降香黄檀心材和边材内生真菌多样性*
2020-06-02刘昌霖周国英刘君昂
刘昌霖 周国英 肖 柏 刘君昂
(南方人工林病虫害防治国家林业和草原局重点实验室 森林有害生物防控湖南省重点实验室经济林培育与保护教育部重点实验室 中南林业科技大学 长沙 410004)
降香黄檀(Dalbergiaodorifera)俗称香红木、黄花梨,属豆科(Leguminosae)蝶形花亚科(Papilionoideae)黄檀属(Dalbergia)半落叶乔木,是国家二级保护植物(吴国欣,2009)、海南省特有珍贵红木树种。降香黄檀木材质地坚硬沉重,纹理细密,花纹美观,永不开裂、变形,坚固耐腐,是高档家具上等用材;木材还含有芳香油,含芳香物高的心材可入药代替降香,以心材蒸馏得到的降香油,气味清香,久不挥发,是香料的定香剂,也是高级镇痛药材,具有抗血凝、氧化、扩冠脉等作用。降香黄檀这些重要的经济价值和药用价值,导致其长期遭受盗伐和过度砍伐,天然林资源现已残存不多,生产力严重下降(Taoetal.,2010)。近年来,在我国南方一些气候适宜的地区如广东、广西、云南、福建等,正在不断尝试营造降香黄檀人工林,对其苗木栽培技术进行了大量研究(邱治军等,2004;何明军等,2008;李太钊,2017;高媛,2017)。降香黄檀纯林抗病虫害、寒害、风害等自然灾害的能力极弱(李铁牛等,2001),人工栽培过程中在树种搭配、良种选育和人工促进心材形成等方面存在一些亟待解决的科学问题(Yuetal.,2007;伍庆均,2014;黄星,2012)。
心材是由边材通过一系列复杂的生理生化过程转化而来的(Chaffey, 2002),除了受本身遗传信息调控外,还受外界环境的影响,特别是微生物的干扰(崔之益等,2016)。有些植物心材是由单一类型的微生物入侵而产生的,而有些植物心材则是几种微生物共同入侵作用的结果(Sorzetal.,2008)。植物内生菌是指在某一阶段或整个生命周期生活在健康生长的植物组织或细胞内,并对寄主植物不会引起明显病害症状的一类微生物群(Saikkonenetal., 2004)。外界微生物的入侵也能成为降香黄檀的内生真菌,降香黄檀具有丰富的内生真菌。同时,外源性真菌能够有效促进降香黄檀木材心材的形成,而降香黄檀内生菌对心材形成的影响却少有报道。如Sun等(2015)从降香黄檀受伤和健康的茎中分离出多种内生真菌,在木材导管内观察到了菌丝定殖;周双清等(2014)利用从已结心材的降香黄檀上钻取的心材粉末内生菌系培养液,且促进降香黄檀心材的形成。
鉴于此,本研究根据试验样地实际情况,选择树龄相同、立地条件基本一致的降香黄檀,采用HiSeq高通量测序技术,结合木材理化性质,对其心材和边材内生真菌的群落结构和多样性进行研究,并分析其与降香黄檀内环境因子的关系,以期为降香黄檀内生真菌评价和资源利用提供依据,为指导降香黄檀人工促进心材形成提供参考。
1 材料与方法
1.1 试验材料
样品采样地位于海南省琼海市市郊。因样品特殊性和国家法律法规等原因,选择立地条件基本一致[土壤类型为砂红壤,主要混交树种有小叶榄仁(Terminalianeotaliala)、母生(Homaliumhainanense)、沉香(Aquilariasinensis)等]、树龄相同(均为15年)、胸径相近、自然形成心材的降香黄檀(树干25 cm以上部分被砍伐5天左右,留下部位长有嫩叶的枝条)。去除顶部5 cm木材组织,在每株降香黄檀地上15 cm处取树干(降香黄檀心材和边材以下分别简写为JXHW和JXSW)。
在无菌条件下,将木材切成约1 cm×0.5 cm×0.5 cm大小。取5 g木片在0.9%NaCl中摇瓶培养2 h(25 ℃),释放内生菌。用0.22 μm滤膜真空抽滤,所得滤膜-20 ℃储存备用,其余样品储存在-80 ℃低温冰箱中(林文雄等,2009)。
1.2 试验方法
1.2.1 木材含水率测定 参考GB/T1931—2009,将试样置于(103±2)℃烘箱中干燥,根据下式计算木材含水率:木材含水率(%)=(W0-W1)/W1×100,式中W0和W1分别为干燥前、后试样质量。
1.2.2 pH测定 将试样破碎后置于通风良好、无酸碱气体的室内气干,均匀混合后取约200 g,用植物原料粉碎机全部制成通过40目筛的木粉,置于广口瓶中备用。称取木粉3 g(精确至0.01 g),置于50 mL烧杯内,加入新煮沸并冷却至室温的蒸馏水30 mL,搅拌5 min,放置15 min后再搅拌5 min,静置5 min测定pH,精确至0.02。
1.2.3 灰分含量测定 精确称取3 g木粉,置于预先灼烧的坩埚中,先将坩埚放在电炉上烧灼使木粉炭化:然后将坩埚置于高温炉中,在(575±25)℃下灼烧至恒质量为止(无黑色炭素)。灰分含量计算公式如下:
(1)
式中:G0为灰渣质量(g);G1为干试样质量(g);W为试样含水率(%)。
