。前期贮存在茎、叶等器官中的镉,在籽粒灌浆−成熟阶段通过“搭便车”的方式被输送至籽粒中,此外,后期根系吸收土壤中的镉也可被快速运输至籽粒[13−14]。有学者认为,在水稻镉积累关键时期采用适当的农艺措施降低土壤有效态镉含量及根系向上运输总量,即可有效降低糙米镉含量[13,15−16]。依据水稻生育后期对镉的吸收和转运特性,并结合前期研究结果,发现通过后期适量施氮可降低水稻镉积累[17]。本研究在此基础上进一步开展生育后期施氮控制水稻镉积累研究,试图建立通过后期施氮来调控水稻镉积累的农艺栽培技术。
1 材料与方法
1.1 试验品种与试验设计
试验于2018 年在湖南省浏阳市岩溪镇花园村进行,土壤类型为第四纪红壤发育的红黄泥水稻土、壤土,理化性状为:pH 5.48,有机质20.11 g·kg−1,全镉含量1.62 mg·kg−1,有效镉含量0.80 mg·kg−1。分别选用早稻株两优819 和陆两优996、晚稻湘晚籼12 号和玉针香为供试材料,经湖南杂交水稻研究中心筛选验证[5],株两优819、湘晚籼12 号为低镉积累品种,陆两优996、玉针香为高镉积累品种。
根据前期大田试验结果[17],双季稻全生育期施氮量180 kg·hm−2,以后期施氮量为总氮量的20%,设3个后期施氮时期处理和后期不施氮对照处理。处理1(F1)、处理2(F2)和处理3(F3)分别在始穗期、齐穗期(始穗后9 d)和灌浆期(齐穗后9 d)施氮,基肥∶穗肥∶粒肥均为6∶2∶2;对照(CK)为后期不施氮处理,基肥∶穗肥∶粒肥为8∶2∶0,每个处理3 次重复。水稻移栽前翻耕后反复多次耙平、耙匀,以减小试验小区间土壤镉含量的差异。小区随机排列,面积为8 m×8 m=64 m2,各小区间田埂覆膜以防肥水串灌。每个小区单独排灌。早稻收获后,在原有小区内用小型打田机翻耕均匀后种植晚稻,早稻于3 月22 日播种,4 月23日移栽,7月15日收获,株行距为16.6 cm×16.6 cm,晚稻于6 月22 日播种,7 月18 日移栽,10 月30 日收获,株行距为16.6 cm×20.0 cm。采用常规水肥管理,试验期间水稻施用氮肥180 kg·hm−2(尿素,N 46.4%)、磷肥90 kg·hm−2(过磷酸钙,P2O512%)、钾肥144 kg·hm−2(氯化钾,K2O 60%)。氮肥按试验基追肥比例在不同时期施用,磷肥一次性基施,钾肥于移栽前1~2 d和幼穗分化始期各施50%,早稻试验期间保持浅水层淹水灌溉(2~3 cm 水层),不晒田,晚稻收割前7 d 停止灌水,其余时期与早稻保持一致,其他栽培管理措施与当地习惯保持一致。
1.2 测定方法
1.2.1 亚细胞组分镉含量测定
采集乳熟期水稻植株鲜样用蒸馏水和超纯水洗净后用滤纸吸干表面水分,称取4 g根系、茎、叶,剪成1~2 mm2,按料液比1∶10 加入配制预冷的250 mmol·L−1蔗糖、50 mmol·L−1Tris−HCl、1 mmol·L−1DTT 共20 mL 冰浴研磨至匀浆,再加20 mL 冲洗研钵,匀浆与冲洗液同置于50 mL离心管中。采用差速离心,4 ℃、3 000 r·min离心15 min得到沉淀部分为细胞壁组分;将上清液以4 ℃、12 000 r·min 离心15 min 得到沉淀部分为细胞器组分;上清液为细胞液。混酸湿法(VHNO3∶VHClO4=4∶1)消解后采用电感耦合等离子体质谱仪(ICP−MS,Agilent 7700x,USA)测定各组分镉含量[1]。
