MEK5α 和MEK5β 调控Beclin 1 启动子
2021-03-01刘晓芸王雷斌张沙沙赵微苗朱洪新
刘晓芸,王雷斌,王 庆,张沙沙,赵微苗,贺 林,朱洪新
(上海交通大学Bio-X 研究院遗传发育与精神神经疾病教育部重点实验室,上海 200240)
巨自噬(简称自噬)是真核细胞内一种保守的溶酶体降解路径,存在于体内几乎所有的细胞中[1].自噬可人为地分为自噬起始、自噬体形成、自噬体成熟及溶酶体内降解等几个阶段.目前,对自噬的分子机制已有了比较明确的认识.在哺乳动物细胞自噬起始阶段,ULK1-ATG13-FIP200 复合物中,ULK1 激活Beclin 1-VPS34-Atg14L 复合物组分VPS34,促进PI3P 的合成,从而诱导自噬泡的形成.然后,细胞内形成泛素样复合物Atg12-Atg5-Atg16L,行使泛素连接酶的功能,促进LC3-PE(也称为LC3 Ⅱ)与自噬体外膜结合,并募集其他自噬体膜蛋白促进自噬体形成.自噬体形成后与溶酶体外膜融合(自噬体成熟),自噬体内膜及内容物被溶酶体水解酶降解并进入细胞质加以再利用.自噬体成熟受Beclin 1-VPS34-UVRAG 复合物调控[2].因此,Beclin 1在细胞自噬起始及自噬体成熟阶段起着关键作用.
Beclin 1 是哺乳动物的酵母Atg6 同源基因,也是最早发现的哺乳动物自噬相关基因[3].人Beclin 1 基因位于17q21,含12 个外显子和11 个内含子.Beclin 1 转录产物为2 098 个碱基,包括120 个碱基的5′末端未翻译区,1 353 碱基的编码区和625 个碱基的3′末端未翻译区[4].在蛋白结构上,Beclin 1有一个Bcl-2 同源结构域3(Bcl-2 homology domain 3,BH3),一个coiled-coil 结构域(coiled-coil domain,CCD)及一个进化保守结构域(evolutionarily conserved domain,ECD).这些结构域的存在使Beclin 1 可以与多个蛋白质如VPS34 及UVRAG 形成复合物行使相应的分子功能[5].
MEK5 是MEK 家族成员,由选择性剪切产生MEK5α 和MEK5β,相应合成的蛋白质分别为50 和40 kDa[6].MEK5α 主要在脑和肝脏表达,而MEK5β 在多种组织器官广泛表达[6].已有研究表明,MEK5α 可结合并激活ERK5.MEK5α 也是迄今为止发现的唯一能够特异性激活ERK5 的上游激酶.而MEK5β 抑制MEK5α与ERK5 结合从而抑制MEK5α 激活ERK5.因此,MEK5α和MEK5β 对于ERK5 的激活具有拮抗作用[7].
目前,已有研究发现MAPK 家族信号通路如MEK-ERK 及JNK可调控Beclin 1 的表达.而MEK5-ERK5 信号通路对Beclin 1 的表达调控尚不清楚[8-10].Beclin 1 在肌肉分化的过程中表达上调,并调控心肌细胞及骨骼肌细胞分化[11-13].已有研究发现,在肌肉分化过程中,MEK5-ERK5 信号通路活性增强,并调控成肌细胞分化[14-16].本工作推测MEK5-ERK5 信号通路可能通过调控Beclin 1 基因表达从而调控肌肉分化,并主要研究MEK5 对于成肌细胞Beclin 1 启动子活性的调控.
1 材料与方法
1.1 试剂
PyrobestTMDNA Polymerase 及T4 DNA 连接酶购自Takara 公司.限制性内切酶KpnⅠ、HindⅢ及SalⅠ购自New England Biolabs(NEB)公司.1 kb plus DNA ladder 购自Invitrogen 公司.C2C12 细胞购自美国菌种保藏中心(American type culture collection,ATCC).胎牛血清及DMEM(高糖)购自Gibco 公司,Lipofectamine 2000 购自Invitrogen 公司.Dual Luciferase Reporter Assay System 购自Promega 公司.
