澳门生态一区和二区水鸟谱系多样性的季节变化
2020-04-13丁志锋陈述植诗雅谭金庆梁健超冯永军
丁志锋, 陈述, 植诗雅, 谭金庆, 梁健超, 冯永军
(1. 广东省生物资源应用研究所,广东省动物保护与资源利用重点实验室,广东省野生动物保护与利用公共实验室,广州510260;2. 澳门特别行政区市政署园林绿化厅自然护理处,澳门; 3. 澳门特别行政区市政署园林绿化厅自然保护研究处,澳门)
生态学家长期以来一直认为环境异质性与物种多样性密切相关(Alloucheetal.,2012;Seiferlingetal.,2014;Steinetal.,2014;Tonkinetal.,2017)。实际上,环境条件随时间的周期性变化是全球生态系统的重要特征之一,而年度季节性变化是其中最为强烈和最令人熟知的表现形式。随着时间变化,生物群落很少保持稳定,这在鸟类群落中尤为明显(de Casenaveetal.,2008),食物和觅食方式的季节变化寻常可见。因此,仅仅分析单个季节的生物群落结构特征,可能提供不完整的信息,进而影响到生物多样性保护政策的制定和实施。
季节性常常用于解释一些普遍现象,如群落结构(Chesson,2000;Chase,2011)、生活史适应性(McNamara & Houston,2008)、鸟类迁徙动态(Somveilleetal.,2015)等。特别地,利用谱系多样性来探讨鸟类的季节性变化和推断群落聚群规律越来越受到关注(Emerson & Gillespie,2008;Cheetal.,2019)。如当环境因子驱动鸟类群落构建时,谱系多样性指数趋向于聚集;当物种相互作用主宰着群落组成时,谱系多样性指数趋向于发散;当群落随机分布时,谱系多样性指数则表现为随机。Gomez等(2010)发现在温带和热带森林中,夏季鸣禽群落通常受到生物相互作用的影响;Sebastián-González和Green(2017)的研究结果表明,越冬水鸟的谱系聚集模式占据了主导地位,可能与水鸟相似的微生境偏好有关。Che等(2019)通过研究香港特别行政区米埔湿地水鸟群落谱系多样性的季节变化,发现夏季水鸟群落呈现聚集结构,冬季水鸟群落呈现发散结构,推断夏季水鸟群落受到环境过滤的驱动,而冬季受到物种相互作用的影响。这些研究结果表明,季节性变化决定着各种群落构建因子作用力的相对强弱,为了解群落聚群规律提供了更为完整的视角。
基于此,我们对澳门路氹城2块湿地(生态一区和生态二区)的鸟类季节性变化进行分析:(1)2块湿地的鸟类物种和谱系多样性是否存在季节性差异?(2)2块湿地鸟类群落构建机制是否存在季节性差异?根据已有研究的结果,我们预测2块湿地鸟类的谱系结构存在季节变化:在繁殖季(3—5月)表现为聚集趋势(环境过滤驱动),而在越冬季表现为发散趋势(物种相互作用影响)。
1 研究地和方法
1.1 研究地概况
澳门特别行政区(113°32′22″E,22°12′40″N)由澳门半岛、氹仔岛和路环岛组成,嘉乐庇总督大桥、友谊大桥和西湾大桥把澳门半岛和氹仔岛连接起来,而路氹填海区把氹仔和路环2个离岛连为一体。澳门位于亚热带地区,年平均气温为22.8 ℃,最高气温35.8 ℃,最低气温4.6 ℃,年平均降水量1 795.6 mm(据澳门特别行政区政府统计暨普查局2018年环境统计,https://www.dsec.gov.mo/TimeSeriesDatabase.aspx)。生态一区紧靠莲花大桥西北,是面积约15 hm2的封闭区,区内有滩涂、芦苇和树林,优势树种为细叶榕Ficusmicrocarpa、血桐Macarangatanarius、黄槿Hibiscustiliaceus、苦楝Meliaazedarach、桐花树Aegicerascorniculatum和秋茄Kandeliaobovata,并有出入水管与十字门水道相连通,中间小岛上分布着天然草被灌丛,水边芦苇茂密,人工栽植的黄槿和细叶榕环绕在四周。生态二区位于路氹填海区的西岸,面积约40 hm2,区内有3个人工小岛,分布有澳门面积最大的红树林,主要植物有老鼠簕Acanthusilicifolius、桐花树和秋茄等。2块湿地的具体位置见图1。