1.2.2 植株镉含量测定
在水稻乳熟期、成熟期共进行2 次取样,避开小区边3行,采集田间长势一致的植株5穴,用自来水洗净根部与地上部,蒸馏水和超纯水润洗,晾干表面水分后,乳熟期植株分成根、茎、叶、穗四部分,成熟期分成根、茎叶、谷粒三部分,分别装入信封中,然后放入烘箱110 ℃杀青30 min,80 ℃烘干至恒质量,谷粒去糙,所有样品粉碎后过筛密封保存。混酸湿法(VHNO3∶VHClO4=4∶1)消解后,用电感耦合等离子体质谱仪(ICP−MS,Agilent 7700x,USA)测定镉含量。
1.3 数据处理
水稻成熟期根系、茎叶向籽粒转移镉的能力用转运系数(TF)表示。
TF根−茎叶=茎叶镉含量/根镉含量×100%,TF茎叶−糙米=糙米镉含量/茎叶镉含量×100%,TF根−糙米=糙米镉含量/根镉含量×100%[15]。
采用Excel 2016 和SPSS 25.0 进行数据整理统计分析和相关性分析,多重比较采用Duncan 法,相关性分析采用Pearson法。采用Excel 2016作图。
2 结果与分析
2.1 不同处理对水稻各时期不同部位镉含量及成熟期镉转移系数和产量的影响
2.1.1 早稻
除成熟期根和茎叶镉含量外,处理对乳熟期和成熟期各部位镉含量、镉转运系数和产量影响显著,而镉转运系数受品种、处理和品种互作影响不显著(表1)。由表1可知,株两优819乳熟期根、茎在不同处理下均有所提高,其中F2处理乳熟期根、茎镉显著高于其他处理,穗镉含量最低,F1和F3处理则表现为叶镉含量显著降低,穗镉含量提高。至成熟期,糙米镉含量在F1 处理下显著降低28.57%,F3 处理下提高14.29%。结合成熟期镉转运系数,说明施用粒肥可促进株两优819 根系对镉的吸收并向地上部转运,F1处理糙米镉含量显著降低,可能是因其产量显著提高,而“稀释”了糙米中镉含量。

表1 早稻不同时期各部位镉含量及成熟期镉转运系数和产量Table 1 Cd content of early rice of different parts in different stages and Cd transfer coefficient and yield in maturity stage
不同时期施用粒肥对陆两优996 乳熟期根镉含量都没有显著影响,从陆两优996 地上部镉含量来看,茎镉含量在F1 处理下最高,显著高于F2、F3 处理,叶和穗镉含量在F3 处理下显著高于其他处理。成熟期根、糙米镉含量在F3 处理下显著高于其他处理,茎叶镉含量显著高于CK、F2处理,其中F3处理糙米显著较CK 提高92.31%,在F1 和F2 处理下则分别降低23.08%、38.46%。其可能原因为,相比于F1 和F2处理,F3处理产量无显著变化,但其根系和茎叶中镉向籽粒的转运系数提高,致使其糙米镉含量显著提高。
2.1.2 晚稻
