安吉白茶白化叶和返绿叶叶绿体超微结构和叶绿素及其前体物含量的比较
2019-09-10杨荧吕立堂赵德刚
杨荧 吕立堂 赵德刚


摘要:安吉白茶(Camellia sinensis 'Baiye1')是典型的白化茶树品种,为了对其叶片白化现象进行探究,本研究观察了安吉白茶的白化叶和返绿叶的叶绿体超微结构,发现白化葉叶绿体内部结构发育不良,基粒片层排列疏松散乱,片层间缝隙大,层次不清晰,甚至断裂不成形,根据对叶绿素及其前体物质含量的测定和对叶绿素合成的相关基因在两个阶段叶片中的表达量测定结果发现,Mg-proto Ⅸ在白化叶中含量要高于返绿叶,推测叶绿素合成阶段的Mg-proto Ⅸ合成Pchlide a时受阻,确定安吉白茶发生白化是由于叶绿体发育不良,同时叶绿素合成阶段的Mg-proto Ⅸ合成Pchlide a时受阻所致。
关键词:安吉白茶;叶绿体;超微结构;叶绿素前体物
中图分类号:S5711
文献标识码:A
文章编号:1008-0457(2019)06-0071-05国际DOI编码:10.15958/j.cnki.sdnyswxb.2019.06.013
Comparison of Ultrastructure, Chlorophyll and Precursor Contents of Chloroplasts between Albino Leaves and Green Leaves in Camellia sinensis ‘Baiyel 1’
YANG Ying1,LV Li-tang2*,ZHAO De-gang1,3
(1.College of Tea / Research Institute of Agricultural Bioengineering, Guizhou University, key Laboratory of Germplasm Innovation for Conservation and Protection of Mountain Plant Resources, Guiyang, Guizhou 550025,China;2.Guizhou Academy of Agricultural Sciences, Guiyang,Guizhou 550006,China)