1.2 质粒
PCFG5-DD(持续活性 MEK5β),PCFG5-AA(负显性 MEK5β)及 PCFG5-WT(野生 型MEK5β)由Stephan Ludwig 教授(Universitaetsklinikum Muenster)馈 赠.pCMVMEK5αCA(持续活性MEK5α)及pCMV-MEK5αDN(负显性MEK5α)由Xia Zhengui 教授(University of Washington)馈赠.pS6(pCMV.SPORT6),pS6-CREB3,pS6-CREBP,pS6-CREBL1 及pS6-E2F-1 由马钢副教授及郭熙志教授(上海交通大学)提供.
1.3 DNA 片段克隆与亚克隆
用SalⅠ和Hind Ⅲ双酶切PCFG5-DD,将获得的MEK5βDD 片段亚克隆至pDNA3.1(−)质粒,得到pcDNA3.1-MEK5βDD(简称MEK5βDD),并进行酶切鉴定和测序鉴定.用聚合酶链式反应(polymerase chain reaction,PCR)扩增PCFG5-AA及PCFG5-WT中的MEK5β片段,将其亚克隆至pDNA3.1(−)质粒,分别获得pcDNA3.1-MEK5βAA(简称MEK5βAA)及pcDNA3.1-MEK5βWT(简称MEK5βWT).然后分别进行酶切鉴定和测序鉴定.以鼠尾DNA 为模板,用PCR 扩增Beclin 1 起始密码子上游2 088 碱基对区域,并将其克隆至pGL3-Basic 载体KpnⅠ及HindⅢ酶切位点,进行酶切鉴定及测序鉴定.由于Beclin 1转录起始位点位于起始密码子上游232 碱基对位置,因此将含有2 088 碱基对片段的pGL3-Basic 载体命名为p-1 856.类似地,以p-1 856 为模板,用PCR 分别扩增起始密码子上游1 870,1 613,1 448,1 207,1 023,586 及373 碱基对区域并克隆至pGL3-Basic 质粒KpnⅠ及HindⅢ酶切位点,分别获得pGL3-Basic-1 638 (p-1 638),pGL3-Basic-1 381 (p-1 381),pGL3-Basic-1 216 (p-1 216),pGL3-Basic-975(p-975),pGL3-Basic-791(p-791),pGL3-Basic-354(p-354)及pGL3-Basic-141(p-141)载体,进行酶切鉴定及测序鉴定后分析启动子活性.不同长度Beclin 1 启动子片段PCR 扩增引物如表1所示.

表1 小鼠Beclin 1 启动子片段PCR 扩增引物Table 1 Sequences of primers used for PCR-amplification of mouse Beclin 1 promoter fragments
1.4 细胞培养与细胞转染
C2C12 细胞培养于含10%胎牛血清的DMEM(高糖)培养液.将细胞接种至24 孔板,待次日细胞密度达到80%∼90%时用lipofectamine 2000 进行细胞转染.转染操作按lipofectamine 2000 使用说明书进行.如无特殊说明,0.8 µg 质粒DNA 用于细胞转染,转染后24∼48 h 进行荧光素酶报告基因检测实验.
1.5 荧光素酶报告基因检测
采用Promega 公司Dual Luciferase Reporter Assay System 进行荧光素酶报告基因测定,具体操作按说明书进行.
1.6 Real-time RT-PCR
使用TRIzol 试剂提取转染细胞总RNA,并将细胞总RNA 反转录合成cDNA(GENEray,GK8030-50)后用 real-time PCR(RT-PCR) 方法分析基因表达(GENEray,GK8020-200).用于real-time PCR 的Beclin 1 基因正向引物序列为5′-GCTCCTGTGGAATGGAATGA-3′,反向引物序列为5′-GAACAGTACAACGGCAACTC-3′;内参基因Gapdh正向引物序列为5′-GTGTTCCTACCCCCAATGT-3′,反向引物序列为5′-TGTCATCATACTTGGCAGGTTTC-3′.
