H2S作为植物个体间交流的气体信号分子
2019-07-23刘志强曹纯玉李亚文渠娟娟贾云乾裴雁曦
刘志强 曹纯玉 李亚文 渠娟娟 贾云乾 裴雁曦
(山西大学生命科学学院,特色植物资源研究与利用山西省重点实验室,太原 030006)
交流是一种重要的生存能力,人与人之间、动物与动物之间,以及细胞与细胞之间都存在着独特的交流方式,那么植物与植物之间呢?它们是否也在“窃窃私语”?答案是肯定的。

自由跨膜是气体信号分子的特征之一[21]。生理浓度下的外源H2S处理可以提高植物对多种胁迫的抗性;并且胁迫会引起植物内源H2S含量的升高[22~23],那么当植株受到环境胁迫时,细胞内的H2S是否会释放到空气中,成为植株间的报警信号,在植株间起到通讯的作用?本论文对此进行了研究。
1 材料与方法
1.1 植物材料的培养
本实验所用植物材料包括:拟南芥(Col-0、lcd/des1突变体)、谷子(晋谷21号)、白菜(津育75号)、番茄(MicroTom)。选择饱满的种子,4℃春化48 h,种植在营养土和蛭石(2∶1)的混合基质中,生长4周。培养条件为:温度23±1℃,60%相对湿度,昼/夜周期16/8 h,光照强度为3 000 lx。
1.2 植物叶片H2S含量的测定
参考Li等的方法[24],0.1 g叶片用2 mL提取液(50 mmol·L-1磷酸缓冲液pH6.8,0.2 mol·L-1抗坏血酸和0.1 mol·L-1EDTA)研磨;将匀浆液转移到可密闭的小瓶中,放置含0.5 mL 1% Zn(Ac)2溶液的吸收小管,向匀浆中加入1 mL HCl(1 mol·L-1),密封小瓶;于室温反应30 min,向吸收管加入DPD和FeCl3溶液,混匀避光放置15 min,测溶液667 nm的吸光值,根据标准曲线得出H2S浓度后,再根据公式计算H2S含量。
H2S含量(μmol·g-1FW)=H2S浓度×(0.5+0.2+0.2)×0.001/鲜重
(1)
1.3 空气中H2S浓度的检测
对照组和PEG 8000处理组植物材料分别置于玻璃缸内(7 L),放置含0.5 mL 1% Zn(Ac)2溶液的EP管,用于吸收缸内空气中的H2S气体,12 h后取出EP管,分别加200 μL DPD(Dimethyl-p-phenylenediamine,20 mmol·L-1)和FeCl3(30 mmol·L-1)溶液,混匀避光放置15 min,测溶液667 nm的吸光值,根据标准曲线计算空气中H2S浓度,为0 h数据。向处理组植物材料的土壤中加20 mL 40% PEG 8000,放置Zn(Ac)2溶液,12 h后检测。之后每12 h放置新的Zn(Ac)2溶液,分别为胁迫24、36、48 h样品。对照组不进行胁迫处理,置于玻璃缸,每12 h放置新的Zn(Ac)2溶液。
1.4 PEG 8000处理植株对邻近植物H2S含量和基因表达的影响
谷子幼苗分为A、B两组,分别置于7 L的玻璃缸内。A组为对照(Control),缸内植株不进行胁迫处理;B组缸内植株分为PEG 8000处理植株和非处理(un-PEG 8000)植株;处理时向处理植株的土壤加入20 mL 40% PEG 8000,处理时间为12 h。检测Control、PEG 8000、un-PEG 8000三种条件下谷子叶片H2S含量。白菜、番茄和拟南芥Col-0幼苗分别按上述进行处理后检测叶片H2S含量,PEG 8000的处理时间为24 h。
谷子、白菜、番茄和拟南芥Col-0幼苗分为A、B两组,每组含四种植物。分别置于7 L的玻璃缸内。A组缸内植株不进行胁迫处理(Control);B组缸内的部分谷子幼苗进行40% PEG 8000处理12 h;检测A组和B组缸内非胁迫处理的(un-PEG)谷子、白菜、番茄和拟南芥Col-0幼苗叶片H2S含量。检测A组缸内谷子、白菜和拟南芥Col-0和B组缸内非胁迫处理(un-PEG)的谷子、白菜和拟南芥Col-0幼苗叶片中H2S响应基因的表达变化。
