基于温室环境和作物生长的番茄基质栽培灌溉模型
2020-07-10徐立鸿肖康俊蔚瑞华
徐立鸿,肖康俊,蔚瑞华
·农业生物环境与能源工程·
基于温室环境和作物生长的番茄基质栽培灌溉模型
徐立鸿,肖康俊,蔚瑞华
(同济大学电子与信息工程学院,上海 201804)
为解决涵盖土壤蒸发和作物冠层蒸腾的土培作物蒸散模型不能直接应用于稻壳炭基质栽培番茄灌溉的问题,该研究首先通过修改Penman-Monteith模型的原始表达式来去除土壤蒸发部分,并引入TOMGRO模型来模拟番茄冠层生长,给出了阻抗参数的修正计算,得到了新的番茄基质栽培蒸腾模型。考虑到蒸腾模型中净辐射项削弱了室外太阳辐射对冠层及以下部整株植株的耗水影响,进而将新的蒸腾模型与太阳辐射线性比例供水模型结合建立蒸腾-辐射综合灌溉模型。结果表明,蒸腾-辐射综合灌溉模型对上海崇明A8温室番茄灌溉量的模拟结果与实际结果之间的相关系数高于0.95,平均相对误差小于20%。这说明蒸腾-辐射综合灌溉模型能够较好地估算温室稻壳炭基质栽培番茄的灌溉需水量,对深入研究温室灌溉实施具有参考价值。
温室;灌溉;番茄;基质栽培;Penman-Monteith模型;蒸腾作用
0 引 言
番茄是世界上最重要以及种植面积最广泛的园艺作物[1],同时也是中国农业生产过程中大面积种植且产量较高的蔬果之一[2]。无论大田种植还是温室栽培,在整个生长周期中适宜的水分供给对番茄的生长发育起着至关重要的作用[3]。不合理的供水情况会对番茄生产效益产生不利影响;例如,供水过少会造成作物根系水分胁迫,而供水过多会造成根系供氧不足且会使得果实开裂或掉落[4]。在无土基质栽培中,由于基质蒸发较小且持水能力较弱,其水分控制对于防止作物水分胁迫非常重要,这就使得准确估算基质栽培作物的灌溉需水情况十分必要[5]。
在水文生态学中,作物需水量可与作物蒸散量(Evapotranspiration)互换使用,因而在无法准确测量该水文参数的情况下,估算作物蒸散量则是短期预测实际作物需水量的关键环节[6]。
联合国粮食及农业组织(FAO)早于1998年以能量平衡和湍流传导原理来修正Penman模型,提出了适用于大田作物需水量估算的Penman-Monteith蒸散模型,并且给出了以经验作物系数将参考作物蒸散量转换为实际蒸散量来反映作物实际水分需求的方式[7]。对于自然通风或不频繁通风的日光温室,由于室内空气流通较为缓慢且风速几乎为0,则会造成以作物几何特征(零平面位移高度和粗糙度长度)和风速相关的对数函数直接计算的空气动力阻抗趋于无限大。因此,在张培新等[8-9]修正空气动力阻抗中风速项的基础上,刘浩等[10]建立了一个需要日光温室气象数据和作物生长指标的温室土培番茄蒸散量估算模型。更重要的是,在引入草基准面简化Penman-Monteith模型原始表示过程中将叶片表层阻抗作为固定值,区别于这种经验处理无法反映冠层叶片在不同生长阶段受到太阳辐射影响而变化的情况,上述蒸散修正模型引入了Boulard等[11]提出的基于太阳辐射计算单叶气孔阻力,以此来模拟叶片生长变化下受辐射影响的表层阻抗。
尽管许多研究依据实际温室环境进行了阻抗参数的合理修正,使得Penman-Monteith蒸散模型适用于温室土培作物冠层蒸腾量的估算,但这些修正模型在参数计算过程中并未将土壤蒸发部分从蒸散(包含了土壤蒸发和作物冠层蒸腾2个部分)中去除。然而,在西班牙东南部某塑料温室开放基质栽培的模拟番茄蒸腾量的研究中,由于土壤被沙覆盖层所覆盖,PrHo模型(PrHo是一种可用作温室蔬菜作物灌溉计划决策支持系统的软件,其中计算作物蒸散量的模型被称作PrHo模型)所模拟的蒸散量被认为是土壤栽培蔬菜作物的蒸腾量[12-13],这就意味着覆盖土壤处理相当于移除土壤蒸发的影响。在无土栽培辣椒的有效灌溉研究中,Shin等[14]指出基质表面的蒸发很小甚至可以忽略不计。此外,在一些无土基质栽培作物的蒸腾量估算和灌溉管理的研究中[15-17],“蒸腾”一词用于反映作物的需水量,而非包含了土壤蒸发的“蒸散”。因而,在不考虑土壤蒸发量的情况下,利用Penman-Monteith参考蒸散模型及其修正模型来计算温室有机基质栽培作物的冠层蒸腾量具有一定的挑战性[3]。
在无土基质栽培的灌溉管理实践中,传统的灌溉方式是基于室外太阳辐射总量与作物蒸腾的线性比例关系来估算供水量[18],类似的灌溉模型则是基于年内天数和当日辐射总量的线性表式[19],而这些计算中忽略了白天时段光强变化引起水气压差变化对蒸腾速率的影响,尤其是在强光条件下估算作物实际需水量时会产生较大误差[20]。因此,分别针对玫瑰和辣椒2种作物,Suay等[21-22]考虑了水汽压差对作物蒸腾的影响,建立了温室无土基质栽培作物蒸腾量估算的线性模型。然而,这种线性拟合辐射项与水气压差项的模型准确性是作物特异性的[23],在很大程度取决于温室小气候环境,且在机理特性上未考虑阻抗参数对蒸腾表面水汽交换的影响。