1.7 统计学方法
数据均以mean±SD 表示,两组间平均值比较采用student t 检验.多组间平均值比较采用单因素方差分析,P <0.05 表示差异具有显著性.
2 结果
2.1 不同Beclin 1 启动子片段的活性分析
为了确定Beclin 1 启动子活性区域,首先将小鼠Beclin 1 起始密码子上游2 088 碱基对区域克隆至pGL3-Basic 质粒中(p-1 856).将p-1 856 或pGL3-Basic 转染至C2C12 细胞,双荧光素酶报告基因检测实验结果显示p-1 856 荧光素酶活性显著增加(见图1,图中M 代表1 kb plus DNA ladder),表明Beclin 1 基因−1 856 区域具有启动子活性.为了获得Beclin 1 启动子关键区域,将Beclin 1 启动子−1 856 片段5′端删除不同长度后亚克隆至pGL3-Basic,并进行启动子活性分析.实验结果表明,p-354 荧光素酶活性最高,而p-791 荧光素酶活性显著下降,p-141 荧光素酶活性最低(见图1).上述结果表明,−354 ∼−141 区域是Beclin 1 启动子活性的关键区域.因此,接下来将使用p-354 进一步研究Beclin 1 启动子的调控.
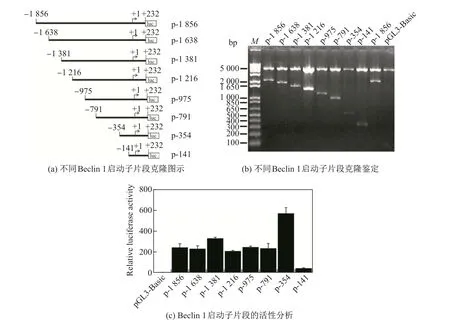
图1 小鼠Beclin 1 启动子的克隆与活性分析Fig.1 Cloning and activity analysis of Beclin 1 promoters in mice
2.2 MEK5α 与MEK5β对Beclin 1 启动子活性的调控
为了研究 MEK5 对 Beclin 1 启动子活性的调控,首先将活性形式的MEK5α(MEK5αCA)或无活性形式的MEK5α(MEK5αDN)分别与p-354 共转染至C2C12细胞.双荧光素酶报告基因检测实验结果显示,MEK5αCA 显著上调荧光素酶活性,而MEK5αDN 显著下调荧光素酶活性(见图2(a)),表明MEK5α能够正向调控Beclin 1 启动子活性.然后分别转染不同量的MEK5αCA,结果表明少量MEK5αCA(0.3 µg)对Beclin 1启动子活性无显著作用,中量MEK5αCA(0.8 µg)可显著上调Beclin 1 启动子活性,而多量MEK5αCA(2.4 µg)可进一步上调Beclin 1 启动子活性(见图2(b)).因此,MEK5αCA对于Beclin 1 启动子活性的调控具有剂量依赖关系.为了研究MEK5β对Beclin 1 启动子活性的调控,分别将活性MEK5β (MEK5βDD),野生型MEK5β(MEK5βWT)及无活性MEK5β(MEK5βAA)与p-354 共转染至C2C12 细胞.双荧光素酶报告基因检测实验结果显示,尽管MEK5βAA 对p-354 活性无显著影响,但MEK5βWT 及MEK5βDD 显著下调Beclin 1 启动子活性(见图2(c)),表明MEK5β 负向调控Beclin 1 启动子活性.为了揭示MEK5α 与MEK5β对Beclin 1 启动子活性调控是否具有拮抗作用,将p-354,MEK5αCA 与MEK5βWT/MEK5βAA/MEK5βDD 分别共转染至C2C12 细胞.双荧光素酶报告基因检测实验结果显示,共转染MEK5βDD 和MEK5αCA 导致p-354 的荧光素酶活性显著降低(见图2(d)),表明MEK5βDD 能够抑制MEK5αCA对Beclin 1 启动子活性的正向调控.