1.2.4 热水抽提物含量测定 精确称取2 g样品,另外称取1份测定试样含水率;将试样小心移入250 mL锥形瓶中,加入200 mL 95~100 ℃的蒸馏水进行热水提取,将装有物料的滤器放入烘箱中,在(105±3)℃下干燥至恒质量。热水抽提物含量计算公式如下:
(2)
式中:G0为试样抽提前绝干质量(g);M1为试样抽提后绝干质量(g)。
1.2.5 矿质元素含量测定 采用硝酸-双氧水消解,ICP-aes 测定钾、钙、镁含量(倪张林等,2013)。称取样品0.2 g(精确至0.000 1 g),置于聚四氟乙烯消解罐中,加入5 mL浓硝酸,2 mL 30%过氧化氢,微波消解 20 min,待溶液澄清后,定容至50 mL,过滤,上机测定各元素含量。
1.2.6 DNA的提取 使用总DNA 提取试剂盒[FastDNA SPIN kit for Soil(Mpbio, USA)],具体方法参照说明书。
1.2.7 DNA的检测 采用1.5%琼脂糖凝胶电泳和 Nanodrop2000 分光光度计检测 DNA 的纯度和浓度。
1.2.8 真菌的特异性扩增 PCR反应体系(25 μL):DNA模板1 μL,引物ITS2F(5′-GCATCG ATGAACGCAGC-3′)、ITS2R(5′-TCCTCCGCTTATTG ATATGC-3′)各1 μL,5X TransStart Fastpfu Buffer 5 μL,2.5 mmol·L-1dNTPs 2 μL,FastPfu DNA Polymerase 0.5 μL,dd H2O 14.5 μL。PCR反应条件:1×(95℃,2 min);30×(95 ℃,20 s;55 ℃,30 s;72 ℃,30 s;72 ℃,5 min);4 ℃ 保存。每种样品取3 μL进行检测(1.5%琼脂条凝胶电泳),3次重复。将同一样品的 PCR 产物混合后用2%琼脂糖凝胶电泳检测,使用 AxyPrepDNA 凝胶回收试剂盒(AXYGEN 公司)切胶,回收 PCR 产物,Tris-HCl 洗脱;2%琼脂糖电泳检测。
1.2.9 HiSeq文库构建及测序 使用BIOO SCIENTIFIC公司的NEXTflexTMRapid DNA-Seq Kit for Illumina 建库试剂盒进行文库构建,构建好的文库经过Qubit3.0和QPCR定量和文库检测。高通量测序试验由美因健康科技(北京)有限公司完成。
1.3 数据处理及分析
1.3.1 数据的过滤、拼接和质控 使用FLASH软件将Read1和Read2的overlap进行拼接, 拼接后的序列为Raw Tags。拼接过程要求重叠区的最小长度为10 bp, 最大错配率为10%。使用Qiime(Caporasoetal.,2010)对Raw Tags进行质控, 截断含5个以上连续N或低质量碱基的Tags, 进一步过滤掉连续高质量碱基长度小于Tags长度75%的Tags, 从而获得Clean Tags。嵌合体的检定采用UCHIME算法(Edgaretal.,2011),与Unite数据库比对,嵌合体序列去除后即获得作为后续数据分析的Effective Tags。
采用R软件对初始结果进行相对丰度制图,利用SPSS 19.0进行方差分析(Duncan)及相关性分析(Pearson),利用Canoco for Windows 4.5进行冗余分析。
1.3.2 OTU聚类分析 通过归类操作,根据各个序列间的相似度分组,将1个小组认定为1个OTU(operational taxonomic units)。本试验在97%相似水平下对OTU进行归类和生物信息统计分析。
采用 RDP classifier 贝叶斯算法对97%相似水平的OTU代表序列进行分类学分析,选用的比对数据库为Unite数据库。
2 结果与分析
2.1 降香黄檀心材和边材内生真菌群落结构和多样性
2.1.1 降香黄檀心材和边材内生真菌数据筛选 6个样本分别测定,总获得2 193 336条序列。经过过滤嵌合体后, 用于后续分析的序列共1 970 694条,碱基总数为761 438 850 bp,有效序列平均长度为385 bp。碱基质量值Q30[Effective Tags中碱基质量值大于30(测序错误率小于0.1%)的碱基所占百分比]比例大于93.84%,表明样本数据可靠。测序数据质控结果统计如表1所示。

表1 测序质量信息汇总①
①Clean Tags:Tags过滤低质量和短长度Tags后的序列Tags filter the sequence after low quality and short length Tags;Q20content(%):测序错误率低于1%的碱基序列所占百分比 Percentage of base sequence with sequencing error rate less than 1%;Q30content(%):测序错误率低于0.1%的碱基序列所占百分比 Sequencing error rate less than 0.1% of base sequence.