灌溉作为温室基质栽培作物水分供给的唯一来源,由于植物吸收和灌溉送水过程造成的额外损失,灌溉需求量始终大于作物需水量。值得注意的是,由于蒸腾模型中的净辐射经过冠层叶面积指数截留,削弱了室外太阳辐射对冠层以下部分植株水分损失的影响,这就意味着在作物生长初期叶面积指数较小时,冠层截留净辐射远小于温室透光后的太阳辐射。而在依据太阳辐射阈值触发灌溉事件的温室中,在叶面积指数小而太阳辐射强的情况下,仅使用蒸腾模型来估算整株植株需水量会严重低估灌溉需水量。为解决这一问题,本文在忽略土壤蒸发来修正Penman-Monteith模型得到番茄基质栽培蒸腾模型的基础上,将其与太阳辐射线性供水模型结合使用多元线性回归来建立蒸腾-辐射综合灌溉模型,为温室番茄基质栽培的灌溉需水量预测和实施规划奠定基础。
1 温室番茄基质栽培场景介绍
本文的研究对象是位于上海崇明国家设施农业工程技术中心示范基地Venlo温室(31°57'N, 121°70'E)的稻壳炭有机基质栽培的荷兰小番茄(品种为Gorioso)。番茄的种植密度约为每平方米2.5株,在A8温室中共有18个间隔1.5 m的种植列,每个种植列有23个间隔0.3 m的种植单位,每个种植单位有4个基质种植盒(0.15 m×0.15 m×0.1 m),分为2组,每组有2个紧密挨着的种植盒,每个种植盒配有1个滴箭,每2个种植盒中间另有1个滴箭,即每2株植株配有3个滴箭实施灌溉供水。采用辐射阈值触发定量灌溉,即室外太阳辐射每累积100 J/cm2时使每个滴箭出水70 mL,废液回收量由人工记录。营养液的正常供给间隔为7~10 d,而在定植前期番茄根系吸收水分和养分能力较弱,在3~5 d浇施速效氮肥促进番茄植株营养生长。
番茄植株于每年9月20日定植,整个生长全周期约为300 d。移栽定植后平均每天植株增高5 cm,在定植约35~40 d后,番茄植株冠层高达2 m且叶面积指数达到最大值,经叶片修剪会维持在3.3~3.5 m2/m2范围内。从定植到番茄成熟约为80 d,约每天可着色一个果实,平均8~10 d采摘一串果实,然后去掉果实下的2~3片叶片并卷起相应的茎干。每株植株大致保持18个茎节,每3个茎节有一串果实,即每株番茄保持6串果实,每串果实数控制在8个,单个成熟果实质量约为45 g,整个结果期每株番茄的总采摘量约为28~32串。
温室内气象站随着番茄植株生长而受控向上移动,与植株冠层保持0.3 m的间距,可获取室内空气温度、湿度和CO2浓度等数据;室外气象站可获取室外太阳辐射日累积量、降雨量和风速等数据。用于研究方法验证分析的试验数据来源于4 a番茄生长全周期的采集记录,具体时间段为2014-09-20至2015-07-16、2015-09-20至2016-07-15(闰年)、2016-09-20至2017-07-16和2017-09-20至2018-07-16。
2 温室番茄基质栽培灌溉模型的建立
2.1 Penman-Monteith蒸散模型及其局限性
估算作物的灌溉需水量取决于作物蒸散量的计算。依据能量平衡和湍流传导,FAO给出了估算大田作物参考蒸散量的应用指导标准——Penman-Monteith蒸散模型,如式(1)所示。

式中ET0表示作物蒸散的潜热通量,mm/d;=2.45为单位转换系数,MJ/kg;为土壤热通量,MJ/m2·d;R为净辐射,是净短波辐射R和净长波辐射R的差值,MJ/m2·d;e为平均饱和水汽压,e为实际水汽压,kPa;D为水汽压差与温度曲线的斜率;r为空气动力阻抗,s/m;r为表层阻抗,s/m;ρ为恒压下的平均空气密度,kg/m3;c为恒压比热,MJ/kg·℃;=0.054为干湿表常数,kPa·℃。
当日光温室中的空气流通较慢且风速近似为0时,空气动力阻抗r的数值会趋于无限大,导致引入草基准面简化参数得到的固定表层阻抗(70 s/m)和反照率为 0.23的Penman-Monteith模型一般式中水气压差近似为0。因此,为了使其能够应用于不常通风的温室中,研究学者通过修正空气阻抗参数的风速项,得到了适用于温室土培作物的参考蒸散模型,如式(2)所示。并且,在估算实际作物蒸散量时,作物系数K以FAO给定不同生长阶段的推荐值(K在苗期为0.75,在开花期为1.05,在结实期为0.8)来转换参考蒸散量。
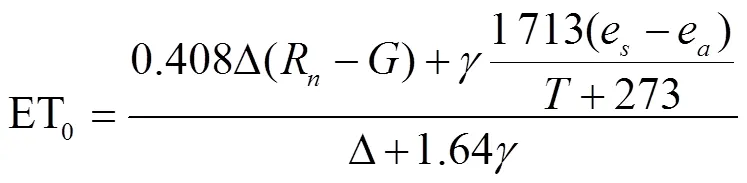
式中为温室空气日平均温度,℃。
对于无土有机基质栽培作物,考虑到温室基质栽培与土壤种植之间作物生长和蒸腾影响因素的明显差异,应考虑从上述蒸散模型中去除土壤蒸发。
2.2 生长参数及阻抗修正