Abstract:Camellia sinensis cv. Baiye1 is a typical albino tea cultivar. In order to investigate the albinism phenomenon of its leaves, the chloroplast ultrastructure of the albino leaves and the green leaves of Camellia sinensis cv. Baiye1 were observed, and it was found that the internal structure of the chloroplasts in the albino leaves was stunted. The grana lamellae were scattered and loosely arranged with big gaps, and the lamellar structure was not clear, even fractured. According to the determination of the content of chlorophyll and its precursors, and the expression of genes related to chlorophyll synthesis in leaves of two stages , it was found that the content of Mg-proto IX in albino leaves was significantly higher than that in returning green leaves and speculated that the synthesis of Pchlide a from Mg-proto IX was blocked in the chlorophyll synthesis pathway. These results indicate that the albinism of Camellia sinensis cv. Baiye1 was due to chloroplast dysplasiaand the blocking synthesis of Pchlide a from Mg-proto IX.
Key words:Camellia sinensis cv. Baiyel; chloroplast; ultrastructure; chlorophyll precursor
茶汤中含有的茶多酚、氨基丁酸、脂多糖,微量元素锰、锌、硒,茶坩宁等具保健作用,以茶树白化突变体为原料制成的绿茶滋味鲜爽,香气、外形、汤色、叶底等品质优秀,提高了其经济价值[1],同时由于叶绿素含量降低,氨基酸含量明显增高,使茶树白化突变体制成的茶具有更高的营养价值[2],植物的叶色突变与叶绿素的合成及叶绿体的发育息息相关[3-4],安吉白茶(Camellia sinensis 'Baiye1')作为典型的白化茶品种,经研究发现返白过程中氨基酸升高,叶绿素含量则下降[5]。本研究以安吉白茶为研究材料,对其白化叶和返绿叶叶绿体超微结构和叶绿素及其前体物含量进行比较和研究,以揭示安吉白茶白化的原因。
1材料与方法
11材料
安吉白茶(Camellia sinensis ‘Baiye1’)的白化叶及返绿叶作为研究材料,采样于中国贵州六盘水市六枝特区黑晒村茶园(由四川引种),白化叶和返绿叶均处于相同的叶片发育阶段,由本实验室保存。
12试剂
植物RNA提取试剂盒(Plant RNA Kit)、Premix TaqTM Version 20 plus dye、DL1000 DNA Marker5、Ehrlich-Hg試剂,三氯乙酸、乙酰丙酮、盐酸、丙酮、正己烷、氨水、醋酸钠、磷酸缓冲液试剂(NaH2PO4&Na2HPO4)、冰醋酸、乙醚、戊二醛等(若无说明均为国产或进口分析纯)。
13主要仪器
分光光度计(BioPhotoMeter),PCR仪(Bio-rad),凝胶成像系统(XRS+),分析天平BS224S(Sartorius),电泳仪(Bio-rad),精密pH计pHS-2C(上海虹益),超纯水仪(MicroPure),台式离心机X1R(Thermo),超净工作台(上海上净净化),制冰机F100 Compact(Lcematic,高压蒸汽灭菌锅MLS-3750(SANYO),移液枪(Eppendorf)、分液漏斗、涡旋振荡器SK-1(上海梅香)、数码相机1000D(佳能)等。
14实验方法
141电镜观察方法
为了确定安吉白茶白化过程中其叶绿体是否结构完整(存在或解体),对安吉白茶白化叶和返绿叶的细胞进行超微结构观察,安吉白茶叶片样本的制片和观察方法均参照杨勇骥等人[6]的方法。