采用随机抽样方法对测序获得的序列进行分析,以抽取到的序列数为横坐标、序列数所代表的OTU数目为纵坐标构建稀释曲线。由图1可知,心材和边材曲线均趋于平坦,说明测序数据合理,更多的序列对发现新的OTU贡献率较小。
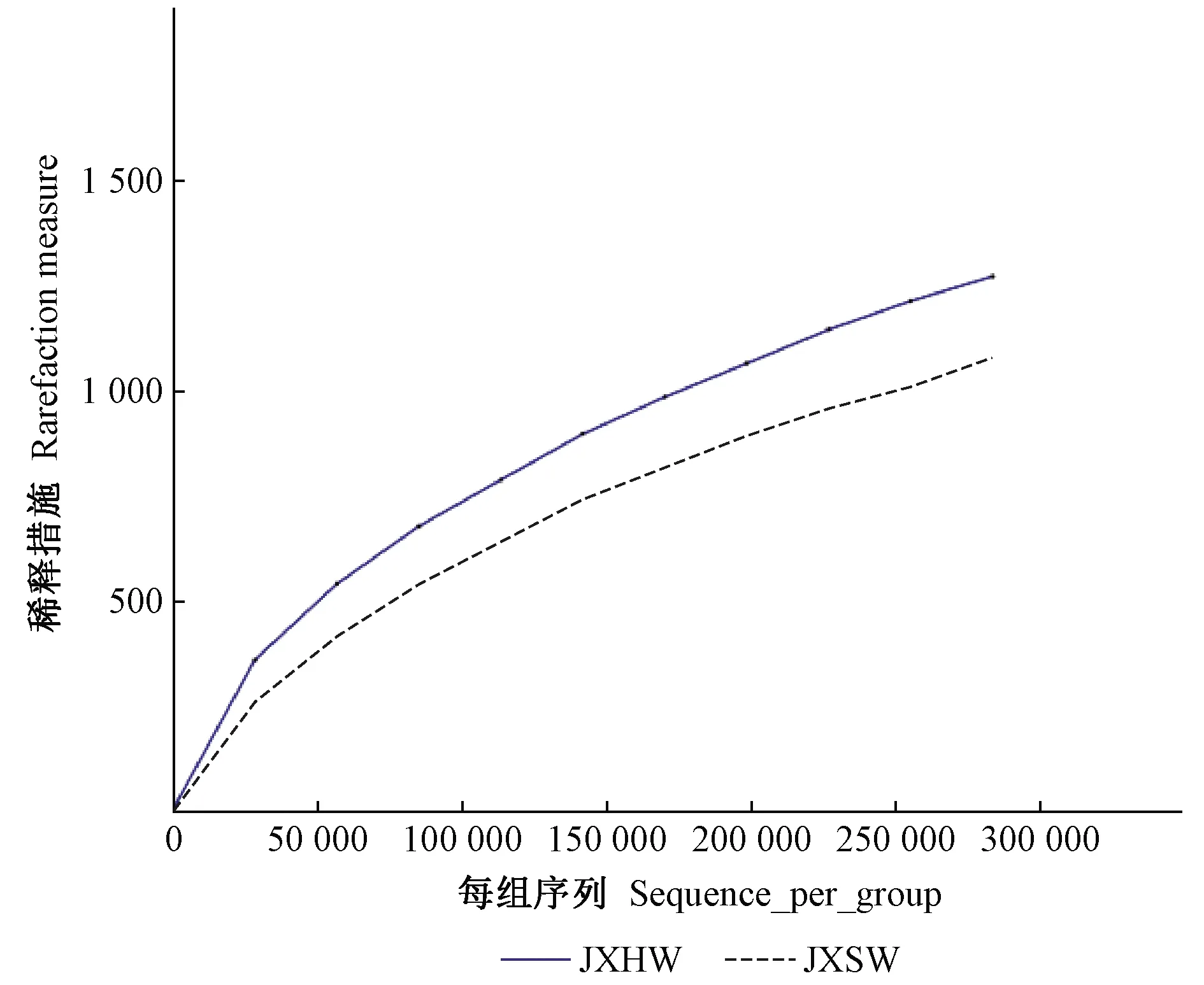
图1 心材和边材内生真菌OTU稀释曲线
2.1.2 内生真菌物种丰富度OTU聚类分析 对6个样品的OTU进行相似性聚类分析见图2。聚类结果Heatmap图展示不同样品中各真菌OTU序列数量的多寡,同时也可以看出不同真菌OTU在6个样品间的分布情况。横向比较可知,6个样品之间的内生真菌相对丰度存在差异,表明即使是同一真菌,不同样品的相对丰度也存在差异,如青霉属(Penicillium)丰度在样品JXSW2和JXHW1相似,在JXHW1和JXW3相似,而在其他2个样本不同;当然在同一个样品,又有内生真菌相对丰度相似,如在样品JXSW3中黑孢属(Nigrospora)、念珠菌属(Candida)、德巴利氏酵母属(Debaryomyces)聚为一支。纵向相比之下,来源于同一株降香黄檀的样本OTU在丰度上聚为一支,如样本JXSW2和JXHW2相似;来源同一部位样品聚为一支,如样本JXHW2和JXHW3相似。
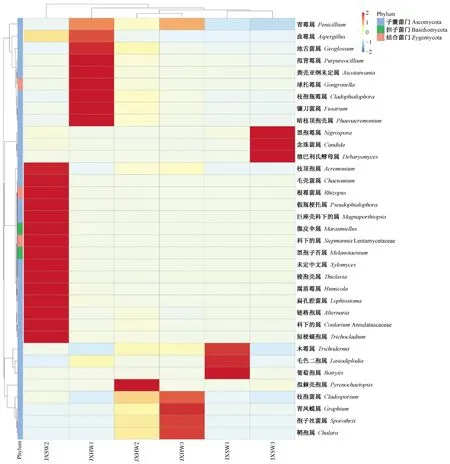
图2 不同样品Heatmap
2.1.3 物种注释及分类学分析 以OTU中出现频数最高的序列为该OTU代表序列,利用OTU代表序列进行分类注释, 采用为UCLUST算法(Edgaretal., 2010),参考序列Unite数据库(Quastetal., 2012)。各样品在各分类水平上的OTU数目统计结果如图3所示。门、科、种分类水平上,各个样本的OTU差别不大;纲和属分类水平上,各个样本的OTU上有一定差距,且心材和边材之间的差别较大,其中降香黄檀心材内生真菌在属级分类水平注释比例相对较高,表明其OUT注释效果较好。
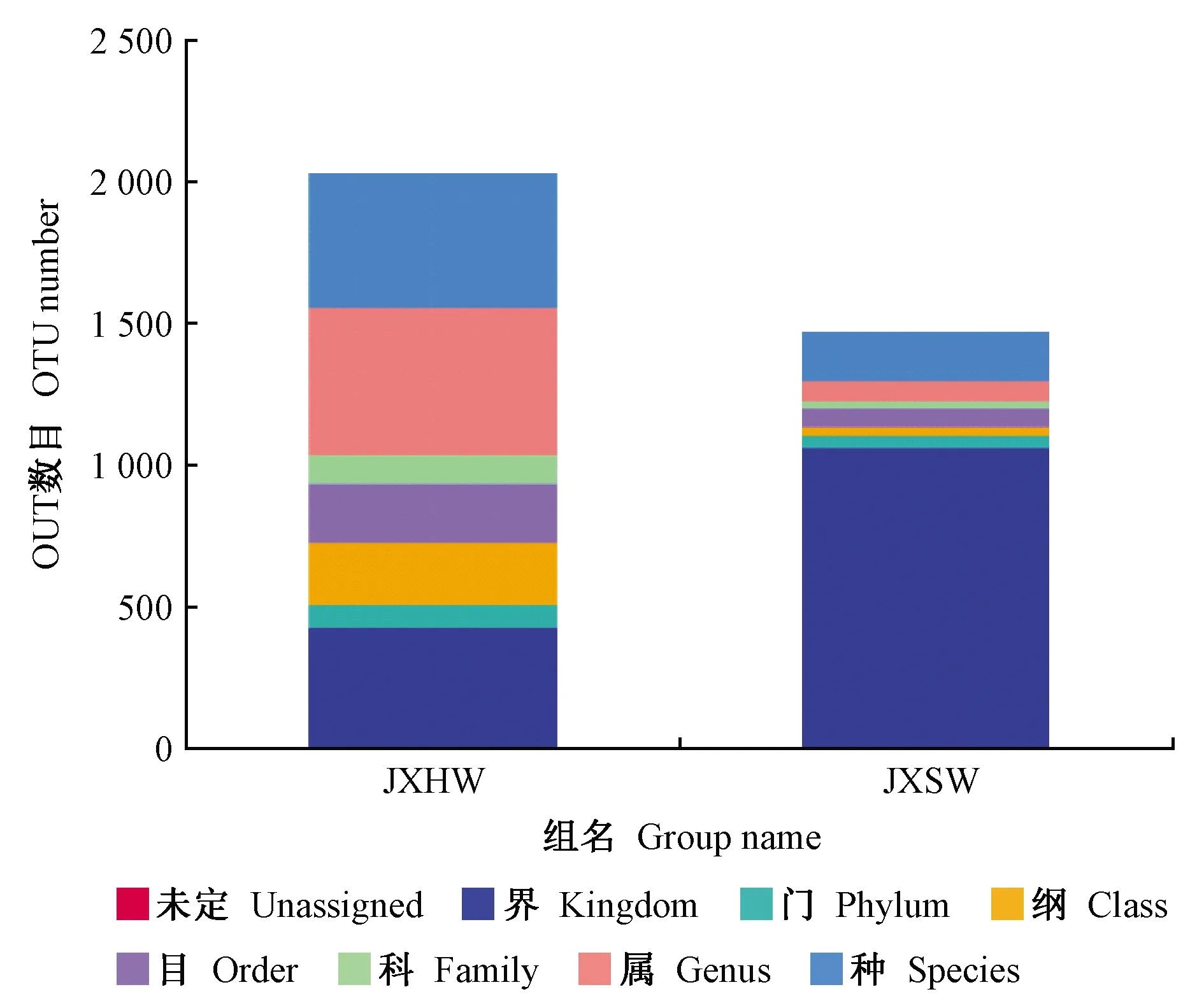
图3 各样品在各分类水平上的序列数目
2.1.4 降香黄檀心材和边材内生真菌群落结构 1)纲水平上的内生真菌群落结构 通过Unite数据库进行归类分析(相似水平为97%)。在纲水平上(图4),降香黄檀木材内生真菌主要菌群依次为伞菌纲(Agaricomycetes)、粪壳菌纲(Sordariomycetes)、散囊菌纲(Eurotiomycetes)、毛霉亚门(Mucoromycotina)、座囊菌纲(Dothideomycetes)、盘菌亚门(Pezizomycotina)、酵母菌纲(Saccharomycetes)、小纺锤菌纲(Atractiellomycetes)、地舌菌纲(Geoglossomycetes)、银耳纲(Tremellomycetes),其中心材内生真菌优势纲为伞菌纲、粪壳菌纲和散囊菌纲,占总数的90%;边材内生真菌优势纲依次为:粪壳菌纲、座囊菌纲、伞菌纲、地舌菌纲和银耳纲,共占95%。