对晚稻而言,处理对乳熟期叶镉含量、产量影响不显著,品种对乳熟期根、茎和成熟期茎叶镉含量影响不显著,处理和品种互作对乳熟期根、叶镉含量及成熟期TF根−糙米影响不显著(表2)。不同时期施用粒肥可降低湘晚籼12号乳熟期各部位镉含量,且随粒肥施用时间的推移呈逐渐降低的趋势,尤其是叶和穗,两者镉含量较CK显著降低(表2)。湘晚籼12号糙米镉含量表现为F1>F3>CK>F2,F1 处理显著较CK 提高30.00%,F2 处理较CK 降低10.00%。究其原因,F1处理在乳熟期穗中已积累较高量的镉,在成熟期阶段,镉转运作用大于灌浆充实对籽粒镉的“稀释效应”,而F3处理产量显著提高,“稀释效应”增强,另外,其根和茎叶中镉向糙米中转运也低于CK。

表2 晚稻不同时期各部位镉含量及成熟期镉转运系数和产量Table 2 Cd content of late rice of different parts in different stages and Cd transfer coefficient and yield in maturity stage
玉针香乳熟期根、茎镉含量在粒肥处理下显著降低,分别降低39.57%~58.30%和48.78%~64.22%,且随粒肥施用时间推移,其降镉效应减弱。与湘晚籼12 号不同之处在于,粒肥处理显著提高了叶中镉含量77.78%~88.89%,即将更多的镉暂时“贮存”于叶当中,而后随叶衰老而重新分配转运至籽粒中。糙米镉含量在F2 处理下较CK 显著降低48.15%,F3 处理较F1、F2 处理显著提高47.62%~121.43%,较CK 提高14.84%。从产量角度来看,F1和F2处理产量与CK相比差异未达到显著水平,但根和茎叶中积累的镉低于CK,糙米中分配得到的镉随之减少,而F3 则反之,这可能是处理间糙米镉含量存在较大差异的原因。
2.2 后期施氮对双季稻乳熟期亚细胞各组分镉含量的影响
2.2.1 早稻
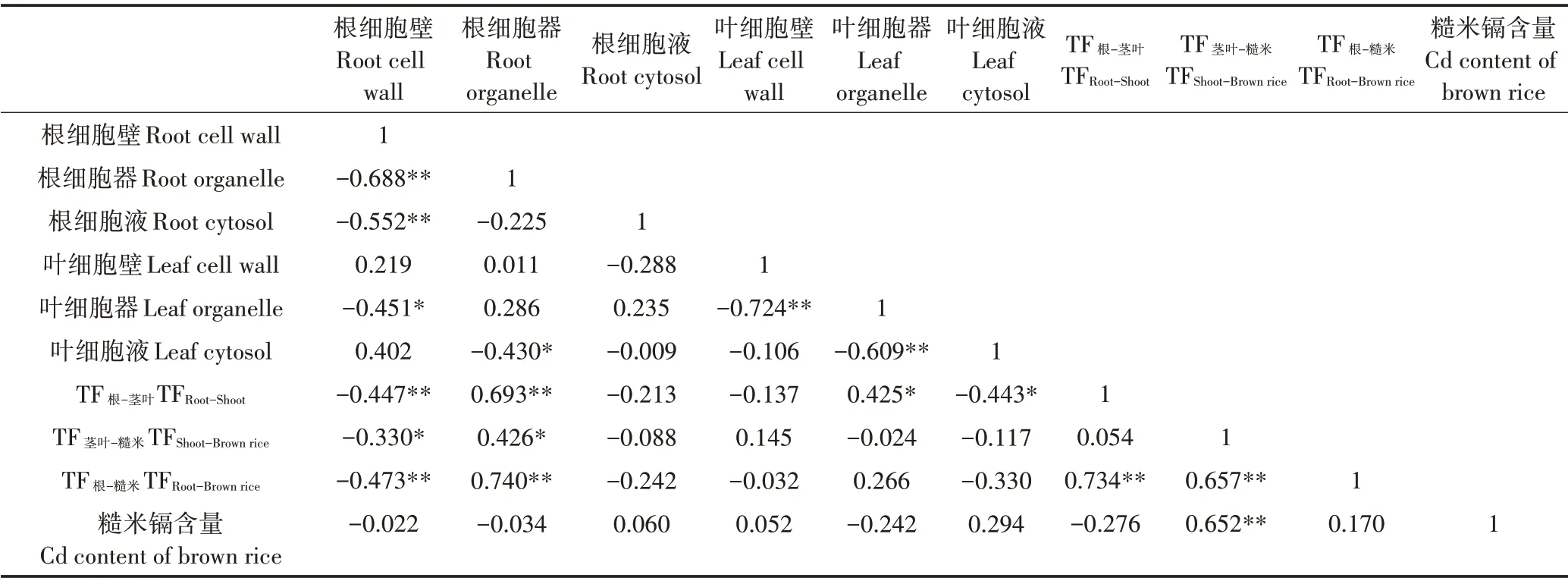
由表3 可知,早稻乳熟期根和叶细胞壁和细胞器镉含量受处理影响显著,品种对根细胞壁和细胞液镉含量及叶细胞器镉含量影响显著,处理和品种互作仅对根细胞液镉含量无显著影响,对根亚细胞其他组分和叶细胞各组分镉含量影响显著。镉在根亚细胞的分布整体上表现为细胞壁>细胞液>细胞器,且各组分的镉含量大于地上部分对应组分。株两优819 根细胞壁镉含量在F1~F3 处理下较CK 降低11.72%~62.50%,且F1 表3 早稻乳熟期根、叶亚细胞镉含量(mg·kg−1)Table 3 Subcellular Cd content of early rice root and leaf at milky stage(mg·kg−1) 叶亚细胞各组分镉含量在CK条件下表现为细胞壁>细胞液>细胞器,在粒肥处理下各组分镉含量变化较大。除细胞器F3 处理下降低25.00%外,株两优819 叶亚细胞各组分镉含量在粒肥处理下均有所提高。其中,叶细胞壁组分提高20.00%~280.00%,在F3处理下显著高于其他处理;细胞器组分在F1、F2处理下显著增加237.50%~262.50%;细胞液组分增加33.33%~66.66%,F2 处理与CK 差异显著。施用粒肥提高了陆两优996 叶细胞壁、细胞器组分镉含量,降低了细胞液镉含量,细胞壁、细胞器镉含量分别在F2、F1 处理下显著高于其他处理,各处理下细胞液镉含量无显著差异。 