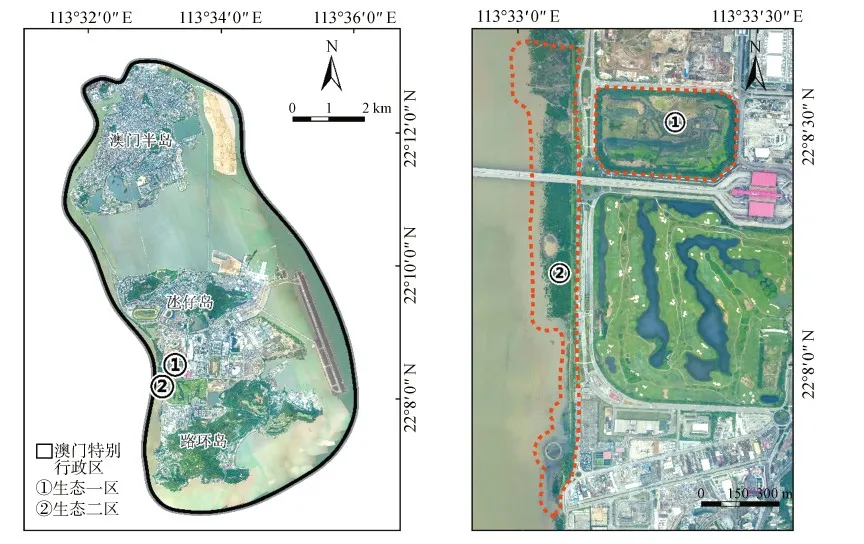
图1 研究样地位置Fig.1 Location of study sites
1.2 鸟类调查方法
2018年1—12月,采用样线法(Bibbyetal.,2000)对2块湿地进行鸟类逐月调查,使用单筒望远镜(Swarovski 20~60倍)和双筒望远镜(Olympus 8×42倍)记录鸟类的种类和数量:当鸟类数量较少时,采用直接计数法;当鸟类集大群(数量超过200只)又处于快速活动时,采用集团估算法,即先估计集群的总数,然后估计集群中主要种类的比例,按主要种类的比例计算各物种的数量。调查仅在天气良好的情况下进行。按照Che等(2019)的方法划分研究区季节:3—5月为春季,6—8月为夏季,9—11月为秋季,12—2月为冬季。
1.3 多样性指数
利用物种丰富度和多度来表征群落的分类多样性,其中多度以最大值保留法(Howes & Bakewell,1989)计算,即从数次调查的同种鸟类统计数值中保留最大值代表该鸟类的数量。谱系多样性指数采用Faith’ PD(Faith,1992),计算方法为:从birdtree.org下载5 000个基于Hackett的谱系树[鸟类的谱系关系源于Jetz等(2012);分类系统参照Jetz等(2012)并遵循Birdlife V3 Word List(9 895种现存物种被确认,2010年6月)],通过TreeAnnotator version 1.10.4获得最大分支置信树,并基于R语言picante程序包计算谱系指数和利用FigTree v1.4.4生成谱系树。
利用基于物种多度加权的平均成对谱系距离(abundance-weighted mean pairwise phylogenetic distance,MPD)来表征群落谱系结构(Webbetal.,2002)。
(1)
式中,n为群落组成物种数量,δi,j是物种i和物种j的谱系距离(欧式谱系距离),fi和fj分别为物种i和j在群落中的相对多度(群落中某一物种的多度占所有物种的多度之和的百分比)。
此外,通过比较MPD实际值与999次随机值来推断群落谱系结构是否与随机期望存在差异。随机的方法为对群落中物种谱系进行种名随机互换运行999次,进行1 000次迭代生成999个随机群落,计算MPD的标准化效应值(standardized effect size of MPD,SES),其公式为:
(2)
式中,Obs为MPD的观测值,Exp是999次零模型模拟的平均值,SDexp是999次零模型模拟的标准差。
当SES<0,则表明环境过滤可能驱动群落聚集;如果SES>0,则表明竞争排斥可能驱动群落发散(Webbetal.,2002)。进一步,当SES>1.96或SES<-1.96时,则认为SES显著高于或低于随机值(P<0.05)。
数据统计分析均在R 2.13.1中完成(R Core Team,2011)。
2 结果