FAO指出,Penman-Monteith模型一般式是监测作物蒸散过程的物理和生理因素的简单表示,主要取决于作物种植环境中的气候条件;其引入作物系数以区分草基准面和实际作物之间的蒸散量,并不涉及作物实时生长,而是从蒸散量的历史测量值与估计值的比值经验得出。因此,本文将固定作物高度的草基准面替换为实际作物冠层蒸腾面,以叶面积指数实际变化来修正阻抗参数经验表示,在无需引入作物系数的情况下直接计算作物蒸腾量。
2.2.1 基于番茄生长模型模拟叶面积指数
叶面积指数(Leaf Area Index,LAI)与实际作物表层阻抗r和到达作物冠层的截留净辐射R相关。对于FAO引入固定作物高度的草基准面,Penman-Monteith模型一般式以经验公式(LAI=24)来获取叶面积指数,忽略了不同生长阶段作物冠层叶片合理的生长变化。考虑引入番茄生长模型(Tomato Growth Model,TOMGRO)来模拟番茄冠层叶面积指数的实际变化,以便更为直观地反映叶面积指数变化对冠层蒸腾的影响。
在实际温室中,当叶面积指数达到允许最大值时会进行叶片修剪使得成熟到采收期间叶面积指数保持不变,这种温室作物栽培特有的种植管理方式使得TOMGRO模型中叶面积指数LAI的增长模式通常以sigmoid函数形式表示[24],如式(3)所示。

式中LAI为初始叶面积指数,m2/m2;LAImax为最大叶面积指数,m2/m2;两者均为实际温室中人工记录。L= 35 000为叶片生长率滞后系数,L=4.1为叶片生长率。为主茎干上的茎节数(节),在该参数的计算表示中,TOMGRO模型假设单个作物生长最适温度,用增减因子来模拟作物生长对室内气温的响应,其计算如下:


式中N=0.5,为最大茎节出现率,f()为温度抑制函数,0()=2.5+0.025,1()=2.5-0.05,分别为作物生长率增长和衰减因子,NUM=1,为最大茎节生长率。
2.2.2 结合太阳辐射和叶面积指数修正表层阻抗
表层阻抗r描述的是水蒸气从作物叶片内部饱和区域或蒸散面以下扩散到蒸散面本身的阻力,在简化得到Penman-Monteith模型一般式时,其经验表示结果(r≈70 s/m)为草基准面的单叶气孔阻抗(r=100 s/m)与有效叶面积指数(LAI=0.5LAI)的比值。
然而,实际温室有机基质栽培的番茄冠层蒸腾表面与大面积覆盖地表的草基准面在蒸腾作用上存在较大差异,这就意味着将表层阻抗当作固定值并不足以反映基质栽培的番茄冠层叶片生长变化对蒸腾作用的影响。因此,考虑将番茄冠层模拟叶面积指数和与太阳辐射密切相关的气孔阻力相结合,以获得合理的表层阻抗计算方法。
对于温室番茄,到达冠层的太阳辐射是影响叶片气孔阻抗的最关键因素。Boulard等[11]给出了与草基准面的经验公式不同的单叶气孔阻抗r计算方式。
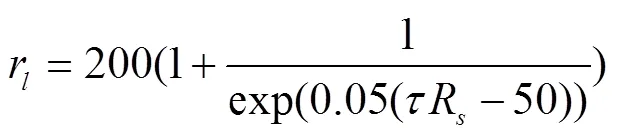
式中R为室外太阳辐射,MJ/m2·d;=0.65为崇明温室透光率。在不考虑修剪叶片影响土壤覆盖度的情况下,草基准面的经验公式中有效叶面积指数由作物生长期的实际模拟叶面积指数表示,与受室外太阳辐射影响的单叶气孔阻抗r相结合,可得到忽略了土壤蒸发影响的基质栽培番茄的新的表层阻抗r计算表示如下:

2.2.3 固定空气动力阻抗
空气动力阻抗r描述的是从零平面位移高度以上的距离向作物的有效表面传递动量的阻力。FAO指出,对于大范围覆盖地表的大田作物,必须考虑零平面位移高度和粗糙度长度等参数,而无土基质栽培环境下的高丛番茄则无需考虑这些与土壤蒸发相关的参数。
引入草基准面后,以2 m高的风速2简化计算空气动力阻抗,从其计算表示(r=208/2)不难发现,当不频繁通风的日光温室中空气流通较为缓慢且风速2≈0时,空气动力阻抗趋于无限大,这使得Penman-Monteith模型中的r/r≈0且水气压差接近于0,从而造成模型缺失作物蒸腾生理特性原理中所涵盖的为从蒸腾表面去除水汽过程的提供动力的水气压差项,模型最终会退化成以太阳辐射为主要影响因素的线性模型。但是,这种由简化计算表示和风速之间的极端关系所产生的一系列问题与实际情况相矛盾,因为实际温室中的番茄作物生长在低风速情况下,仍会与小气候环境之间存在水汽交换和热量传输,并不会出现数学表示直观反映的某项影响因素丢失的情况。
因此,本文在建立温室番茄的基质栽培蒸腾模型过程中,考虑忽略与土壤蒸发相关的土壤覆盖度、零平面位移高度和粗糙度长度的经验计算影响,在封闭温室或空气流速缓慢的温室中将新的空气动力阻抗取为固定值(r=500 s/m),以简化依据湍流传导和热传递原理推导的复杂计算表示。针对上述处理,通过敏感性分析验证了空气动力阻抗在可能取值范围内变化对蒸腾量模拟结果影响较小而作为固定值的合理性,并且在Wang等[25]的相关研究中也明确表示在封闭温室或低气流低风速温室中,将其看作是一个固定值是合理的。
2.3 基质栽培番茄蒸腾模型