2.2.2 晚稻 由表4 可知,晚稻乳熟期根和叶亚细胞各组分镉含量受处理影响显著,品种仅对叶乳熟期细胞器镉含量影响显著,而处理和品种互作对根、叶细胞壁镉含量无显著影响,对其他两组分镉含量有显著影响。晚稻根和叶亚细胞各组分中的镉含量表现为细胞壁>细胞液>细胞器(表4)。湘晚籼12号和玉针香根亚细胞组分镉含量在粒肥处理下的变化趋势分别与株两优819 和陆两优996 基本保持一致,说明施用粒肥对不同稻季同一积累型水稻根亚细胞镉分布影响一致。湘晚籼12 号根亚细胞各组分在F1 处理下最低,细胞器和细胞液组分镉含量较CK 分别显著降低40.00%和49.77%,相比CK,F2、F3 处理对根细胞各组分镉含量无明显影响。玉针香根细胞壁组分镉含量表现为F2>CK>F1>F3,F2 较F1、F3 处理显著增加,细胞器组分在F2 处理下显著高于CK 和F1,细胞液组分在F3处理下显著高于其他处理。 表4 晚稻乳熟期根、叶亚细胞镉含量(mg·kg−1)Table 4 Subcellular Cd content of late rice root and leaf at milky stage(mg·kg−1) 湘晚籼12号叶细胞壁、细胞液组分镉含量在F1、F2 处理下较CK 无明显变化,在F3 处理下达到最高,其中F3 细胞液组分较CK 显著增加,这点与株两优819 一致,说明灌浆期施用粒肥可提高低镉积累品种细胞壁和细胞液组分对镉的固持作用;细胞器组分在F2、F3 处理下显著降低。施用粒肥对玉针香叶细胞壁组分镉含量无明显影响,细胞器镉含量在F2 处理下显著增加33.33%,细胞液镉含量在F2、F3 处理下亦显著增加。 后期不施氮情况下,镉在水稻根和叶中的分配比例为细胞壁>细胞液>细胞器(株两优819叶除外),后期施氮明显改变了镉在水稻乳熟期体内亚细胞中的分配比例(图1)。株两优819、陆两优996根细胞壁中镉分配比例在粒肥处理下降低,株两优819 细胞壁镉占比随氮施入时间推移呈增加趋势,陆两优则996 相反。株两优819、陆两优996 细胞器中镉的分配比例分别增加0.32~7.58、8.38~37.51 个百分点,其中陆两优996 细胞器中镉的分配比例在F2 处理时明显高于其他处理。株两优819 细胞液中的镉分配比例以F1最高,F2 次之,均高于CK;陆两优996 细胞液中镉分配比例在F1、F3 处理下高于CK。粒肥处理对镉在湘晚籼12 号根系中的分配比例影响较小,镉在细胞壁中的分配比例为53.33%~64.53%,细胞液中为26.27%~31.35%,细胞器中仅有6.36%~9.20%。在CK条件下,玉针香根亚细胞各组分镉分配比例分别为86.10%、2.41%和11.48%,在F1、F3 处理下细胞壁中镉占比分别降低17.11、50.09 个百分点,细胞器中镉分配比例在后期施氮处理下增加4.44~7.72 个百分点,细胞液中镉占比在F1、F3 处理下分别增加12.68、44.57个百分点。 株两优819 叶细胞壁组分镉占比在F1、F2 处理下降低19.94、19.61 个百分点,细胞器中镉分配比例在F1、F2 处理下增加27.49、22.92 个百分点;F3 处理下,细胞壁组分镉占比高达77.73%,而细胞器和细胞液中镉占比则明显降低。