2)属水平上的内生真菌群落结构 通过Unite数据库进行归类分析(相似水平为97%),共获得331个属。分析相对丰度排名前20的属(图5),可以看出降香黄檀心材内生真菌优势菌群依次为暗枝顶孢壳属(Phaeoacremonium)、镰刀菌属(Fusarium)、孢子丝菌属(Sporothrix)、黑酵母样真菌Cladophialophora属和球托霉属(Gongronella),5个优势属共占心材内生真菌的90%。边材内生真菌优势菌群依次为镰刀菌属、毛色二孢属(Lasiodiplodia)、枝孢属(Cladosporium)、念珠菌属(Candida)、德巴利酵母属(Debaryomyces)和木霉属(Trichoderma),共占边材内生真菌的65%。
3)种水平上的内生真菌群落结构 通过Unite数据库进行归类分析(相似水平为97%),降香黄檀心材和边材中内生真菌分属于6门26纲67目116科167属186种。在种水平上,心材优势菌为:孢子丝菌环境种Sporothrixlignivora(7.13%)、卵形孢球托霉菌(Gongronellabutleri)(4.14%)和尖孢镰刀菌(Fusariumoxysporum)(1.95%),其余群落丰度均低于1%。边材优势菌为:可可毛色二孢菌(Lasiodiplodiatheobromae)(0.147%)、酵母菌(Debaryomycesudenii)(0.054%)和镰刀菌(Fusariumpseudensiforme)(0.052%),其余群落丰度低于0.05%。
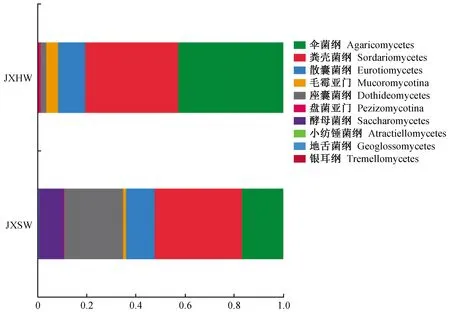
图4 心材和边材纲水平上内生真菌的群落结构
4)心材和边材内生真菌物种差异显著性分析(LEfSe分析) 为进一步探究心材和边材内生真菌种对其划分产生显著差异影响的物种,本研究进行了LEfSe(LDA effect size)分析。心材和边材内生真菌的LEfSe分析结果如图6所示,LDA得分图显示了LDA Score大于4的物种, 可作为代表各组的Biomarker。
降香黄檀心材内生真菌物种丰度大于边材。结合图6 LDA得分图发现,降香黄檀边材中内生真菌种类较少、丰度也较低,且在测序过程中受到植物基因的干扰严重。比对Unite数据库,由物种分支进化图可知,心材和边材内生真菌中有显著差异影响的Biomarker物种有1门2纲5目5科5属2种。其中门水平上具有显著差异影响的菌为接合菌门,纲水平有粪壳菌纲和毛霉亚门,属水平的菌是暗枝顶孢壳属、孢子丝菌属、镰刀菌属、黑酵母样真菌(Cladophialophora)属和球托霉属,种水平的菌为孢子丝菌环境种Sporothrixlignivora和卵形孢球托霉菌(Gongronellabutleri)。