图2 MEK5α 与MEK5β对Beclin 1 启动子的调控Fig.2 MEK5α and MEK5β regulated Beclin 1 promoters
2.3 MEK5α 与MEK5β 调控Beclin 1 mRNA 表达
为了验证MEK5α 与MEK5β 能够调控细胞Beclin 1 基因表达,将MEK5αCA 与MEK5βDD单独或共转染细胞,用real-time PCR 方法检测Beclin 1 mRNA的表达.由图3可见,单独转染MEK5αCA 显著上调Beclin 1 mRNA 表达,单独转染MEK5βDD 显著下调Beclin 1 mRNA 表达.在MEK5βDD 与MEK5αCA 共转染细胞,MEK5βDD 则显著抑制MEK5αCA 对Beclin 1 mRNA 表达的促进作用.这些结果说明MEK5α 与MEK5β分别正向和负向调控Beclin 1 mRNA表达,二者具有拮抗作用,这与MEK5α 与MEK5β对Beclin 1 启动子的调控结果一致.

图3 MEK5α与MEK5β调控Beclin 1 mRNA 表达Fig.3 MEK5α and MEK5β regulated the expression of Beclin 1 mRNA
2.4 MEK5α 与CREB 对Beclin 1 启动子活性调控具有协同效应
在p-354 启动子区域存在CREB 结合位点,而MEK5-ERK5 信号通路能够调控CREB 活性.因此,MEK5-ERK5 可能通过CREB 调控Beclin 1启动子活性.首先研究CREB 是否能够调控p-354 活性,将CREB 家族成员CREB3,CREBBP 及CREBL1 分别与p-354 共转染至C2C12 细胞,双荧光素酶报告基因检测实验结果显示,CREB3,CREBBP 及CREBL1 均能够显著上调p-354 荧光素酶活性(见图4(a)).将不同量的CREB3 与p-354 共转染至C2C12细胞,双荧光素酶报告基因检测实验结果显示,转染少量CREB3(0.3 µg)对p-354 荧光素酶活性无显著影响,而转染中量CREB3(0.8 µg)显著上调p-354 荧光素酶活性,转染多量CREB3(2.4 µg) 也显著上调p-354 荧光素酶活性,但与转染中量CREB3 无显著差别(见图4(b)),证明CREB 能够上调Beclin 1 启动子活性.接下来将少量CREB(0.3 µg)或MEK5αCA (0.3 µg)分别与p-354 共转染至C2C12 细胞,或者将少量CREB (0.3 µg)和MEK5αCA(0.3 µg)与p-354 共转染至C2C12 细胞.双荧光素酶报告基因检测实验结果显示,单独转染少量CREB3,CREBP,CREBL1 或MEK5αCA 对p-354 荧光素酶活性无显著影响,但共转染少量CREB 和MEK5αCA 可显著上调p-354 荧光素酶活性(见图4(c)).上述结果表明MEK5α 与CREB 对Beclin 1 启动子的调控具有协同效应.

图4 MEK5α 与CREB对Beclin 1 启动子调控的协同效应Fig.4 Synergistic effects of MEK5α and CREB on Beclin 1 promoter
2.5 E2F-1 调控Beclin 1 启动子活性
在p-1 381 与p-1 216 之间存在E2F-1 转录因子结合位点,因此选用长Beclin 1 启动子区域p-1 856 研究E2F-1 对Beclin 1 启动子的调控.将E2F-1 及p-1 856 共转染至C2C12 细胞.双荧光素酶报告基因检测实验结果显示,E2F-1 显著上调p-1 856 荧光素酶活性(见图5),表明E2F-1 能够调控Beclin 1 启动子活性.MEK5 是否能够通过E2F-1 调控Beclin 1 启动子活性则有待进一步研究.