142叶绿素前体物质含量测定
为确定安吉白茶白化过程中是否存在叶绿素合成受阻现象,对叶绿素、类胡萝卜素和叶绿素合成过程中主要的叶绿素前体物质的含量进行测定,其中δ-氨基乙酰丙酸(ALA)的测定参照Dei等人[7]的方法,胆色素原(PGB)的测定参照Bogorad等人[8]的方法,原卟啉IX protoporphyrin IX(ProtoIX:)、镁原卟啉IX Mg-protoporphyrin IX(Mg- Proto IX)和原脱植基叶绿素(Pchlide)的相对含量测定参照Rebeiz和Lee等人[9-10]的方法。叶绿素和类胡萝卜素的含量参照张治安等人[11]的方法。
143叶绿素合成相关基因表达量的测定
为对白化过程中叶绿素合成过程进行进一步的分析,将白化叶和返绿叶的叶绿素合成相关基因进行表达量的测定,参照Omega公司E.Z.N.A.Plant RNA Kits操作指南进行安吉白茶叶片RNA的提取。
cDNA合成参照TaKaRa Reverse Transcriptase M-MLV说明书进行,使用RTase M-MLV(RNase H-)反转录酶将提取的安吉白茶总RNA反转录为cDNA。
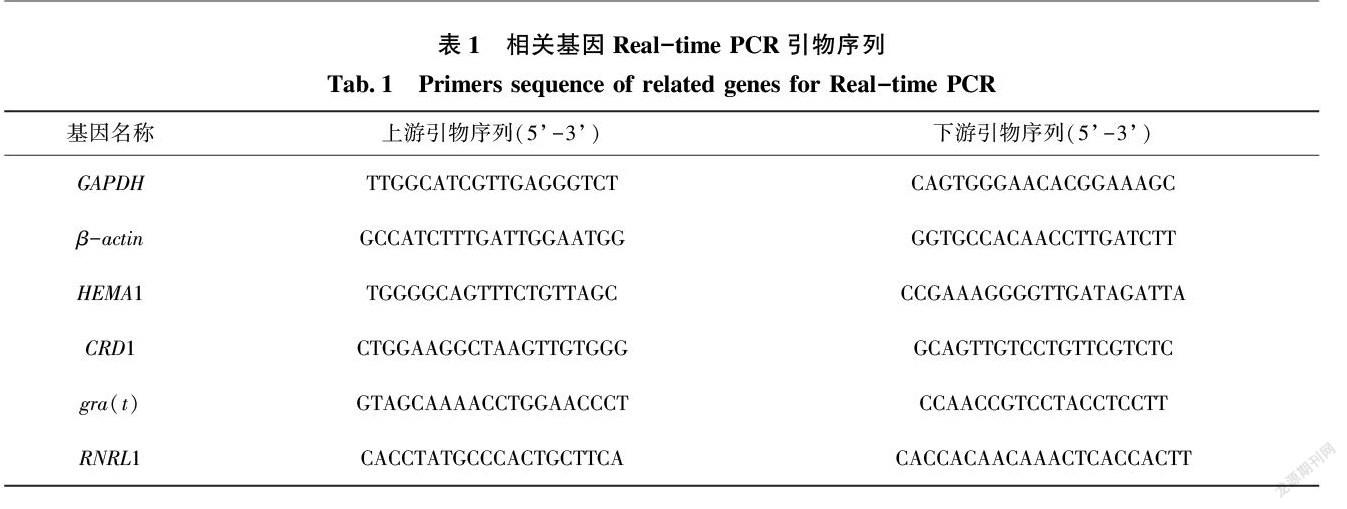
用Primer Premier 50软件,根据定量PCR引物的设计原则,设计以下基因引物:白化转绿基因gra(t),核糖核酸还原酶大亚基基因(RNRL1),谷氨酰-tRNA还原酶基因(HEMA1),镁原卟啉原 IX 单甲酯环化酶基因(CRD1)设计各基因的RT-PCR扩增引物(表1)。以茶树GAPDH、β-actin基因(actin)作为内参,定量分析相关基因的表达情况。参照ABI公司的操作指南,使用SYBR Green染料进行实时定量PCR分析,数据分析方法为ΔΔCT法。
144数据统计分析方法
采用Excel 2016和SPSS 23进行图像绘制、统计分析、方差分析和多重比较,结果以x±s表示,文中图表的表示方式均为平均值,文中所有数据均是3次重复试验的平均值,所有图也为3次重复试验的平均值,误差线表示3次重复试验的标准偏差。
2结果和分析
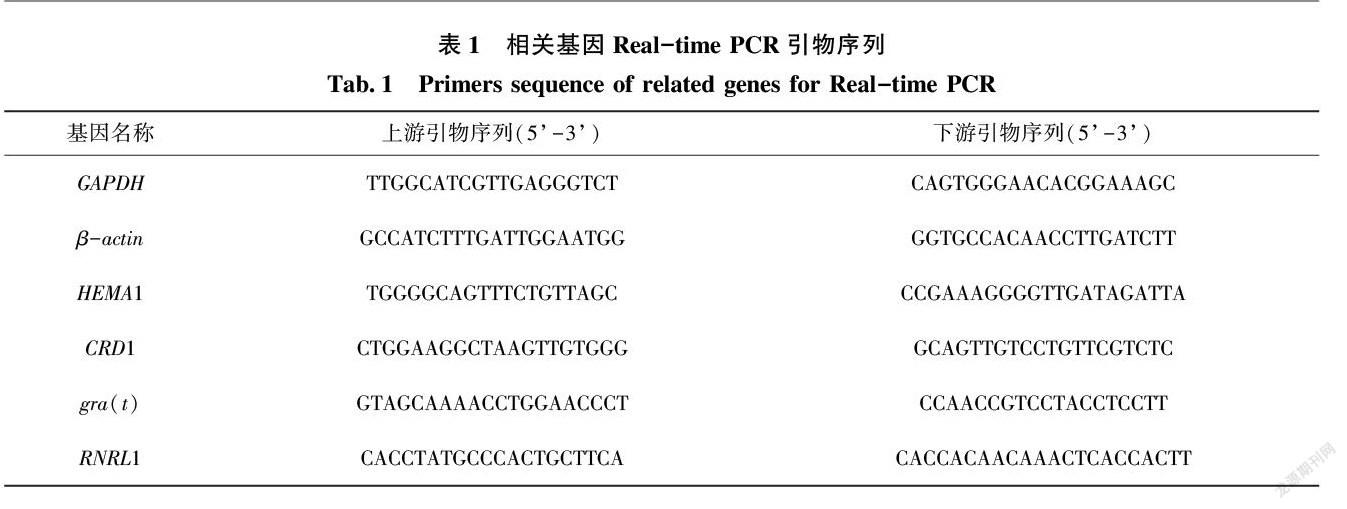
21安吉白茶白化叶和返绿叶的叶色观察