5)内生真菌群落结构多样性分析 在相似性水平97%的条件下,对心材和边材内生真菌多样性系数进行计算(表2)。降香黄檀心材内生真菌的Chao1指数和Observed OTUs指数高于边材,说明降香黄檀心材中内生真菌的物种数量和OTU多于边材。Simpson指数和Shannon指数,表明心材中内生真菌多样性要远多于边材内生真菌多样性。
2.2 降香黄檀心材和边材内生真菌群落结构和多样性与木材理化性质关系
2.2.1 内生真菌群落结构与木材理化性质的关系 降香黄檀心材和边材内生真菌属水平群落结构(丰富度排名前20)与木材理化性质的RDA分析见图7,Axis 1解释51.7%的信息,Axis 2解释44.1%的信息。心材优势菌群相对集中,而边材优势菌则相对分散。pH和钾元素刚好处在含水率、热水抽提物、钙镁元素的反向延长线上,呈极显著负相关。pH、钾元素和灰分与心材优势菌负相关,与边材呈正相关。热水抽提物、含水率和钙镁元素之正好相反。几种木材因子相比之下,暗枝顶孢壳属受到灰分、钾镁元素和pH的影响较大,受含水率、热水抽提物和钙元素影响较小。对于边材优势菌群毛色二孢属和木霉属,木材因子对其影响极小。对于孢子丝菌属,木材因子的影响大小为热水抽提物>含水率>钙>pH>镁>钾>灰分。经过Forword分析表明,pH、灰分、钾元素与降香黄檀内生真菌群落结构有显著相关性。在pH一定的情况下,含水率越高,心材优势菌暗枝顶孢壳属、镰刀菌属、黑酵母样真菌属和球托霉属丰度越高。作为木材内少有的病原拮抗菌木霉属菌,受木材内环境因子的影响最小,且与大部分优势菌负相关。
(4)符合条件的投资人进行认购,该环节完成后,全部资金将转移至信托账户,从而使资金完成从分散到归集再到成为支付承销对价的过程,并最终流向融资人(即发起人)[3]。完成上述步骤后,基础资产所有权归集至信托公司名下。
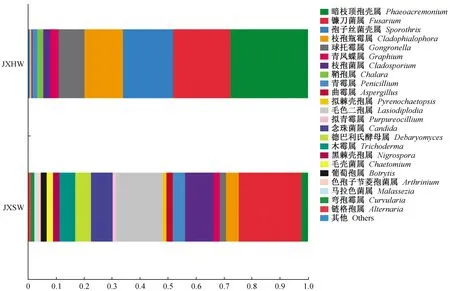
图5 心材和边材属水平上内生真菌群落结构
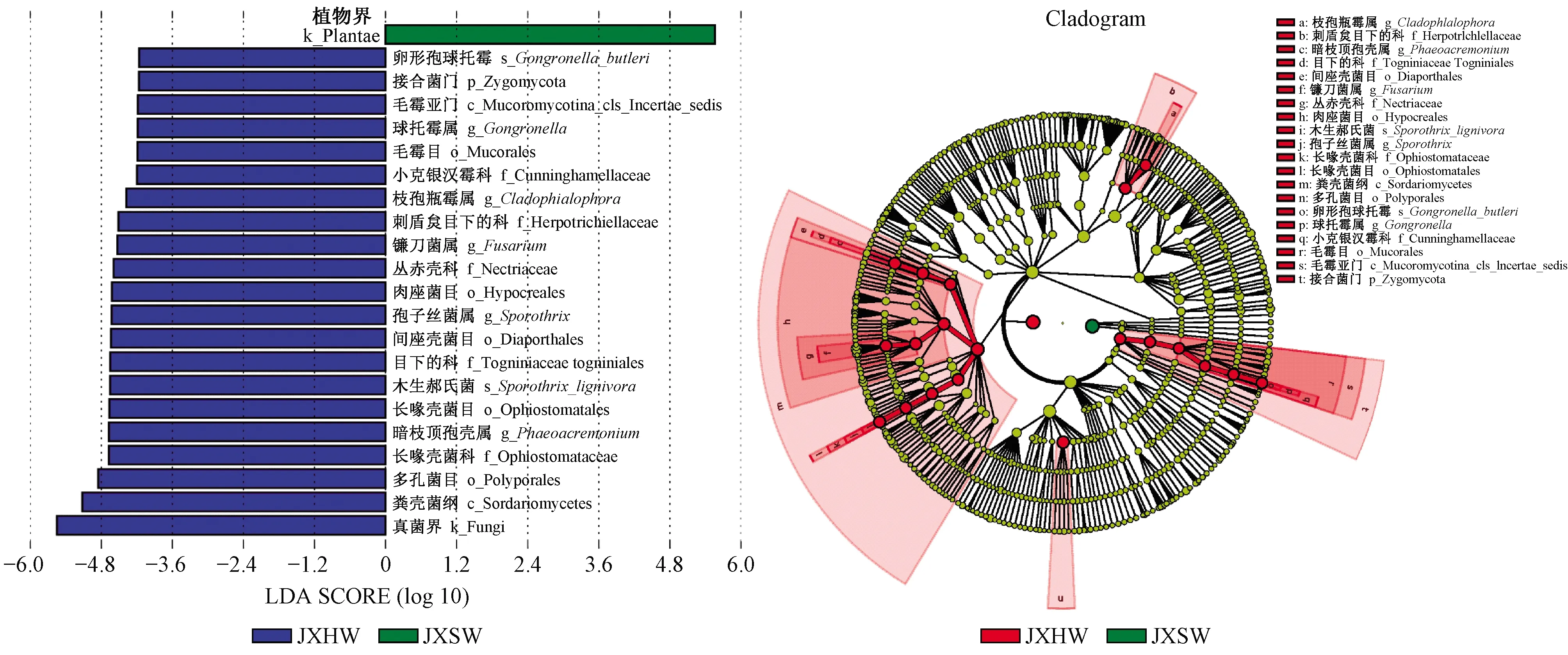
图6 LDA得分与物种分支进化
2.2.2 内生真菌群落多样性指数与木材理化性质的关系 通过对心材和边材样品内生真菌多样性指数和木材理化性质进行相关性分析(表3),结果表明,丰度指数Chao1、多样性指数Simpson、Shannon之间呈正相关,且Simpson、Shannon指数之间呈显著正相关;指数与含水率、热水抽提物、矿质元素钙和镁呈正相关,且与含水率呈显著正相关,表明内生真菌受到含水率的影响较大,与pH、灰分和矿质元素钾呈负相关。
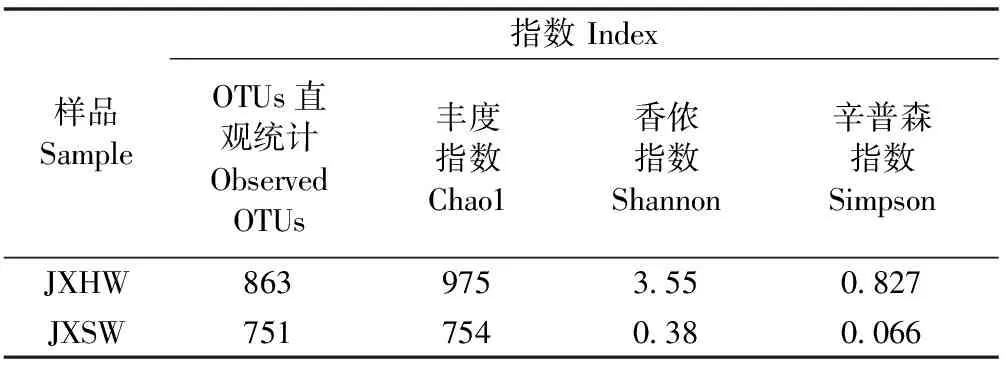
表2 不同样品内生真菌群落的多样性分析(相似度为97%)
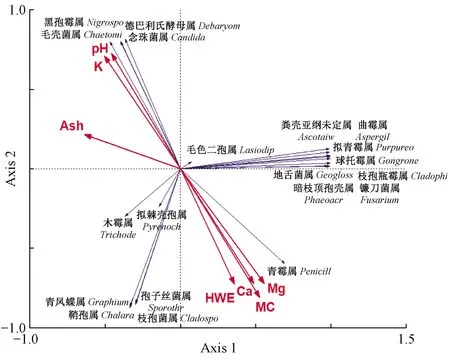
图7 内生真菌菌群与木材性质的RDA分析
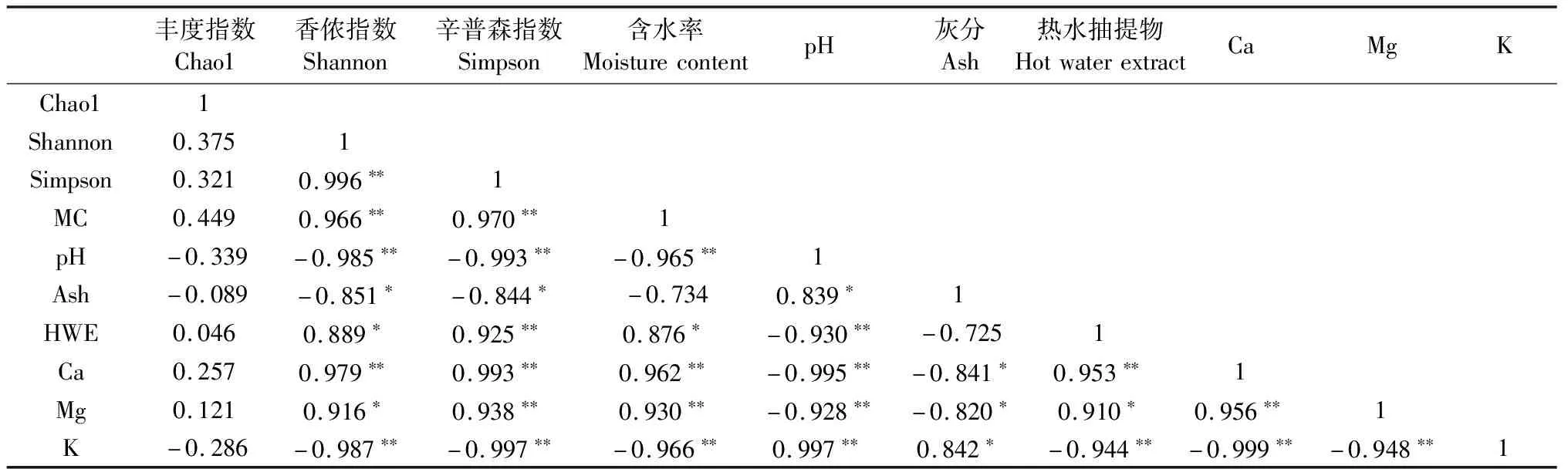
表3 木材理化与内生真菌多样性的相关性分析①