图5 E2F-1 促进Beclin 1 启动子活性Fig.5 E2F-1 facilitated Beclin 1 promoter activity
MEK5α 通过活化ERK5 促进CREB 活性,进而上调Beclin 1 基因表达.MEK5β能够抑制MEK5α 对ERK5 的激活从而抑制MEK5α 对Beclin 1 的表达调控.E2F-1 能促进Beclin 1基因表达.
3 讨论
本工作主要发现如下:①Beclin 1 转录起始位点上游354 碱基对区域是Beclin 1 启动子的关键活性区域;②MEK5α 和MEK5β 对Beclin 1 启动子的调控具有相互拮抗作用;③CREB 促进Beclin 1 启动子活性,并与MEK5α 具有协同效应.
在Beclin 1 起始密码子上游约2 kb 碱基对区域,Beclin 1转录起始位点上游354 碱基对区域具有最大启动子活性,而转录起始位点上游141 碱基对区域启动子活性最低.因此,Beclin 1 启动子活性的关键区域位于转录起始位点上游354 碱基对区域与141 碱基对区域之间.Beclin 1 转录起始位点上游791 碱基对区域启动子活性下降的原因可能是存在转录抑制因子结合位点,这有待进一步鉴定.Beclin 1 转录起始位点上游1 381 碱基对与1 216 碱基对之间区域存在E2F-1 结合位点.E2F-1 能够促进Beclin 1 启动子活性,这可能是−1 381 片段比−1 216 片段启动子活性强的原因.
本工作发现MEK5α 和MEK5β 对Beclin 1 启动子具有相反的调控作用,并且在mRNA水平上证实了MEK5α 和MEK5β对Beclin 1 表达的拮抗调控.已有研究中MEK5α能够与ERK5 结合并激活ERK5,而MEK5β 抑制MEK5α与ERK5 结合从而抑制MEK5α 诱导的ERK5 活化[7].因此,可认为MEK5α 通过活化ERK5 上调Beclin 1 基因表达.MEK5β 能够抑制MEK5α 对ERK5 的激活从而抑制MEK5α 对Beclin 1 的表达调控(见图6).鉴于−354 碱基对区域Beclin 1 启动子片段活性最强,−141 碱基对区域Beclin 1 启动子片段活性最弱,而CREB 结合位点位于−354 碱基对区域与−141 碱基对区域之间,并能剂量依赖性调控Beclin 1 启动子活性,这提示CREB 可能是调控Beclin 1 表达的关键因子.本工作发现MEK5α 与CREB 对Beclin 1 启动子活化具有协同效应,这与文献[17]发现的MEK5-ERK5信号通路能够活化CREB 一致.因此,CREB 可能是MEK5-ERK5 信号通路调控Beclin 1表达的关键下游效应因子.此外,本工作还发现E2F-1 能够促进Beclin 1 启动子活性,这与Weinmann 等[18]的报道一致.目前尚不清楚MEK5α 和MEK5β 是否可以直接调控E2F-1的活性.但已有报道表明CREB 能够上调E2F-1 的表达[19],因此MEK5-ERK5 有可能通过CREB 间接调控E2F-1 的活性,从而调控Beclin 1 基因表达.

图6 MEK5 调控Beclin 1 基因表达模式Fig.6 Pattern of MEK5 regulating Beclin 1 expression
Beclin 1 在生物体内广泛表达,能够调控多种生物学过程如细胞自噬、增殖、分化和凋亡[20].本工作使用小鼠成肌细胞C2C12 研究了MEK5 对Beclin 1 启动子活性的调控.Beclin 1 在肌肉细胞分化过程中表达增加,自噬上调[11-13].因此,本工作对于肌肉细胞分化调控研究具有一定意义,也对Beclin 1 调控其他生物学过程的研究具有一定的启示.此外,Beclin 1 在多种疾病如肿瘤、心脏疾病及神经系统疾病的发生发展中均起着重要作用[21-23],因此本工作对于与Beclin 1 相关的疾病研究也具有一定意义.