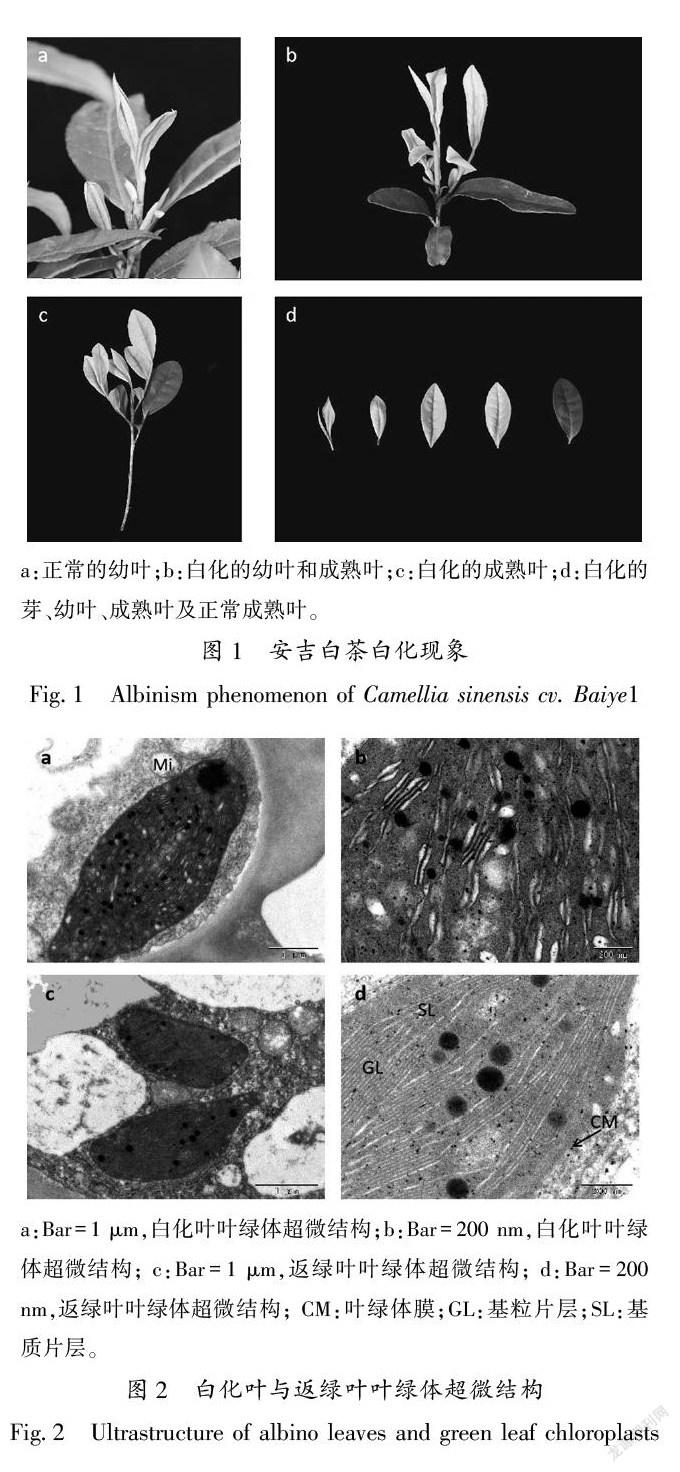
对安吉白茶白化阶段的表型观察,如图1所示,安吉白茶白化性状(图1-b、c、d)基本出现在芽到芽下三到四叶(图1中白化的芽、白化的幼叶和白化的成熟叶),白化性状明显(图1-b、c、d),而完全成熟的老叶(图1中芽下三、四叶以后的老叶)不会出现白化性状(图1- b、c、d)。
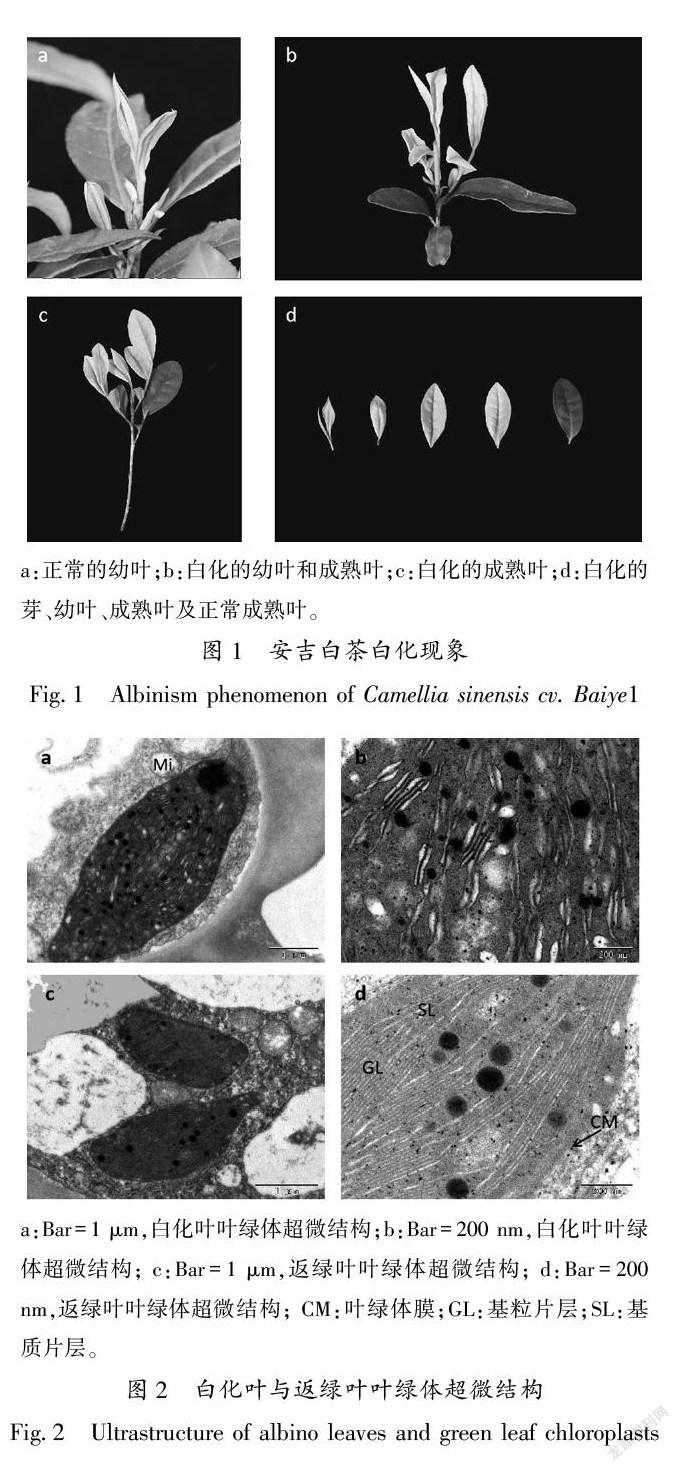
22安吉白茶白化叶和返绿叶叶绿体超微结构的观察
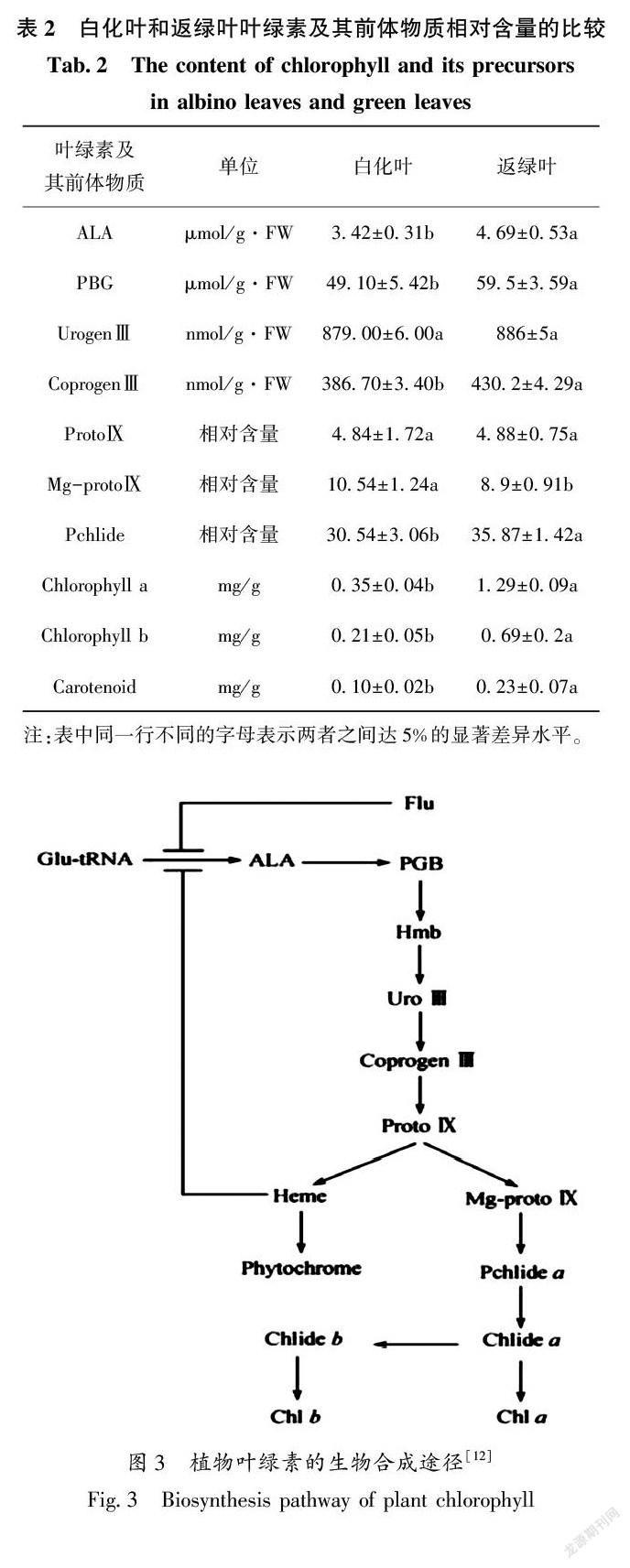
将安吉白茶的白化叶和返绿叶样本制片,观察叶绿体的超微结构,发现在白化叶的细胞中叶绿体内部结构发育不良,基粒片层排列疏松散乱,片层间缝隙大,层次不清晰,甚至断裂不成形(图2-a、b);返绿叶中的叶绿体结构良好,形状多为长椭圆形或梭形,叶绿体被膜完整,片层结构垛叠整齐,基粒片层和基质片层均排列紧致且清晰(图2-c、d)。从图2可看出,安吉白茶的白化叶由于其叶绿体发育不良,使安吉白茶的叶色呈现出白化的性状(图1)。
23安吉白茶白化叶和返绿叶叶绿素及其前体物质含量的测定
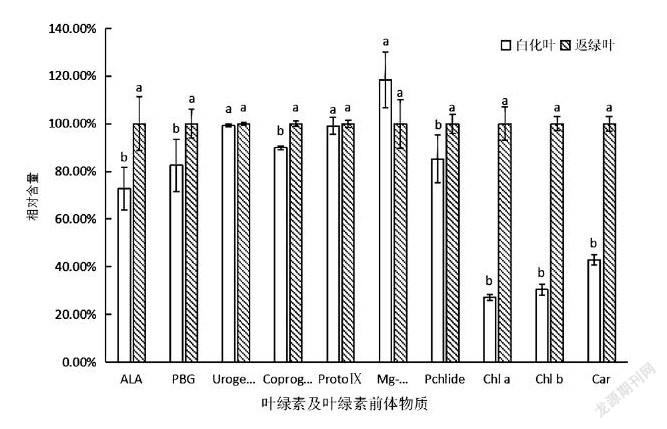
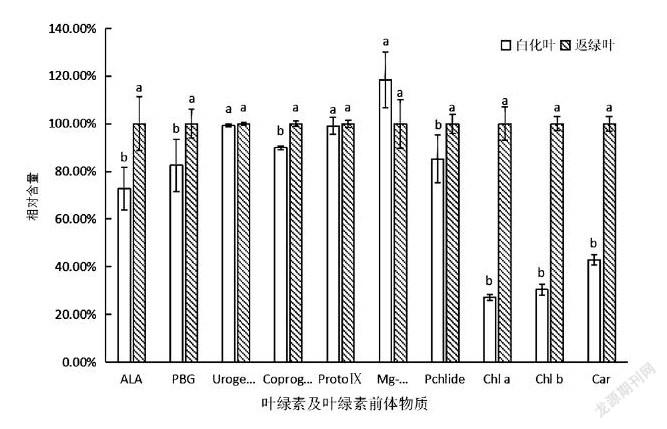
为确定安吉白茶白化过程中叶绿素合成过程[12]受阻的具体阶段,本试验测定了安吉白茶的白化叶和返绿叶中主要的几种叶绿素、类胡萝卜素及叶绿素前体物质的含量,将白化叶和返绿叶的叶绿素及其前体物质的含量进行方差分析和多重比较(表2)。从表2可以看出,δ-氨基乙酰丙酸(ALA)、胆色素原(PGB)、粪卟啉原Ⅲ(CoprogenⅢ),原脱植基叶绿素a(Pchlide a)、叶绿素a(Chlorophyll a)、叶绿素b(Chlorophyll b)和类胡萝卜素(Carotenoid)在返绿叶中的含量均高于白化叶,在Mg-proto Ⅸ之前的原卟啉Ⅸ(ProtoⅨ)的含量在两种叶片中的差异并不显著(P>005),但由于Pchlide a合成受阻,导致之后的各前体物质含量在两种叶片中都差异显著(P<005)(表2)。
