矮牡丹与芍药属其他5个种叶绿体基因组特征的比较
2020-06-02周晓君彭正锋孙姗姗押辉远张延召程彦伟
周晓君 张 凯 彭正锋 孙姗姗 押辉远 张延召 程彦伟
(1.洛阳师范学院生命科学学院 洛阳 471934; 2.河南小秦岭国家级自然保护区管理局 三门峡 472500;3.洛阳牡丹研究院 洛阳 471000)
牡丹是芍药属牡丹组(Paeoniasect.Moutan)植物的统称(APG Ⅳ, 2016)。除了以“花中之王”的美誉供观赏之外,牡丹还有很高的经济价值:其根皮“丹皮”为复方中的常用药材之一,列入中国药典;牡丹籽油富含α-亚麻酸等不饱和脂肪酸,是很有发展前途的健康食用油之一(洪德元等, 2017)。由于牡丹具有较高的观赏和经济价值,在世界各地广泛栽培。中国特有的野生牡丹一直被国内外视为珍贵的种质资源,开展野生牡丹系统进化的研究以及栽培牡丹起源的研究对阐明牡丹的起源、培育和改良栽培品种具有重要的理论和实践价值(Zhaoetal., 2008)。叶绿体普遍存在于陆地植物、藻类和部分原生生物中,是进行光合作用的关键场所,是具有自主遗传信息的重要细胞器(邢少辰等, 2008)。典型的被子植物叶绿体基因组为环状四分体结构,即2个反向互补重复序列(inverted repeat,IRa和IRb)、大单拷贝区(large single-copy,LSC)和小单拷贝区(small single-copy,SSC)。与核基因组相比,叶绿体基因组较小,序列保守,结构稳定,遗传重组率低,属于母系遗传,其结构和序列信息在揭示物种起源、进化演变及物种间亲缘关系等方面具有重要价值(Mccauleyetal., 1996; 王玲等, 2012)。
近年来,先后有不同学者对芍药属植物叶绿体基因组进行了研究。Zhang等(2016)对芍药属芍药组(PaeoniaSect.Paeonia)药用植物川赤芍(P.veitchii)进行叶绿体基因组测序;Li 等(2018)对芍药属牡丹组中仅有的2种开黄花的野生牡丹大花黄牡丹(P.ludlowii)、滇牡丹(P.delavayi)的叶绿体进行了测序并分析其特征;Guo等(2018)对油用凤丹牡丹(P.ostii)进行了绿叶体测序并将其与其他几种芍药属植物的叶绿体特征进行了对比。作为7种野生牡丹之一,矮牡丹(P.jishanensis)被认为是现代栽培牡丹品种重要的祖先种(Yuanetal., 2014; Xuetal., 2016)。对矮牡丹叶绿体基因组特征进行分析,并将其与已发表的其他5种芍药属植物叶绿体基因组进行对比,旨在为牡丹种质资源的开发和利用提供科学依据,也可为今后芍药属植物系统进化关系研究及种质资源的鉴定提供基础性数据。
1 材料与方法
1.1 试验材料
用于叶绿体基因组测序的矮牡丹组织样本采集于山西省稷山县马家沟(35°43′N, 110°58′E)。利用天根试剂盒(天根,北京)提取基因组DNA备用。
1.2 总DNA测序
将检测合格的DNA样本用超声波进行片段化,经纯化后进行末端修复、3′端加A及连接测序接头,用琼脂糖电泳选择合适片段大小。通过PCR扩增后构建测序文库,质检合格的文库利用Illumina HiSeq 2500平台进行测序。
1.3 组装与注释