陆两优996 叶细胞壁中镉分配比例在F1、F2处理下明显提高,细胞器中镉占比在F3处理下增加30.19个百分点,细胞液镉占比在F3处理下降低17.67 个百分点。湘晚籼12 号、玉针香叶细胞壁、细胞器中镉分配比例在后期施氮处理下降低,细胞液中镉占比增加,湘晚籼12 号、玉针香叶细胞液组分镉分配比例分别在F3、F2时达到最高。 水稻亚细胞镉分配比例与镉转运系数和糙米镉含量之间相关分析见表5。根、叶亚细胞各组分中镉分配比例与糙米镉含量没有相关性,但与根、叶亚细胞各组分中镉分配比例之间及镉转运系数存在较好的相关性。其中,根细胞壁与根细胞器、根细胞液、叶细胞器、镉转运系数显著负相关,根细胞器与叶细胞液显著负相关,根细胞器与镉转运系数显著正相关。叶细胞壁与叶细胞器极显著负相关,叶细胞器和叶细胞液极显著负相关,叶细胞器和TF根−茎叶显著正相关,叶细胞液和TF根−茎叶显著负相关。TF根−茎叶、TF茎叶−糙米两者与TF根−糙米极显著正相关,TF茎叶−糙米与糙米镉含量极显著正相关。由此可知,降低根细胞器中镉分配比例有利于减少水稻植株中镉向籽粒中转移,而糙米镉含量主要取决于茎叶中镉向籽粒中转运。 表5 水稻亚细胞镉分配比例与镉转运系数和糙米镉含量之间相关性分析Table 5 Correlation analysis of distribution ration of Cd in subcellular and Cd content of brown rice and Cd transfer coefficient 水稻籽粒灌浆所需的光合同化产物来自于花后光合作用和开花前储存在茎和其他器官的非结构性碳水化合物,后者对籽粒产量的贡献约占1/6~1/3,其量的多寡取决于生长条件和氮素供应水平,此外,花前储存物质还是启动灌浆的重要物质基础,其转运速率和转运量对灌浆初期籽粒库的活性起重要作用[18−19]。水稻齐穗后,生长环境中有效镉含量较低时,籽粒镉来源于前期储存在根系和茎秆中的镉和根系吸收与直接运输;生长环境中有效镉含量较丰富条件下,籽粒镉主要来源于生长介质,少部分来自各器官的转移[13]。灌浆时水稻物质运输、转运及合成较快,水稻根系在此阶段吸收的镉可被快速运输至籽粒中[13]。本研究发现,不同时期施用粒肥对不同稻季低镉积累品种糙米镉含量影响存在差异,早稻低镉积累品种株两优819 在始穗期施氮处理下糙米镉含量显著降低,晚稻低镉积累品种湘晚籼12 号则显著提高,齐穗期和灌浆期施氮对两者糙米镉含量没有明显影响。不同稻季高镉积累品种糙米镉含量均在齐穗期施氮处理下显著降低,灌浆期施氮处理下显著提高。说明在齐穗期施氮对低镉品种糙米镉含量影响较小,但可显著降低高镉品种的糙米镉含量,在灌浆期施氮则促进稻米镉积累。 尿素施入土壤后,经土壤微生物作用生成的脲酶水解生成氨和含氮氧化物,可在短期内提高土壤pH,而后发生硝化反应,酸化土壤,使土壤中镉的活性提高[8,20]。杨锚等[21]研究发现,在淹水条件下,施入尿素可显著提高土壤pH 值,显著降低土壤水溶态镉含量和有效态镉含量,随培养时间延长,水溶态镉含量降低幅度和土壤pH 值提升幅度减弱。另有报道指出,施入适量尿素5 d 后,土壤pH 显著提高,而后缓慢回落趋于中性,在60 d 时显著降低,土壤有效镉含量与土壤pH 值呈线性负相关,籽粒镉含量显著降低[11,22]。