经过上述推导和修正过程可以看出,辐射项和对流项仍被保留以反映作物蒸腾生理特性,并且考虑与作物冠层蒸腾表面水汽交换直接相关的阻抗参数的修正计算,根据实际温室有机基质栽培环境来去除土壤蒸发的影响,并以实际作物冠层蒸腾表面的生长变化来修正相关参数的经验计算公式,得到最终适用于温室稻壳炭有机基质栽培场景的番茄蒸腾模型表示如下:

式中Tr为温室基质栽培番茄蒸腾量,mm/d;C=86 400/ 1.01=4.79×105为计算系数, =0.62为水蒸气与干空气的分子量比,0.287为比气体常数;VPD=e−e为水汽压差,kPa;R=R(1exp(·LAI))为番茄冠层截留净辐射,MJ/m2·d;在冠层截留净辐射的计算中,=0.8为番茄消光系数,LAI为TOMGRO模型模拟的冠层叶面积指数;r=500 s/m为新的蒸腾模型中固定空气动力阻抗表示;r为新的蒸腾模型中冠层表层阻抗表示。
2.4 温室基质栽培番茄蒸腾-辐射综合灌溉模型
由番茄基质栽培蒸腾模型辐射项中截留净辐射R的计算表示可知,到达作物冠层经过叶面积指数截留的净辐射远小于温室透光后的太阳辐射,这就削弱了实际太阳辐射对番茄整株植株蒸腾的影响。在移栽定植约35~40 d后,番茄植株冠层高达2 m且叶面积指数维持在3.3~3.5 m2/m2范围内,这就导致温室透光后的太阳辐射在株间分布不均衡,番茄基质栽培蒸腾模型在很大程上用于反映作物冠层蒸腾变化,而不能较为准确地反映整株植株冠层及其以下部分的所有叶片蒸腾耗水情况。
此外,由于温室灌溉方式的不同,在生长初期叶面积指数较小造成截留净辐射较小时,强烈的太阳辐射使得温室灌溉用水供给量要远高于蒸腾模型估算的作物需水量。这主要是因为蒸腾模型中所引入的番茄生长变化仅针对冠层蒸腾表面而言,较小的叶面积指数使得冠层覆盖度小,而冠层以下部分的耗水情况未被蒸腾模型考虑进去。针对上述问题,需要在反映番茄冠层耗水的基质栽培蒸腾模型的基础上,额外考虑太阳辐射对冠层以下部分耗水的影响,建立蒸腾-辐射综合灌溉模型来模拟番茄整株植株的灌溉需水情况。
由于新建立的番茄基质栽培蒸腾模型和太阳辐射线性供水模型的表示形式均已确定,它们在灌溉模型相关研究中均被以经验比例系数线性转换为灌溉量,因而考虑以番茄基质栽培蒸腾模型和太阳辐射线性供水模型作为二元自变量以及模拟灌溉量作为因变量来建立带有偏置项的多元线性回归[26-27]灌溉模型,其表示如下:

式中I为蒸腾-辐射综合灌溉模型模拟灌溉需水量,mm/d;Tr为新建立的番茄基质栽培蒸腾模型模拟冠层蒸腾量,mm/d;I=τR/λ为太阳辐射线性供水模型模拟作物耗水量,mm/d;ω和ω分别表示番茄基质栽培蒸腾模型和太阳辐射线性供水模型的权重系数,ω为偏置项回归系数。
SPSS软件[28]被用于求解温室基质栽培番茄的蒸腾-辐射综合灌溉模型的各个权重系数,以及验证建立蒸腾-辐射综合灌溉模型是否可以满足使用多元线性回归的约束条件,即试验数据样本残差需要服从或近似为正态分布。
为了确保使用多元线性回归建立的温室基质栽培番茄蒸腾-辐射综合灌溉模型具有一定的合理性,并能较好地估算其他年份的灌溉需水量,这就需要足够的数据样本进行训练支撑。基于现有试验数据用于训练和验证的样本量不足,本文在现有数据样本基础上加入适当的高斯噪声来形成新的样本数据,以达到合理扩充样本的目的。
以灌溉用水量数据为例,所加入的扰动表示为0均值且方差为0.3的高斯噪声,以此反映实验温室人工记录排液量来获取实际灌溉消耗所允许的误差值。经过随机高斯采样对现有数据产生干扰后,用于多元线性回归的样本数据量从4个全周期数据扩展了一倍。以6个番茄生长全周期(2014-09-20至2015-07-16和2015-09-20至2016-07-15的原始数据以及在4个全周期原始数据基础上加入高斯噪声后的新数据)作为训练样本,可得到蒸腾-辐射综合灌溉模型的回归系数;并以余下2个全周期(2016-09-20至2017-07-16和2017-09-20至2018-07-16)的原始数据作为测试样本,来验证多元线性回归模型估算温室稻壳炭基质栽培的番茄灌溉量的准确性。
在试验样本数据扩充后,使用SPSS软件对训练输入(番茄基质栽培蒸腾模型模拟冠层蒸腾量和太阳辐射线性供水模型模拟作物耗水量)和训练输出(实际灌溉量)进行线性回归拟合,可得到蒸腾-辐射综合灌溉模型的回归系数分别为ω=0.412,ω=0.706,ω=0.338,则温室稻壳炭基质栽培番茄的蒸腾-辐射综合灌溉模型的具体表示如下:
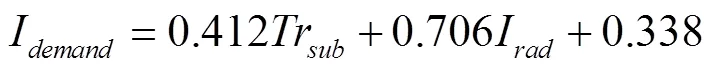
此外,SPSS软件线性拟合得到的蒸腾-辐射综合灌溉模型的拟合精度为R2=0.916;并且它还验证了使用多元线性回归建立灌溉模型是否满足约束条件——样本残差近似或服从正态分布,样本标准残差的直方图和正态P-P图验证结果如图1所示。