测序获得的原始数据利用CLC Genomics Workbench 9.0软件(CLC Bio,Aarhus,Denmark)过滤去除低质量序列(参数设置为:Ambiguous limit=2,Quality limit=0.05)(Wambuguetal., 2015)。以大花黄牡丹叶绿体基因组(KY817592)作为参照(Lietal., 2018),将过滤后的11 322 016条reads采用Hahn等(2013)的组装方法构建叶绿体基因组。最终得到长度为152 628 bp的矮牡丹叶绿体全基因组序列。共有145 741条叶绿体reads参与组装,平均覆盖率为141.9×。利用Geneious 8.0软件进行矮牡丹与大花黄牡丹序列的CLUSTAL比对,进行碱基修正、修改蛋白质和tRNA等注释,完成叶绿体基因组的分区。采用OGDraw在线工具(http:∥ogdraw.mpimp-golm.mpg.de/)进行基因组图谱绘制。
1.4 叶绿体基因组特征分析

图1 矮牡丹叶绿体基因组物理图谱
利用MISA(MicroSatellite identification tool)软件(http:∥pgrc.ipk-gatersleben.de/misa/)搜索矮牡丹叶绿体基因组中SSR位点并分析其特征。具体参数为:单核苷酸、二核苷酸、三核苷酸、四核苷酸、五核苷酸和六核苷酸为基序(Motif)的重复数分别设置为8、5、4、3、3、3,2个复合SSR位点间隔碱基数最大为100。用EMBOSS 6.4.0软件(http:∥emboss.open-bio.org/)分析蛋白编码基因密码子偏好性(Riceetal., 2000)。从GenBank下载凤丹牡丹、大花黄牡丹、滇牡丹、川赤芍和草芍药5个种的叶绿体基因组全序列,与本项目矮牡丹叶绿体基因组序列进行多重比对。
2 结果与分析
2.1 叶绿体基因组基本特征
与已发表的其他芍药属叶绿体基因组类似,矮牡丹叶绿体基因组为包含LSC、SSC、IRa和IRb 4个区段的共价闭合双链环状分子(图1),序列全长152 628 bp,GC含量为38.4%。其中LSC区段长84 292 bp,SSC区段长17 044 bp,IR区段为25 646 bp(Zhouetal., 2019)。基因组的GC含量是判断物种间亲缘关系的重要指标,矮牡丹LSC、SSC、IR区段GC含量与其他芍药属一致,分别为36.7%、32.7%、43.1%(Guoetal., 2018; Lietal., 2018)。在基因组大小方面,矮牡丹叶绿体基因组比凤丹牡丹大475 bp,比川赤芍、大花黄牡丹、滇牡丹和草芍药分别小54、59、70、70 bp(表1)。
叶绿体注释结果表明,矮牡丹叶绿体基因组共有112个基因,包括蛋白编码基因78个、tRNA基因30个和rRNA基因4个(表2)。其中,有19个基因(包括4个rRNA、7个tRNA、8个蛋白编码基因)在IR区重复。LSC区段含有22个tRNA基因和60个蛋白编码基因,SSC区段含有1个tRNA基因和10个蛋白编码基因。研究表明,内含子在基因的表达调控中起着重要作用,许多内含子能够增强外源基因在植物特定时间及特定部位的高水平表达,从而产生相关的农艺性状(Jiaoetal., 2012)。矮牡丹蛋白编码基因中,9个基因(atpF,ndhA,ndhB,petB,petD,rpl16,rpl2,rpoC1,rps16)含有1个内含子,3个基因(clpP,rps12,ycf3)含有2个内含子。

表1 芍药属6个种叶绿体基因组基本特征比较①