共记录水鸟37种,隶属于6目9科,其中,鸻形目Charadriiformes物种数最多,为15种,其次为鹈形目Pelecaniformes 11种,其余各目皆在10种以下;鹭科Ardeidae鸟类最多,为11种,其次为鹬科Scolopacidae鸟类9种,其余各科皆在5种以下(图2)。白鹭Egrettagarzetta多度最高,为540只,其次为绿翅鸭Anascrecca(295只)、夜鹭Nycticoraxnycticorax(216只)和大白鹭Casmerodiusalbus(209只),其余各物种皆在200只以下。
生态一区和二区的物种丰富度在冬季最高,其次为秋季。生态一区的物种多度在秋季最高,春、夏季较低;生态二区的物种多度在冬季最高,夏、秋季较低。PD和MPD的季节变化规律与物种丰富度相似,即冬季最高(生态二区的MPD在秋季最高),春、夏季较低(图3)。
在谱系结构方面,生态一区和二区呈现出不同变化(图4):生态一区的鸟类群落在春、秋和冬季趋向于谱系发散,在夏季呈现出显著的谱系聚集;生态二区的鸟类群落在春、夏季呈现出显著的谱系聚集,在秋季趋向于谱系发散,冬季趋向于谱系随机(接近0)。
3 讨论
本文通过分析澳门特别行政区2块湿地的水鸟谱系多样性的季节变化模式来探讨水鸟群落构建机制,发现水鸟群落的物种和谱系多样性均呈现出明显的季节变化模式,谱系发散和聚集具明显的季节差异,群落构建因子的相对重要性在不同季节之间发生了显著变化。
张敏等(2009)的研究结果显示(调查时间段为2006年7月—2008年2月),路氹离岛的水鸟优势种为白鹭、环颈鸻Charadriusalexandrinus、绿翅鸭、池鹭Ardeolabacchus、苍鹭Ardeacinerea和夜鹭等;在间隔10年后,调查结果显示水鸟优势种为白鹭、绿翅鸭、夜鹭、大白鹭、青脚鹬Tringanebularia和环颈鸻等,10年前后优势种的组成略有变化,这可能与物种周转(时间尺度上的物种组成变化)有关。如在斑块生境中,所有斑块平均物种周转率为每年30.4%,表明每年超过30%的鸟类发生了周转,其群落组成是动态变化的(Sietal.,2014);此外,也可能与鸟类生境的改变有关(张敏等,2009)。
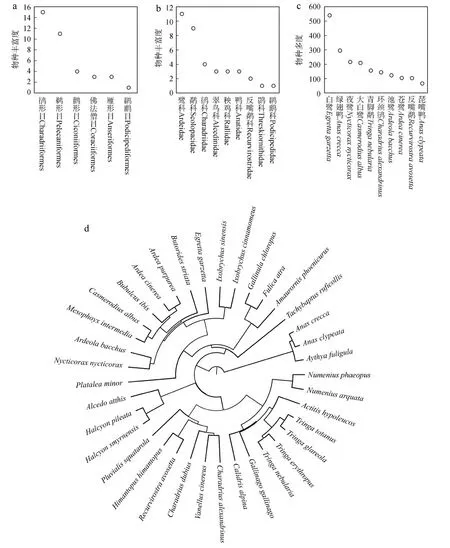
图2 澳门记录的37种水鸟目的物种数(a)、科的物种数(b)、物种多度(c, 前10物种)和谱系树(d)Fig. 2 Species richness across orders (a) and families (b), species abundance (c, top 10 abundant species),and phylogenetic tree (d) of the 37 waterbird species recorded in Ecological Zone Ⅰ and Ⅱ in Macao, China
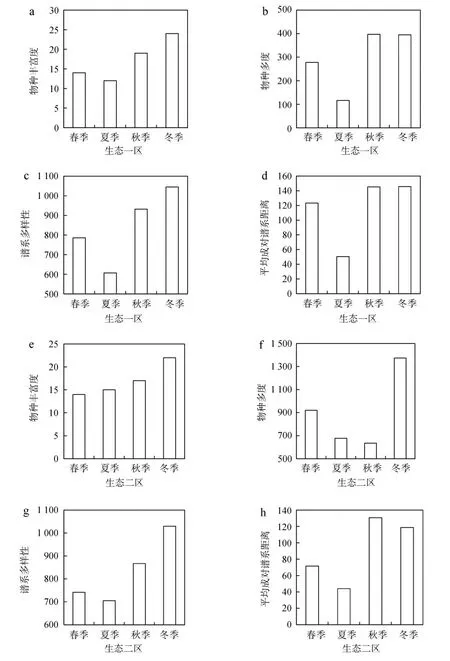
图3 澳门生态一区和二区水鸟的丰富度(a、e)、物种多度(b、f)、谱系多样性(c、g)和平均成对谱系距离(d、h)的季节变化Fig. 3 Seasonal dynamics of richness (a, e), abundance (b, f), phylogenetic diversity (c, g) and abundance-weightedmean pairwise phylogenetic distance (d, h) of waterbird species in Ecological Zone Ⅰ and Ⅱ in Macao, China
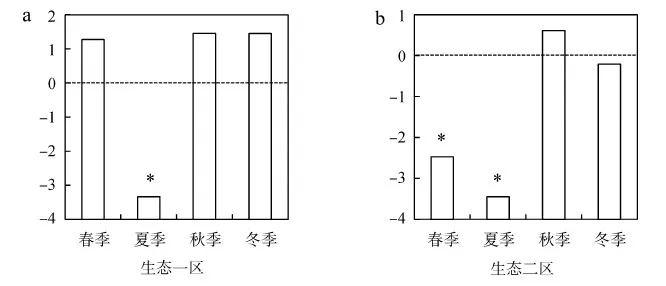