本研究中,F1、F2处理相比于F3处理,施氮时间前移,在尿素施入土壤后,土壤pH 经历“先升后降”的变化趋势,可致使水稻在此期间镉积累量减少,进而使籽粒在灌浆至成熟阶段分配、转运得到的镉总量降低,这可能是灌浆期施氮(F3)处理下,株两优819、陆两优996糙米镉含量高于始穗期(F1)和齐穗期施氮(F2)的原因之一;此外,F3 处理促进了根和茎叶中的镉向籽粒转移(TF茎叶−糙米和TF根−糙米显著提高,表1)。对湘晚籼12 号、玉针香而言,其根和茎叶中的镉向籽粒转运虽降低(表2),但因其前期积累在根、茎叶中的镉显著高于其他处理,致使籽粒在后期分配得到的镉增加。 植物对重金属的耐受性与植物对重金属的吸收运输、重金属在植物细胞各部位的分布及与体内物质的结合形态密切相关,细胞壁固持和液泡区隔化在提高植物对重金属的耐性和富集能力方面起着主要作用。养分输入对水稻细胞各组分中镉的分布影响显著[23−27]。适量施用尿素,一方面可以在短期内降低土壤有效镉含量,降低水稻在这期间的镉积累量,另一方面,细胞壁中富含果胶、纤维素等多糖分子及羟基、羧基、醛基和磷酸基等亲金属离子的配位基团[28],施氮可增加根细胞壁中果胶和半纤维素含量,增加金属硫蛋白、植物螯合蛋白合成酶和磷酸烯醇式丙酮酸羧化酶,促进细胞壁和液泡对镉的固定、区隔化,减少镉向细胞器运输[9]。本研究发现,在CK 条件下,4 个品种乳熟期根和叶细胞中镉含量和分配比例均表现为细胞壁>细胞液>细胞器,这与前人研究结果一致[24]。在早稻季中,低镉积累品种株两优819 根细胞壁、细胞液镉含量显著高于高镉积累品种陆两优996(表3),这与唐杰等[29]研究结果一致,而晚稻季不同积累型品种根细胞各组分镉含量并无明显差异。水稻根系直接与外界环境接触,是水稻从外界环境中吸收所需养分最关键的部位,土壤中的镉可通过占用锌、铁、锰等特异度较低的阳离子通道进入植株体内,镉离子还可以通过锌、钙转运通道与锌转运蛋白和钙转运蛋白结合,以共质体途径进入根表皮细胞[30]。进入细胞内的镉可对原生质体产生毒害作用,例如破坏叶绿体膜、破坏线粒体膜、损伤细胞核仁、抑制核糖核酸酶活性、改变RNA 合成等[31]。根系细胞壁是水稻防御镉毒害的第一道屏障,细胞器是主要生理场所,提高根细胞壁中镉分配比例,有利于减少镉向地上部的转运。本研究发现,根细胞器组分中镉分配比例与镉转运系数显著正相关(表5),始穗期施氮(F1)处理下双季稻4 个品种乳熟期根细胞器组分镉含量最低;理论上来讲,F1 处理下糙米镉含量理应最低,而事实上,仅株两优819糙米镉含量在F1处理下相符,其原因可能为株两优819产量在F1处理下显著提高,对糙米中镉起“稀释”作用。因此,在大田生产中,可考虑从提高水稻细胞耐镉性和产量出发,在抽穗至灌浆期间适量施氮来调控水稻籽粒镉含量。 (1)始穗期施氮可显著降低低镉积累品种株两优819 糙米镉含量,齐穗期施氮可显著降低高镉积累品种陆两优996 和玉针香糙米镉含量,灌浆期施氮提高糙米镉含量。 (2)始穗期至灌浆期阶段施氮可提高水稻乳熟期根细胞器中的镉分配比例;根细胞壁中镉的分配比例与镉的转运系数显著负相关,根细胞器中镉的分配比例与镉的转运系数显著正相关。

2.3 后期施氮对双季稻乳熟期亚细胞各组分镉分配比例的影响
2.4 相关性分析
3 讨论
4 结论