由图1可知,样本标准残差的均值(Mean=6.94E-15)近似为0,且在直方图分布形式上近似为0均值且方差(Std.Dev.=0.999)约为1的正态分布,而正态P-P图也显示了偏差的期望累积概率和与预测累积概率大致相同,大部分点都位于直线上,只有少部分点小距离偏离直线。这进一步验证了用于求解模型拟合系数的试验数据在引入高斯噪声扩充样本的情况下,数据量达到了偏差服从正态分布的约束条件,这表明使用多元线性回归建立的蒸腾-辐射综合灌溉模型具有一定的合理性。
3 结果与分析
3.1 温室土培蒸散模型与基质栽培蒸腾模型的对比
为了验证本文所建立的温室番茄基质栽培蒸腾模型修正的合理性,以及相比于温室土培作物蒸散模型应用于无土基质栽培场景估算的准确性,对比分析了这2种模型模拟番茄蒸腾量的结果,如图2所示。验证分析所使用的试验数据来源于上海崇明A8温室4 a采集记录的番茄生长全周期300 d的气象数据、TOMGRO模型模拟生长参数数据和实际蒸腾量数据;4个番茄生长全周期的具体时间段为2014-09-20至2015-07-16、2015-09-20至2016-07-15(闰年)、2016-09-20至2017-07-16和2017-09-20至2018-07-16。
从图2中可以看出,土培作物蒸散模型与番茄基质栽培蒸腾模型模拟蒸腾量的变化趋势非常接近,主要是因为2个模型都是在Penman-Monteith蒸散模型原始表示的基础上依据栽培环境进行简化和修正的;而且作为影响番茄蒸腾能量供应的主要因素,室外太阳辐射使得2种模型的模拟蒸腾量与净辐射之间高度相关。依据温室土壤栽培环境修正的Penman-Monteith蒸散模型高估了作物蒸腾量,而且比番茄基质栽培蒸腾模型具有更高的模拟值,这与其他研究学者[29-31]的试验结果相似。造成这种现象的原因是由于Penman-Monteith蒸散模型主要是针对室外大田种植环境下农作物的灌溉指导而建立的,而露天土壤种植作物受到土壤蒸发和额外降水供应对蒸散过程的影响,基质栽培和土壤种植之间的生长环境和水分补给方式的差异导致了作物蒸腾量的估算存在明显差距。
此外,由图2中显示的模拟番茄蒸腾量的对比可以看出,约25 d内的模型模拟蒸腾量与实际蒸腾量的数值较为相近,这主要是由于生长初期番茄冠层叶面积指数较小,在净辐射的计算中对温室透光后的太阳辐射截留的影响较小,使得作物蒸腾量的模拟值与实际值都很小,这符合叶面积指数与潜在蒸腾速率成正比相关的认知。而在番茄摘果后期(距离摘果结束约50~60 d),由于每次摘果后修剪该果实下2~3片叶片和干枯茎干,直至一株番茄6串果实完全采摘,该阶段的叶面积指数会相应减小,这与生长初期叶面积指数较小的情况类似,会使得2种模型模拟蒸腾量的结果与实际蒸腾量均较为相近,模拟误差相对变小。
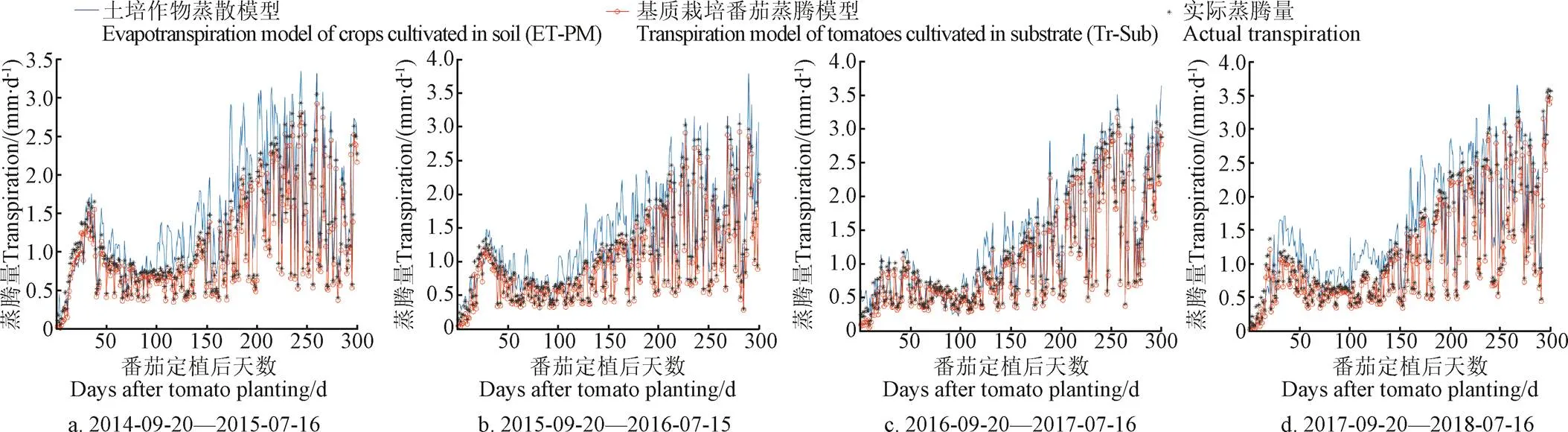
图2 2种模型(ET-PM和Tr-Sub)在不同年份的模拟蒸腾量与实际蒸腾量的比较
表1给出了温室土培作物蒸散模型和番茄基质栽培蒸腾模型模拟蒸腾量分别与实际蒸腾量之间的相关系数和误差结果,由表1可以看出,与土培作物蒸散模型相比,番茄基质栽培蒸腾模型模拟番茄蒸腾量具有更小的误差,其与实际蒸腾量的相关系数大于0.95,平均绝对误差小于0.1 mm/d、平均相对误差小于10%,均方根误差小于0.1,且均比土培作物蒸散模型的模拟误差小2倍以上。这表明,在估算温室稻壳炭有机基质栽培的番茄蒸腾量时,依据Penman-Monteith蒸散模型修正得到的番茄基质栽培蒸腾模型具有较高的准确性。并且,相比于土培作物蒸散模型以经验作物系数来反映草基准面和实际作物蒸散面之间的生长差异,番茄基质栽培蒸腾模型通过模拟番茄冠层蒸腾表面的叶片实时生长变化,在保留了作物蒸腾机理特性的同时,从蒸散中消除土壤蒸发的影响以此来反映在土培作物蒸散和基质栽培作物蒸腾之间的差异是合理的。
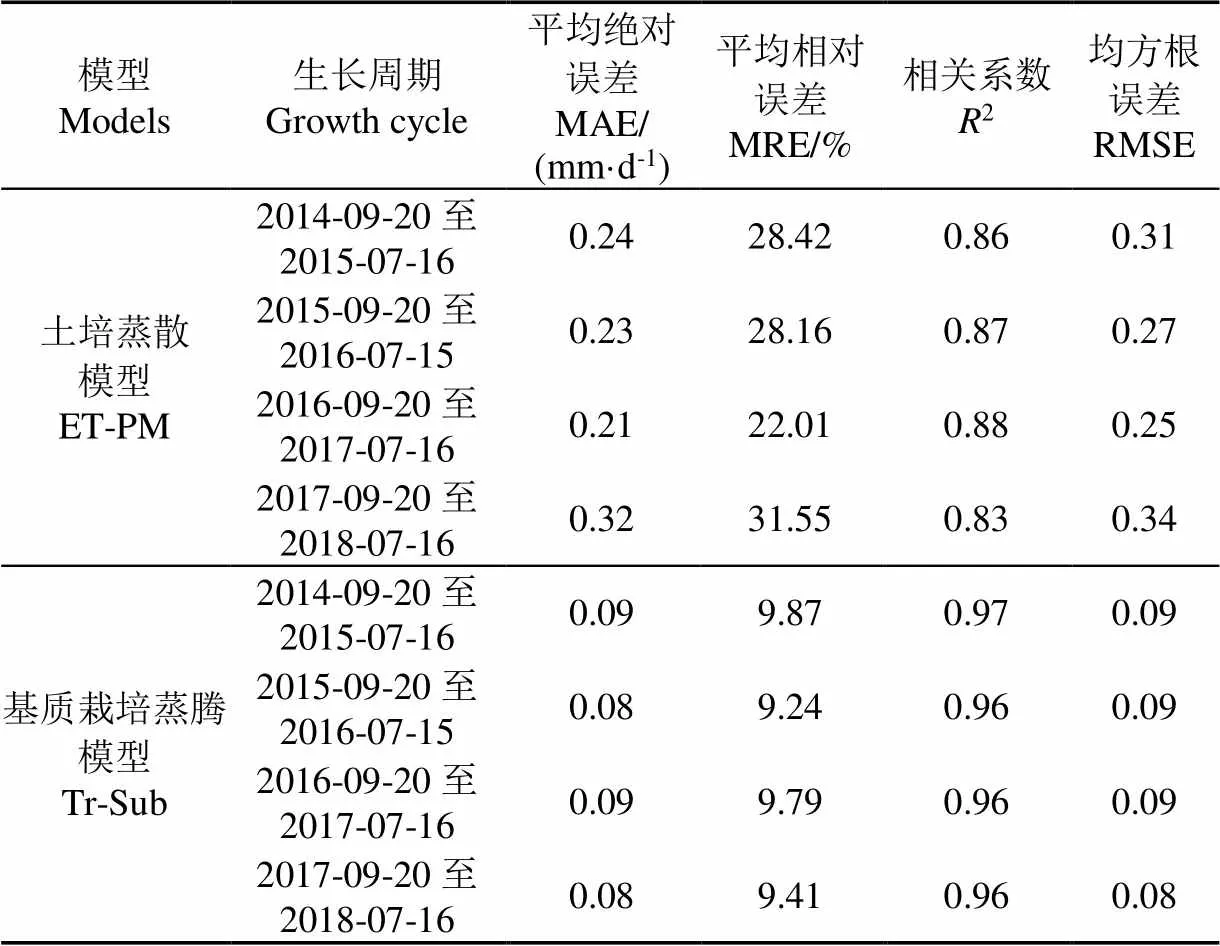
表1 2种模型模拟蒸腾量与实际蒸腾量的相关系数和误差结果
3.2 蒸腾-辐射综合灌溉模型模拟灌溉量与实际灌溉量的对比分析
为了进一步验证使用多元线性回归方法建立的蒸腾-辐射综合灌溉模型模拟番茄灌溉需水量的准确性,使用保留的2个全周期内(2016-09-20至2015-07-16和2017-09-20至2018-07-16)的原始数据对灌溉模型进行了仿真模拟,并与实际灌溉量数据进行对比分析,如图3所示。

图3 蒸腾-辐射综合灌溉模型模拟灌溉量与实际灌溉量的比较
由于番茄生长初期叶面积指数较小,蒸腾模型中参与计算的冠层截留净辐射严重削弱了室外太阳辐射的实际影响,使得估算蒸腾量较小;而从图3中可以看出,番茄生长初期的灌溉量并非与蒸腾量直接相关,这也就意味着在基质栽培番茄蒸腾模型的基础上附加考虑室外太阳辐射的影响来建立综合灌溉模型是有必要的。一方面,基质栽培番茄蒸腾模型考虑了温室小气候环境变化、作物实时生长变化和冠层截留净辐射对作物耗水的综合影响;另一方面,额外考虑室外太阳辐射的影响,以补偿其在不同生长阶段冠层叶片截留下受到不同程度的削弱,两者结合则能更好地反映影响番茄耗水的因素。