将返绿叶测得的各物质含量定为100%,并与白化叶测得的含量进行比较(图4),其中ALA、PGB、CoprogenⅢ、ProtoⅨ和 Pchlide a合成的各前体物质含量均为返绿叶高于白化叶,或是差异不显著(P>005),但Mg-原卟啉Ⅸ(Mg-proto Ⅸ)的含量却是白化叶高于返绿叶,并且差异显著(P<005)。从图4可以发现,这是由于Mg-proto Ⅸ合成Pchlide a时受阻,导致Mg-proto Ⅸ积累,因此,白化叶中的Mg-proto Ⅸ含量高于返绿叶。
24与叶绿素合成相关基因在安吉白茶白化叶和返绿叶中的表达量
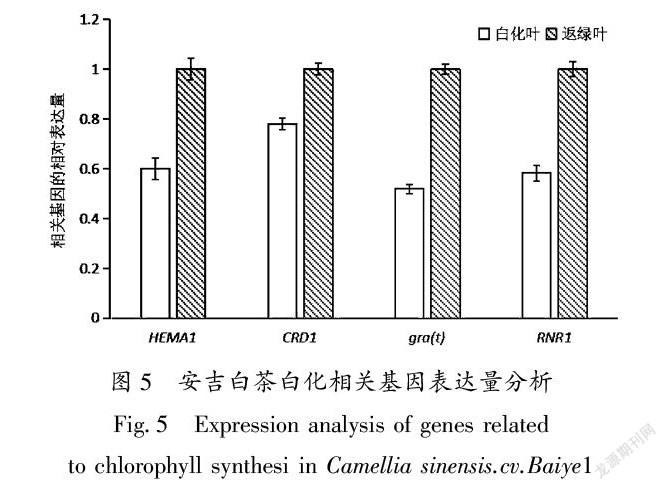
对安吉白茶的白化叶和返绿叶进行Real time-PCR分析结果如图5:谷氨酰tRNA还原酶1基因(HEMA1)在白化叶中的表达量是返绿叶中的060倍,白化转绿基因(gra(t))在白化叶中的表达量是返绿叶中的052倍,核糖核酸还原酶大亚基基因(RNRL1)在白化叶中的表达量是返绿叶中的058倍,镁原卟啉原 IX 单甲酯环化酶基因(CRD1)在白化叶中的表达量是返绿叶中的077倍。以上各基因表达差异较大,HEMA1、gra(t)、RNRL1在白化叶中的表达量均比返绿叶中的表达量低。
镁原卟啉原 IX 单甲酯环化酶基因(CRD1)编码的蛋白主要功能是催化Mg-proto Ⅸ合成Pchlide a,CRD1在白化叶中表达量低,与本实验室对白化叶和返绿叶叶绿素及其前体物质含量的测定结果相一致。其次,与叶绿素合成相关的基因HEMA1、gra(t)、RNRL1在白化叶中的表达量均低于返绿叶,使白化叶中叶绿素含量低于返绿叶。
3结论与讨论
叶绿体的发育异常和叶绿素的合成受阻都会导致植物发生叶色突变[13-14],本研究通过对白化叶叶绿体超微结构的观察发现,白化叶叶绿体基粒片层排列疏松散乱甚至断裂不成形,只有些许类囊体残留(图2),推测白化叶的叶绿体是在其发育阶段中的基粒構建阶段受到影响,其叶绿体发育受到影响而导致安吉白茶叶白化。同时在对两种叶片叶绿素及其前体物含量测定(表2、图4)时发现,白化叶中Mg-原卟啉Ⅸ(Mg-proto Ⅸ)含量高于返绿叶,推测是因为叶绿素合成阶段中Mg-proto Ⅸ合成Pchlide a时受阻,使Mg-proto Ⅸ在白化叶片中积累[15],使白化叶中Mg-proto Ⅸ含量高于返绿叶。