①LSC: Large single-copy region; SSC: Small single-copy region; IR: Inverse repeats.

表2 矮牡丹叶绿体基因组基因信息①
①*: 具有内含子的基因;a: 重复基因(存在于IR区域的基因)。*: Genes containing introns; a: Duplicated gene(genes present in the IR regions).
2.2 矮牡丹叶绿体基因组密码子使用情况
在矮牡丹叶绿体基因组中,共有25 988个密码子,编码蛋白78个,其中编码亮氨酸的密码子数量最多,为2 716个,异亮氨酸、色氨酸的密码子数量分列第2(2 223个)和第3(1 984个)。TAA为常用的终止密码子(43个),略高于TAG(21个)和TGA(21个)。矮牡丹叶绿体基因组中蛋白编码基因密码子偏好使用A/T碱基。在所有密码子中,第1个、第2个和第3个碱基为A/T的分别占总密码子数的54.04%、61.59%和68.89%。
2.3 叶绿体基因组SSR分析
在矮牡丹叶绿体基因组中搜索到143个SSR位点。其中,141个为普通型SSRs,2个为复合型SSRs;单核苷酸重复Motif位点最多为116个,二核苷酸、三核苷酸、四核苷酸和五核苷酸重复基序分别有13、6、5、1个,没有发现六核苷酸重复基序(表3)。143个SSRs位点中,A/T组成的位点有134个,仅有9个位点含有G/C,这表明SSRs的碱基组成偏好;从分布区段上看,14个位点位于IR区段,103个位于LSC区段,26个位于SSC区段,说明矮牡丹叶绿体基因组SSRs分布的不均匀性。
2.4 芍药属6个种的叶绿体基因组IR边界分析
植物叶绿体基因组IR的扩张与收缩是较为普遍的现象,它决定了不同植物叶绿体基因组大小(Huangetal., 2014)。为了探索IR潜在的扩张与收缩,比较了芍药属6个种的叶绿体基因组IR与SC边界区域基因的分布状况(图2)。如图所示,LSC/IR和SSC/IR边界分布的基因包括rps19,rpl2,ycf1,ndhF和trnH。芍药属6个种的叶绿体基因组SSC/IRa边界均为ycf1基因,且在不同的区域内ycf1基因分布长度不同,在草芍药、凤丹牡丹、大花黄牡丹、滇牡丹、川赤芍、矮牡丹的SSC区域内长度分别为4 325、4 352、4 340、4 355、4 327和4 346 bp,在IRa区域内的长度分别为1 087、1 078、1 078、1 078、1 076和1 078 bp。同样地,trnH基因在6个种中均位于LSC区域内,分别距IRa/LSC边界1、79、0、1 534、3、3 bp。rpl2基因仅在凤丹牡丹中位于LSC/IRb边界,与其他5个种不同。ndhF基因位于5个种的IRb/SSC边界,而在川赤芍中位于SSC区域距边界20 bp。

表3 矮牡丹叶绿体基因组SSRs类型信息
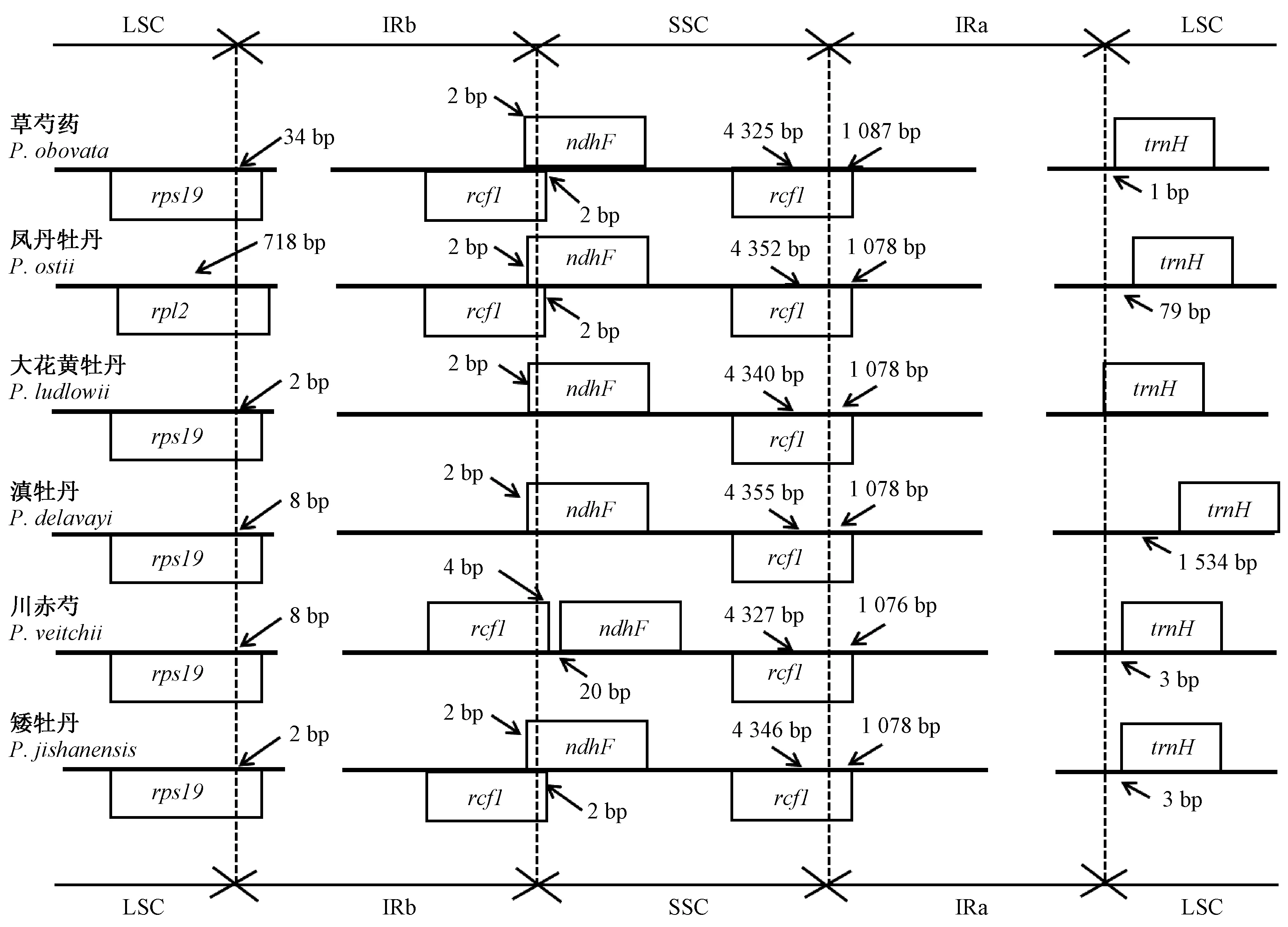
图2 芍药属6个种的叶绿体基因组反向重复区(IR)、大单拷贝区(LSC)和小单拷贝区(SSC)边界比对
3 讨论