由图3可知,2017-09-20至2018-07-16的生长全周期内出现了少数情况(2017-09-24、2017-10-01、2017-10-02和2018-06-25)的模拟灌溉量与实际灌溉量之间存在较大偏差,而绝大部分模拟灌溉需水量变化趋势上与实际灌溉量基本一致且数值上的误差较小。此外,图3中显示在灌溉量数值较大时,模拟值与实际值较为相近;而在灌溉量数值较小时,两者存在一定误差,这主要是因为灌溉模型在蒸腾项和辐射项的数值较小时,偏差项的存在对模型估算结果产生了主导影响。为了更精确地分析蒸腾-辐射灌溉模型估算有机基质栽培番茄的灌溉需水量的总体准确性,表2给出了蒸腾-辐射综合灌溉模型模拟灌溉量与实际灌溉量之间的误差结果以及相关系数。
由表2可以看出,2个生长全周期的模拟灌溉量与实际灌溉量之间的平均相对误差均小于20%,平均绝对误差均小于0.29 mm/d且均方根误差也都不大于0.3,这些误差数值也都小于扩充试验数据所使用的高斯噪声的方差,也就是崇明温室人工记录灌溉相关数据所允许的误差0.3 mm/d,且表中的决定系数均高于0.95。因此,基于多元线性回归建立蒸腾-辐射综合灌溉模型具有一定的合理性,且能够较为准确地预测温室稻壳炭有机基质栽培番茄的灌溉需水量。

表2 实际灌溉量与模拟灌溉量的决定系数和误差分析结果
3.3 叶面积指数与空气动力阻抗的敏感性分析
模型参数的敏感性分析是指其他参数保持不变的情况下,模型中某一参数的变化幅度对结果的影响程度。为了验证有机基质栽培番茄蒸腾模型中将空气动力阻抗r作为固定值的合理性,以及考虑冠层蒸发面叶面积指数LAI反映实际作物生长情况的必要性,针对这2个参数使用SimLab软件中FAST方法作了局部敏感性分析,分析参数r在对数函数表示中风速2变化范围0.05~1.0 m/s内其值变化以及参数LAI在LAI~LAImax范围变化分别对测试期间有机基质栽培番茄蒸腾模型模拟值的影响。
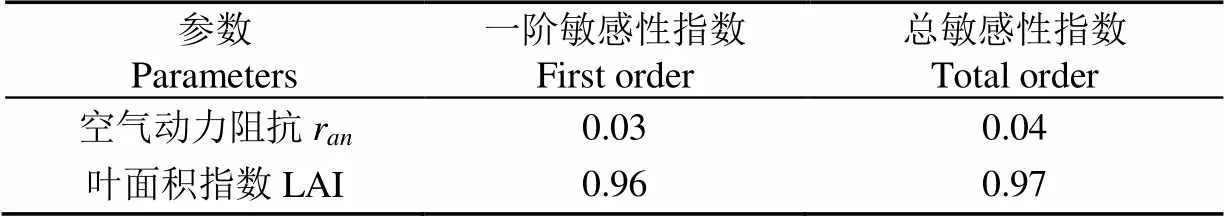
表3 叶面积指数与空气动力阻抗的敏感度
一阶敏感性指数反映的是参数对模型输出总方差的直接贡献率,总敏感性指数反映了参数直接贡献率和通过与其他参数的交互耦合作用间接对模型输出总方差的贡献之和。
从表3中可以看出,参数r的一阶敏感性指数和总敏感性指数均小于0.1,这表明该参数对模拟结果影响较小,将其作为固定值较为合理;而参数LAI的敏感性指数远大于0.1,这表明该参数对模拟结果影响非常大,其主要原因是蒸腾模型中考虑室外太阳辐射到达作物冠层的截留净辐射与叶面积指数紧密相关,因而根据作物冠层蒸腾面的实际生长情况模拟叶面积指数是有必要的。
4 结 论
为解决温室土培作物蒸散模型不能直接适用于温室稻壳炭基质栽培番茄灌溉的问题,本研究基于作物蒸散机理特性去除土壤蒸发的影响,首先建立了温室基质栽培番茄蒸腾模型。选取了上海崇明A8温室番茄4个生长全周期采集记录的试验数据进行仿真,由仿真结果可以看出,温室土培作物蒸散模型过高估计番茄蒸腾量,而温室基质栽培番茄蒸腾模型模拟番茄蒸腾量具有更小的误差,其与实际蒸腾量的平均绝对误差小于0.1 mm/d、平均相对误差小于10%和均方根误差小于0.1,且均比温室土培作物蒸散模型的模拟误差小2倍以上,因而更适合于不考虑土壤蒸发的基质栽培番茄蒸腾量估算。
然后,考虑到基质栽培番茄蒸腾模型中净辐射经过冠层叶片截留后削弱了室外太阳辐射对冠层及以下部分整株植株的耗水影响,本研究在基质栽培番茄蒸腾模型的基础上额外考虑了太阳辐射对整个番茄植株耗水量的影响,并使用多元线性回归建立了蒸腾-辐射综合灌溉模型。以番茄2个生长全周期内(2016-09-20至2015-07-16和2017-09-20至2018-07-16)的试验数据对蒸腾-辐射综合灌溉模型进行模拟精度测试,由仿真结果可以看出,模拟灌溉量与实际灌溉量之间的平均相对误差小于20%,平均绝对误差小于0.29 mm/d且均方根误差不大于0.3,这些误差数值也都小于崇明温室人工记录灌溉相关数据所允许的误差。因此,蒸腾-辐射综合灌溉模型在整个生长期内对番茄灌溉需水量的估算具有较高精度,且模型参数的获取和仿真估算过程相对简单。
综上所述,蒸腾-辐射综合灌溉模型在反映番茄耗水情况受到温室小气候环境、室外太阳辐射和作物生长变化的综合影响上具有一定的优势,可以作为有效估算温室稻壳炭有机基质栽培番茄的灌溉消耗的一种方法,并能为温室精准灌溉控制提供模型支撑。
[1]Fernqvist F. Consumer Experiences of Tomato Quality and the Effects of Credence[D]. Uppsala: Swedish University of Agricultural Sciences, 2014.