①**:在0.01 水平(双侧)上显著相关,*:在 0.05 水平(双侧)上显著相关。**:Significantly correlated at the 0.01 level(both sides), *:Significantly correlated at the 0.05 level(both sides).
3 讨论
3.1 降香黄檀心材和边材的内生真菌多样性与差异性
目前,有关降香黄檀内生真菌群落多样性对心材的影响尚未报道。本研究通过高通量测序,从基因组水平上解析其心材和边材内生真菌群落结构。多样性分析Observed OTUs 指数和Chao1指数表明,真菌群落OTU数目的种类和各样地真菌的物种总数为心材多于边材。根据Shannon指数和Simpson指数分析结果可知,心材内生真菌群落多样性大于边材,说明降香黄檀心材内生真菌物种丰度和多样性均高多于边材。这表明降香黄檀内生真菌群落结构分布受到本身木材结构性质的影响,同一树种的不同植株内生真菌也会有所不同,可能是因为心材和边材的组织内生理条件和结构性质特征等因素不同,从而影响到内生真菌的侵染和定殖过程,使内生真菌的分布表现出组织差异性(Alyetal.,2011),如Gond(2007)对木橘(Aeglemarmelos)、Gazis(2010)对橡胶树(Heveabrasiliensis)、Kharwar(2010)对药用樟树(Cinnamomumbodinieri)和宋海燕等(2019)对棉花(Gossypium)不同部位的研究。
分析降香黄檀内生真菌群体组成,许多种类分布广泛,在纲水平上,伞菌纲在降香黄檀心材中占比比边材中更大,表明其更加适应心材环境,对心材表现出组织偏好性,该菌纲在不同植物内生真菌都有发现,如蛇足石杉(Huperziaserrata)的茎、叶和根的内生真菌(Chenetal.,2011)、白芨(Bletillastriata)的根和叶的内生真菌(韦艳梅等,2016;席刚俊等,2017)。座囊菌纲和酵母菌纲在边材中的占比比心材要高,在边材中更具有竞争能力,这2个菌纲对心材表现出组织偏好性,座囊菌纲如在巴西仙人掌(Opuntiastricta)的内生真菌(Bezerraetal.,2016)、山苍子(Litseacubeba)的内生菌(杨鼎超,2018),酵母菌纲如在山药(Dioscoreaoppositifolia)地下茎、茎、叶的内生真菌(詹寿发等,2012)。粪壳菌纲和散囊菌纲在心材和边材是优势纲,能适应心材和边材2种不同的环境,在植物内环境环境变换中更稳定,这2个菌纲不表现出组织偏好性,粪壳菌纲如在石斛(Dendrobiumnobile)的内生菌(Chenetal., 2011)、小麦(Triticumaestivum)内生真菌(赵芹等,2017),散囊菌纲如在针叶树(Pinales)的针叶的内生真菌(Yooetal., 2012)、美味石耳(Umbilicariaesculenta)内生真菌(张惠,2014)。