叶绿体基因组序列包含着解决复杂系统发育关系的诸多信息,被广泛用于植物系统发育及农作物栽培种进化关系重建等领域(Cuénoudetal., 2002; 谢海坤等, 2017)。近年来,随着高通量测序技术的飞速发展,基因组测序成本大大降低,叶绿体基因组获得更加便捷。大多数被子植物叶绿体基因组都属于典型的质体基因组4部分结构,即1对IR区将整个基因组划分成2个单拷贝区域;都具有74个蛋白编码基因,少数植物会在此基础上增加5个蛋白编码基因(Millenetal., 2001)。矮牡丹cp DNA基因组具有蛋白编码基因78个、tRNA基因30个和rRNA基因4个。通过比较分析矮牡丹与芍药属其他5个种的叶绿体基因组序列可发现,从基因组大小和基因内容来看,芍药属叶绿体基因组高度保守。
密码子是遗传信息正确表达的关键。一般情况下,叶绿体基因组DNA上的起始密码子序列分别为ATG、ATT、ATA。本研究中矮牡丹叶绿体基因组起始密码子中除了ATG是最常见起始密码子外,还有ATA、GTG、ATT 3种类型,这在其他被子植物中也有报道(Bortirietal., 2008)。不同叶绿体基因组的碱基组成有一定的差异,矮牡丹叶绿体基因组偏向于使用含有A/T的密码子。密码子偏好性与生物长期的进化历史密切相关,因此研究矮牡丹密码子的使用情况对芍药属物种进化模式的探究具有重要作用。SSR是一种重要的分子标记,常用于物种的居群遗传学分析(Doorduinetal., 2011)。对矮牡丹叶绿体基因组中SSR的统计结果显示,其SSR中含有丰富的A/T,而且多聚的A/T类型远大于多聚的G/C类型,与地黄属(Rehmannia)植物类似(Zengetal., 2017)。矮牡丹叶绿体基因组SSR位点的发现对芍药属植物鉴定、群体水平的多态性检测及系统发育分析具有一定的意义。
不同物种间叶绿体基因组IR/SC边界位置的变化是较为普遍的现象,这些变化通常在同一科的物种中也会有所不同。研究表明,芍药属6个种的叶绿体基因组尽管在结构和大小方面比较保守,但不同物种间IR和SC的边界位置仍有一定变化。与其他5个种相比,凤丹牡丹的IR(24 863 bp)是6个种中最小的,存在收缩现象,这主要表现在LSC/IR的rpl2基因上。该基因在凤丹牡丹中有718 bp延伸至LSC区域,而其他种的rpl2基因均完整地位于IR区。所以芍药属LSC/IRb的边界变化是IR区扩张与收缩的主要原因,这与地黄属叶绿体基因组的研究结果一致(Zengetal., 2017)。
芍药科(Paeoniaceae)在被子植物中十分孤立,其系统位置一直存在激烈争论。基于分子系统学的被子植物系统发生显示:芍药科与虎耳草科(Saxifragaceae)、景天科(Crassulaceae)和金缕梅科(Hamamelidaceae)等构成一大支,即虎耳草目(Saxifragales)(APG Ⅳ, 2016)。但芍药科在虎耳草目中究竟与哪类植物最近,仍然是一个悬而未决的问题(洪德元等, 2017; Jianetal., 2008)。本研究可为今后芍药科植物系统进化关系研究提供基础性数据。
4 结论
芍药属6个种的叶绿体基因组为典型的包含4个区段的共价闭合双链环状分子,GC含量均为38.4%,且在LSC、SSC、IR区段GC含量一致,所有芍药属植物叶绿体基因组从长度到GC含量均高度保守。矮牡丹叶绿体基因组中,共有25 988个密码子,编码亮氨酸的密码子数量最多,为2 716个,矮牡丹叶绿体基因组偏向于使用含有A/T的密码子。矮牡丹叶绿体基因组中共有143个SSR位点,单核苷酸重复Motif位点最多为116个,SSR中多聚的A/T类型远大于多聚的G/C类型。明确了6种芍药属植物在每个区段的不同,指出LSC/IRb的边界变化是IR区扩张与收缩的主要原因。