对白化叶和返绿叶中与叶绿素合成的相关基因表达量测定时发现,编码催化Mg-proto Ⅸ合成Pchlide a的镁原卟啉原 IX 单甲酯环化酶[16]的基因(CRD1)在白化叶中的表达量要低于返绿叶(图5),其他与叶绿素合成相关的基因(HEMA1、gra(t)、RNRL1)[17]在白化叶中的表达量均低于返绿叶(图5),使安吉白茶在白化过程中的叶绿素合成减少,叶片发生白化。对类胡萝卜素含量的测定发现,白化叶中类胡萝卜素的含量低于返绿叶,叶绿素没有类胡萝卜素的保护,也会使叶绿素含量进一步降低[18](表2、图4)。
通过上述对安吉白茶两个阶段的叶绿体超微结构的观察和对叶绿素及其前体物质含量的测定发现,安吉白茶在白化期叶绿体结构异常,同时叶绿素合成受阻,本研究推测是由于在叶绿素合成阶段Mg-proto Ⅸ合成Pchlide a时受阻及叶绿体发育不良导致安吉白茶在白化期白化。
参考文献:
[1]陈春梅,马春雷,马建强,等. 茶树cpDNA测序及基于cpDNA序列的山茶属植物亲缘关系研究 [J].茶叶科学,2014,4):371-380
[2]邵济波,唐茜,周晓兰,等. 四川引种安吉白茶主要生化成分分析 [J].食品科学,2012,33(16):179-183
[3]涂美艳,宋海岩,陈栋,等. 川中丘陵区碱性土对GF677和毛桃叶片光合特性及叶绿素荧光参数的影响 [J].山地学报,2018,36(1):153-162
[4]黄小贞,赵懿琛拟南芥PGK基因家族功能的初步分析 [J].山地农业生物学报,2017,36(1):12-17
[5]于淑池,刘畅,王乘慧,等. 安吉白茶茶多酚的抗氧化活性研究 [J].时珍国医国药,2012,23(5):1184-1187
[6]杨勇骥医学生物电子显微镜技术[M].上海:第二军医大学出版社,2012
[7]DEI MBenzyladenine-induced stimulation of two components of chlorophyII formation in etiolated cucumber cotyledons [J].Physiologia Plantarum,2010,62(4):521-526
[8]Bogorad, L. [Methods in Enzymology], Volume 5 || [122] Porphyrin synthesis[J]. Methods in Enzymology,1962,5:885-895.