同时李永等(2013)报道2个欧美杨(Populus×euramericana)品种的内生真菌优势菌纲,包括在囊菌纲2个分支、散囊菌纲、类壳菌纲等,席刚俊等(2017)报道白芨根的内生真菌优势菌纲为伞菌纲等,说明降香黄檀内生真菌在纲级水平种群组成与其他植物差异不大。在属水平上,镰刀菌属是心材和边材共有的优势属,在心材和边材环境中都具备一定的适应性和稳定性,这个菌属不表现出组织偏好性,是常见的菌属,可侵染和定殖到多种植物,如在豇豆(Vignaunguiculata)种子的内生真菌(Rodriguesetal.,2005)、帝枫皮(Illiciumdifengpi)根的内生真菌(孙文斌等,2018)。同时郑建华等(2013)报道银杏(Ginkgobiloba)优势内生真菌为刺盘孢属(Colletotrichum)、链格孢属(Alternaria)、镰孢菌属(Fusarium)和拟茎点霉属(Phomopsis)为优势菌群,孙思胜(2015)报道在檀香(Santalumalbum)结香部位优势真菌属是镰孢属和拟茎点霉属、檀香健康木材优势真菌属是拟茎点霉属,张苗苗(2017)报道白木香中内生真菌菌种,其中青霉菌属(Penicillium)、可可毛色二孢属、木霉菌属、曲霉菌属(Aspergillus)、镰刀菌属是优势真菌,可见降香黄檀内生真菌在属级水平种群组成与其他植物差异甚大。其原因可能是降香黄檀分布区常年气温较高,内生微生物需耐受高温度;其木材优质、纹理致密、降香油含量高,对内生微生物的侵入定殖和生长要求高。
3.2 降香黄檀内生菌真菌对降香黄檀生长的影响
内生菌在植物体内利用植物产生的营养物质,同时产生代谢产物(Liheng,2012;Wangetal., 2011)。某些内生真菌对植物也有毒害作用。降香黄檀树干中内生真菌中有些真菌类群虽然在很多文献中报道与植物病害相关,如暗枝顶孢壳属的真菌一些为植物的致病菌(张秋娥,2012),能通过树干伤口侵入木质部,并在心材和边材中定殖。如褐枝顶孢霉(Phaeoacremoniumaleophilum)、寄生褐枝顶孢霉(Phaeoacremoniumparasiticum)能够引起葡萄(Vitisvinifera)退化,毛色二孢属的一些菌能够在植物上致病(谢红辉,2016),同时一些能够促进沉香结香(陈旭玉等,2017);但是内生于植物可能只具有较弱的致病性或者已经丧失了致病能力(Schulzetal.,2005; De Brumetal.,2012),甚至转为对植物的有益菌,当然在植物衰退的过程中又可能转化为植物的病源菌(Alyetal.,2011)。如镰刀菌属通常被报道为病原菌,但内生菌镰刀菌属却能降低玉米黑斑病对玉米(Zeamays)的病害严重程度(Leeetal., 2010)。