图4 澳门生态一区(a)和二区(b)水鸟平均成对谱系距离的标准化效应准值的季节变化Fig. 4 Seasonal dynamics of the standardized effect size of abundance-weighted mean pairwisephylogenetic distance of waterbird in Ecological Zone Ⅰ (a) and Ⅱ (b) in Macao, China
*P<0.05
澳门路氹湿地是多种候鸟重要的迁徙停歇地和越冬地,被列入亚洲的重点鸟区之一(Chanetal.,2009)。该区域春、秋季的候鸟迁徙形成了鸟类丰富度和多度的季节变化:由于候鸟的秋季迁入自8月开始,因此秋、冬季生态一区和二区鸟类物种数逐渐增加,丰富度在冬季最高,秋季次之;多度亦呈现出相似规律。这与杭州湾、钱塘江河口水鸟和海南北黎湾、后水湾湿地水鸟的季节变化类似(杨灿朝等,2009; 蒋科毅等,2011)。此外,蒋科毅等(2011)发现在杭州湾及钱塘江河口春季鸻鹬类迁徙高峰在数量上远超过秋季迁徙高峰,达到了后者的4倍以上,类似的研究结果也出现在崇明东滩(Maetal.,2002),这可能与该区域在鸻鹬类迁徙路线中的特殊地理位置有关。本研究结果则与之不同:春、秋季鸻鹬类迁徙数量相近,这可能与路氹湿地在候鸟迁徙途中的主要作用为非能量补给站有关(主要作为越冬地),但具体原因需要更深入细致的研究(蒋科毅等,2011)。
谱系多样性和平均成对谱系距离的季节变化与物种丰富度相似:即秋、冬季较高,这与物种数越多,增加群落中亲缘关系较远的物种概率越大,因此导致PD和MPD越高(Webbetal.,2002)。然而,夏季PD的下降程度明显高于物种丰富度,这是因为:生态一区夏季鸟类群落以鹭科为主(占比为66.67%),生态二区以鹭科和鹬科为主(占比分别为46.67%和20.00%),主要由亲缘关系很近的物种组成,故PD(谱系树的枝长之和)很小,同时也说明了PD的下降主要由某些谱系分支的消失引起,而不是谱系中一些物种的消失引起。
生态一区夏季和二区春、夏季水鸟呈现出显著的谱系聚集,这与Che等(2019)对香港特别行政区米埔湿地水鸟的研究结果一致,表明了环境过滤对夏季水鸟群落构建具有重要作用。在繁殖季节(春、夏季),由于需要寻找配偶和巢址,水鸟对栖息地有更加严格的选择性(Sagario & Cueto,2014),因此环境因子起着重要作用:生态二区是澳门地区最大的鹭鸟觅食地和繁殖地,其大多在3月开始繁殖,持续至8月(主要繁殖地),而生态一区夏季仅有少量鹭鸟繁殖(次要繁殖地)。此外,生态一区春、秋和冬季,以及生态二区秋季水鸟皆趋向于谱系发散,可能表明种间竞争在群落构建中作用更大:在这些季节,水鸟的分布主要受到食物丰富度和分布的影响,其相似的食物需求和有限的斑块面积可能导致了群落存在较强的种间竞争。杨月伟等(2005)认为,有限的斑块面积增加了水鸟种间竞争;迁徙水鸟的大量涌入导致了资源竞争(Hamdietal.,2008; Cheetal.,2019)。特别地,生态二区冬季水鸟趋向于谱系随机,这可能与多种因素有关:食物的可获得性可能会驱动随机过程(Ramos-Roblesetal.,2016),此外,相反作用的群落构建机制可能互为抵消,导致明显的随机过程(Stegenetal.,2013)。
虽然鸟类谱系结构在推断群落聚群规律时具有较强的敏感性,但我们对这些结论需保持谨慎态度。有学者指出,谱系聚集并不常常反映了环境过滤的影响(如HilleRisLambersetal.,2012;Gerholdetal.,2015),这需要更多研究案例验证。
致谢:广东省生物资源应用研究所动物调查监测与恢复中心的李秋剑副研究员参与了大量野外工作,在此表示感谢。