[2]孙婧,田永强,高丽红,等. 秸秆生物反应堆与菌肥对温室番茄土壤微环境的影响[J]. 农业工程学报,2014,30(6):153-164.
Sun Jing, Tian Yongqiang, Gao Lihong, et al. Effects of straw biological reactor and microbial agents on physicochemical properties and microbial diversity of tomato soil in solar greenhouse[J]. Transactions of the Chinese Society of Agricultural Engineering (Transactions of the CSAE), 2014, 30(6): 153-164. (in Chinese with English abstract)
[3]Nam D S, Moon T, Lee J W, et al. Estimating transpiration rates of hydroponically-grown paprika via an artificial neural network using aerial and root-zone environments and growth factors in greenhouses[J]. Horticulture, Environment and Biotechnology, 2019, 60: 913-923.
[4]Sezen S M, Yazar A, Tekin S. Physiological response of red pepper to different irrigation regimes under drip irrigation in the Mediterranean region of Turkey[J]. Scientia Horticulturae, 2019, 245: 280-288.
[5]Barrett G E, Alexander P D, Robinson J S, et al. Achieving environmentally sustainable growing media for soilless plant cultivation systems: A review[J]. Scientia Horticulturae, 2016, 212: 220-234.
[6]Shiri J, Landeras G, Nazemi A H, et al. Daily reference evapotranspiration modeling by using genetic programming approach in the Basque Country (Northern Spain)[J]. Journal of Hydrology, 2012, 414(2): 302-316.
[7]Allen R G, Pereira L S, Raes D, et al. Crop evapotranspiration: Guidelines for computing crop water requirements[M]. Rome: Irrigation and Drainage Paper No 56, 1998.
[8]张培新,贺超兴,张志斌,等. 基于生理发育时间的日光温室番茄发育模拟模型[J]. 中国农业气象,2006,27(4):314-317.
Zhang Peixin, He Chaoxing, Zhang Zhibin, et al. A simulation model for tomato plant growth and development in solar greenhouse based on physiological development time[J]. Chinese Journal of Agrometeorology, 2006, 27(4): 314-317. (in Chinese with English abstract)
[9]王健,蔡焕杰,李红星,等. 日光温室作物蒸发蒸腾量的计算方法研究及其评价[J]. 灌溉排水学报,2006,25(6):11-14.
Wang Jian, Cai Huanjie, Li Hongxing, et al. Study and evaluation of the calculation methods of reference crop evapotranspiration in solar-heated greenhouse[J]. Journal of Irrigation and Drainage, 2006, 25(6): 11-14. (in Chinese with English abstract)
[10]刘浩,段爱旺,孙景生,等. 基于Penman-Monteith方程的日光温室番茄蒸腾量估算模型[J]. 农业工程学报,2011,27(9):208-213.
Liu Hao, Duan Aiwang, Sun Jingsheng, et al. Estimating model of transpiration for greenhouse tomato based on Penman-Monteith equation[J]. Transactions of the Chinese Society of Agricultural Engineering (Transactions of the CSAE), 2011, 27(9): 208-213. (in Chinese with English abstract)
[11]Boulard T, Wang S. Greenhouse crop transpiration simulation from external climate conditions[J]. Agricultural and Forest Meteorology, 2000, 100(1): 0-34.
[12]Castilla N, Hernández J. The plastic greenhouse industry of Spain[J]. Chron Hort, 2005, 45(3): 15-20.
[13]Gallardo M, Thompson R B, Rodríguez J S, et al. Simulation of transpiration, drainage, N uptake, nitrate leaching, and N uptake concentration in tomato grown in open substrate[J]. Agricultural Water Management, 2009, 96(12): 1773-1784.
[14]Shin J H, Park J S, Son J E. Estimating the actual transpiration rate with compensated levels of accumulated radiation for the efficient irrigation of soilless cultures of paprika plants[J]. Agricultural Water Management, 2014, 135(2): 9-18.
[15]Tai N H, Park J S, Shin J H, et al. Analyses of transpiration and growth of paprika (L.) as affected by moisture content of growing medium in rockwool culture[J]. Korean Journal of Horticultural Science and Technology, 2014, 32(3): 340-345.
[16]Salcedo G A, Reca J, Pérez-Sáiz M, et al. Irrigation water consumption modelling of a soilless cucumber crop under specific greenhouse conditions in a humid tropical climate[J]. Ciência Rural, 2017, 47(2): 1-10.
[17]Nikolaou G, Neocleous D, Katsoulas N, et al. Modelling transpiration of soilless greenhouse cucumber and its relationship with leaf temperature in a mediterranean climate[J]. Emirates Journal of Food and Agriculture, 2017, 29(12): 911-920.
[18]Rouphael Y, Colla G. Modelling the transpiration of a greenhouse zucchini crop grown under a Mediterranean climate using the Penman-Monteith equation and its simplified version[J]. Crop & Pasture Science, 2004, 55(9): 931-937.
[19]Baille M, Baille A, Laury J C. A simplified model for predicting evapotranspiration rate of nine ornamental species vs. climate factors and leaf area [J]. Scientia Horticulturae (Amsterdam), 1994, 59(3/4): 217-232.
[20]Shin J H, Son J E. Application of a modified irrigation method using compensated radiation integral, substrate moisture content, and electrical conductivity for soilless cultures of paprika[J]. Scientia Horticulturae, 2016, 198: 170-175.
[21]Suay R, Martinez P F, Roca D, et al. Measurement and estimation of transpiration of a soilless rose crop and application to irrigation management[J]. Inverse Problems, 2003, 614: 409-433.
[22]Ta T H, Shin J H, Ahn T I, et al. Modeling of transpiration of paprika (L.) plants based on radiation and leaf area index in soilless culture[J]. Horticulture Environment and Biotechnology, 2011, 52(3): 265-269.
[23]Fazlil-Ilahil W F. Evapotranspiration Models in Greenhouse[D]. Wageningen: Wageningen Agricultural University, 2009.