植物内生真菌对植物的生长具有重大影响,一些木本植物的某些内生真菌能促进植物心材的形成。Cui等(2013)报道木霉属菌和可可毛色二孢属菌能促进白木香心材的形成(Cuietal.,2013)。檀香健康的茎干部位的内生真菌与环境中真菌通过伤口侵染茎干木材,然后形成特殊的真菌群落,进而诱导檀香局部心材的形成(结香)(孙思胜等,2015)。人工接种镰孢菌属系可以诱导沉香结香(郑科等,2016)。多种内生真菌能诱导土沉香结香(谭小明等,2018)。可可毛色二孢菌与腐皮镰孢菌(Fusariumsolani)侵染白木香树体后,会促使树体产生树脂以及胼胝质以抵御真菌的侵染,相应产生一些沉香类物质(谷丽萍等,2018;郑科等,2019)。报道柑橘葡萄座腔菌(Ascomycetes)、镰孢菌属、胶孢炭疽菌(Colletotrichumgloeosporioides)、Khuskia属能促进檀香心材的形成(刘小金,2012)。降香黄檀的某些内生真菌对其心材的形成也存在促进作用,如茄病镰刀菌(Fusariumsolani)、层生镰刀菌(Fusariumproliferatum)、多毛孢(Pestalotiopsispalmarum)和红褐肉座菌(Hypocreajecorina)均能促进降香黄檀产生心材,其中以多毛孢效果最佳(如贾瑞丰,2014)。降香黄檀内生真菌资源的利用还需进一步深入的研究。
3.3 降香黄檀内生真菌群落的影响因素
植物内生真菌的来源主要是根际土壤微生物和空气中的微生物(Compantetal.,2011),很多因素都会影响植物内生真菌的群落结构(Hoffmanetal.,2008),如根际土壤的质地、植物生长环境、植物本身内部微环境性质、基因水平以及人类生产活动等都会影响到植物内生真菌的群落结构。本研究主要是对引起降香黄檀内生真菌群落结构差异的主要木材内环境因子进行研究,造成降香黄檀心材和边材内生真菌群落数量和结果差异其机制是多方面而又较为复杂的,需要深入研究。
本研究结果初步显示,降香黄檀心材和边材内生真菌群体在群落结构的组成和潜在功能上具有一些独特特征。降香黄檀因其样本珍贵以及国家政策等原因,考虑到群落结构及理化性质结果与实际情况的差距,未来的研究工作还可以选择对正常生长的降香黄檀直接取样进行研究。研究不同地区降香黄檀内生菌是否存在种属专一性和环境差异性,以及内生菌的功能多样性等方面进行研究,以揭示降香黄檀内生菌群落结构多样性特征和其生态分布规律;研究降香黄檀内生菌与心材互作的分子机制等,为利用内生微生物促进降香黄檀心材形成提供理论基础等研究。
4 结论
对降香黄檀心材和边材的内生真菌HiSeq高通量测序并优化后,结果表明,降香黄檀心材和边材内生真菌种群资源丰富多样,其中在内生真菌的不同分类水平上心材和边材内生真菌群落结构具有显著差异影响的内生真菌有2纲5属2种,且心材内生真菌群落丰度和多样性均高于边材,同时也说明了降香黄檀心材和边材内生真菌的入侵定殖既有相似又有区别,同时又具有组织偏好性和专一性。
降香黄檀心材和边材的内生真菌与木材理化性质的关系中,引起降香黄檀内生真菌群落结构差异的主要木材环境因子有pH、灰分和矿质元素钾。而引起降香黄檀内生真菌群落结构多样性差异的主要木材内环境因子是pH、含水率、矿质元素钙和钾,且含水率影响最大。