[9]REBEIZ C Z,SMITH B B,MATTHEIS J R,et al Chloroplast biogenesis Biosynthesis and accumulation of Mg-protoporphyrin IX monoester and other metalloporphyrins by isolated etioplasts and developing chloroplasts [J].Archives of Biochemistry & Biophysics,1975,167(1):351-365
[10]LEE H J,BALL M D,PARHAM R,et al Chloroplast Biogenesis 65:Enzymic Conversion of Protoporphyrin IX to Mg-Protoporphyrin IX in a Subplastidic Membrane Fraction of Cucumber Etiochloroplasts [J].Plant Physiology,1992,99(3):1134-1140
[11]张治安,张美善,蔚荣海 植物生理学实验指导[M].北京:中国农业科学技术出版社,2004
[12]史典义,刘忠香,金危危 植物叶绿素合成、分解代谢及信号调控 [J].遗传,2009,31(7):698-704
[13]CHEN T,ZHANG Y,ZHAO L,et al Fine mapping and candidate gene analysis of a green-revertible albino gene gra(t)in rice [J].Journal of Genetics & Genomics,2009,36(2):117-123
[14]LI C,ZHENG Y,ZHOU J,et al Changes of leaf antioxidant system,photosynthesis and ultrastructure in tea plant under the stress of fluorine [J].Biologia Plantarum,2011,55(3):563-566
[15]吕典华 水稻叶色突变体的光合和生理生化特性研究 [D]; 重庆:西南大学,2010