[24]Bacci L, Battista P, Rapi B. Evaluation and adaptation of TOMGRO model to Italian tomato protected crops[J]. New Zealand Journal of Crop & Horticultural Science, 2012, 40(2): 115-126.
[25]Wang S, Boulard T, Haxaire R. Air speed profiles in a naturally ventilated greenhouse with a tomato crop[J]. Agricultural and Forest Meteorology, 1999, 96(4): 181-188.
[26]Bianco V, Manca O, Nardini S. Electricity consumption forecasting in Italy using linear regression models[J]. Energy, 2009, 34(9): 1413-1421.
[27]Hu T, Wang T, Li J, et al. Binary linear regression and statistical learning of behavior model for human teleoperator [C]//IEEE International Conference on Mechatronics & Automation. IEEE, 2016.
[28]Waghmare S N, Sakhale C N. Formulation of experimental data based model using SPSS (Linear Regression) for stirrup making operation by human powered flywheel motor[J]. International Research Journal of Engineering and Technology (IRJET), 2015, 4(2): 461-468.
[29]Prenger J J, Fynn R P, Hansen R C. A comparison of four evapotranspiration models in a greenhouse environment[J]. Transactions of theAmerican Society of Agricultural Engineers, 2002, 45(6): 1779-1788.
[30]Villarreal-Guerrero F, Kacira M, Fitz-Rodríguez E, et al. Comparison of three evapotranspiration models for a greenhouse cooling strategy with natural ventilation and variable high pressure fogging[J]. Scientia Horticulturae, 2012, 134: 210-221.
[31]López-Cruz I L, Olivera-López M, Herrera-Ruiz G. Simulation of greenhouse tomato crop transpiration by two theoretical models[J]. Acta Horticulturae, 2008, 797: 145-150.
Irrigation models for the tomatoes cultivated in organic substrate based on greenhouse environment and crop growth
Xu Lihong, Xiao Kangjun, Wei Ruihua
(201804,)
Water supply is an essential to the normal growth of plants, due to too much or insufficient water can seriously confine the yield and quality of greenhouse crops. The traditional greenhouse irrigation method mainly triggers quantitative irrigation system through timing control and radiation thresholds, without considering the comprehensive impact of microclimate changes on water consumption of crops in the greenhouse environment. In the conventional model, such as converting crop transpiration to irrigation water demand with the empirical ratio factor, there is a lacking the fact that the growth of crop canopy leaves can weaken the effect of solar radiation on water consumption of plants below the canopy. The conventional water supply methods cannot meet the actual needs of crops, and thereby inevitably cause serious waste of water resources and environmental pollution by excessive discharge of nutrient solution waste liquid, eventually deteriorate the economic benefit of crop production in greenhouse. Therefore, it is of great practical value to establish an irrigation model based on crop transpiration physiological characteristics and intercepted radiation effects of actual canopy growth changes. The relevant research of the soilless substrate cultivation crop transpiration model pointed out that the surface evaporation of the substrate can be small or even negligible, indicating that the soil evaporation does not need to be considered when estimating the tomato transpiration of the soilless substrate cultivation. Penman-Monteith evapotranspiration model including soil evaporation cannot accurately estimate the transpiration of substrate-cultivated tomato in terms of greenhouse. In addition, some models were introduced a grass datum with a fixed crop height and a large area covering the ground, in order to empirically simplify the calculation of impedance parameters, and then used historical data to fit crop coefficient as a medium for converting reference evapotranspiration to actual evapotranspiration. However, the estimation process did not consider the actual growth of the crop evapotranspiration surface. In this study, a new transpiration model was established for the tomato plants cultivated on rice-husk charcoal substrate in the greenhouse. The calculation of empirical parameters was modified based on soil evaporation in the original representation of the evapotranspiration model, according to the distinction between soil-cultivated crop evapotranspiration and substrate-cultivated crop transpiration. TOMGRO model was introduced to simulate real-time growth changes of tomatoes to replace the grass datum of fixed growth parameters. The new transpiration model has added the resistance parameters of the water vapor exchange between the leaves and outside air, while maintaining the characteristics of crop transpiration, indicating more suitable for the greenhouse microclimate environment and the real-time growth of crops. In the simulated transpiration results of historical data from each full cycle of tomato growth in four years, compared with the greenhouse soil-cultivated crop evapotranspiration model, the transpiration model of substrate tomato shows a small error to simulate tomato transpiration, where the average absolute error are less than 0.1 mm/d, the average relative error are less than 10%, the root mean square error are less than 0.1 and the correlation coefficient are greater than 0.95. Considering that due to the net radiation interception weakening solar radiation, the transpiration model of substrate tomato can be used to describe the transpiration changes of crop canopy, but ignore the water consumption of the entire plant canopy and below. Therefore, a transpiration-radiation integrated irrigation model was established by multiple linear regression method, where introducing the solar radiation linear water supply model as a linear compensation to estimate the tomato irrigation water requirement using the substrate tomato transpiration model. Comparing the actual irrigation data, the average absolute error of the simulated irrigation volume are less than 0.29 mm/d, the average relative error are less than 20%, and the root mean square error are no more than 0.3. These errors are also less than the allowable error 0.3 mm/d of manual recording irrigation data in the experimental greenhouse. The estimation of greenhouse tomato water consumption in substrate cultivation scenario, transpiration-radiation integrated irrigation model have high accuracy, certain rationality and practicability. These models can provide a theoretical basis for the prediction of irrigation water consumption of rice-husk charcoal substrate cultivated tomato in the greenhouse and irrigation implementation plan.
greenhouse; irrigation; tomato; substrate cultivation; Penman-Monteith model; crop transpiration
徐立鸿,肖康俊,蔚瑞华. 基于温室环境和作物生长的番茄基质栽培灌溉模型[J]. 农业工程学报,2020,36(10):189-196.doi:10.11975/j.issn.1002-6819.2020.10.023 http://www.tcsae.org
Xu Lihong, Xiao Kangjun, Wei Ruihua. Irrigation models for the tomatoes cultivated in organic substrate based on greenhouse environment and crop growth[J]. Transactions of the Chinese Society of Agricultural Engineering (Transactions of the CSAE), 2020, 36(10): 189-196. (in Chinese with English abstract) doi:10.11975/j.issn.1002-6819.2020.10.023 http://www.tcsae.org
2020-04-13
2020-05-10
上海市科技兴农重点攻关项目(沪农科创字(2018)第3-2号);上海市科委创新行动计划项目(17391900900)
徐立鸿,教授,博士生导师,主要从事温室环境建模与控制、预测控制和智能控制研究。Email:xulihong@tongji.edu.cn
10.11975/j.issn.1002-6819.2020.10.023
S271
A
1002-6819(2020)-10-0189-08