1.5 基因表达检测
提取植株叶片总RNA,按abm公司试剂盒(Cat.No.G492)进行反转录得到cDNA,以各植物的Actin基因作为内参,利用qRT-PCR检测基因转录水平变化,结果以2-δδCt计算,所用引物见表1。
1.6 气孔观察
拟南芥野生型Col-0和突变体lcd/des1分成三组:对照组、PEG 8000处理的Col-0及处于同一玻璃缸内的不处理Col-0和lcd/des1、PEG 8000处理的lcd/des1及处于同一玻璃缸内的不处理Col-0和lcd/des1。PEG 8000处理12 h,显微镜观察和记录玻璃缸内叶片气孔孔径。
1.7 统计分析
使用SPSS Statistics软件对3次生物学重复的实验数据进行分析,用“平均值±标准偏差”表示。
2 实验结果
2.1 PEG 8000处理植物提高周围环境空气中H2S含量
可自由通过细胞膜是气体信号分子的特征之一。生理浓度的外源H2S处理提高植物对干旱、盐等逆境胁迫的抗性[25~26],即外源的H2S气体可以进入细胞发挥其生理功能。H2S气体信号分子参与植物对多种非生物胁迫的响应,即胁迫引起植物内源H2S含量升高,那么细胞内的H2S是否会向空气中释放?40% PEG 8000处理拟南芥,谷子,白菜和番茄植株,每12 h检测空气中H2S浓度,结果表明:胁迫处理谷子12、36 h,处理白菜24、36 h,处理番茄24、36、48 h,处理拟南芥Col-0野生型12、24、36 h后,都检测到空气中的H2S含量升高(图1A);而拟南芥内源H2S产生酶基因突变体lcd/des1在40% PEG 8000处理后,没有引起空气中H2S浓度升高(图1A)。胁迫引起空气中H2S浓度升高,因此对邻近的非胁迫植株的内源H2S含量进行检测。结果表明:受胁迫植株(图1B,PEG)和处于同一个玻璃缸中的非胁迫植株叶片的H2S含量(Fig.1B,un-PEG)都高于对照(图1B,CK),即受胁迫植株释放的H2S可引起邻近的同物种植株内源H2S含量升高。

表1 qRT-PCR所用的引物列表
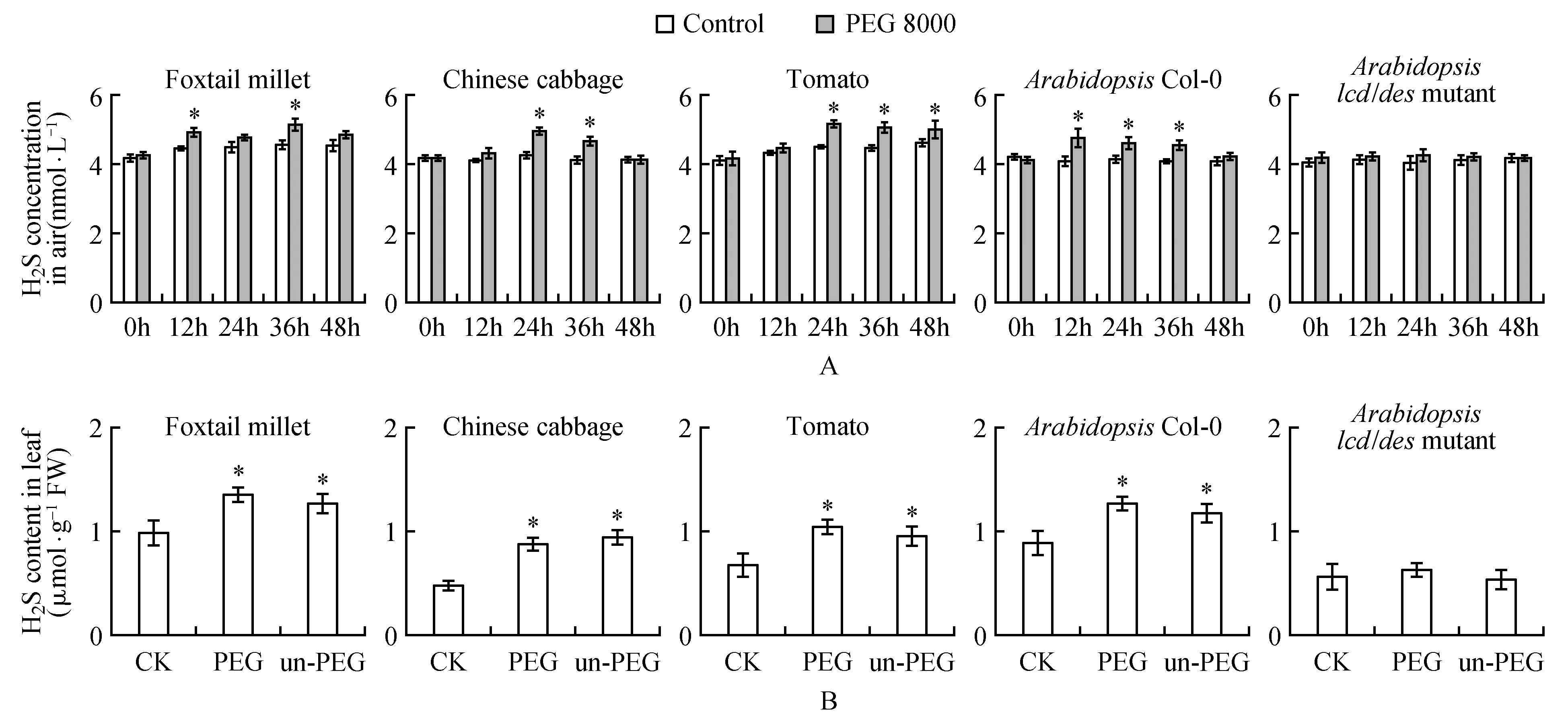
图1 PEG 8000处理植物对空气中H2S浓度和邻近非胁迫植株内源H2S含量的影响 A.纵坐标表示空气中H2S浓度,横坐标表示PEG 8000处理时间;B.纵坐标表示植物叶片内源H2S含量(CK.未处理植株;PEG. 40% PEG 8000处理12 h;un-PEG.与PEG 8000处理植株处于同一玻璃缸内的未处理植株) *表示在P<0.05水平存在显著性差异,Duncan test(下同)。Fig.1 Effect of PEG 8000 on the concentration of H2S in air and endogenous H2S content in nearby un-treated plantA.The ordinate indicates the H2S concentration in the air,and the abscissa indicates the PEG 8000 treatment time; B.The ordinate indicates the endogenous H2S content of the plant leaves(CK. Untreated plants; PEG. 40% PEG 8000 for 12 h; un-PEG. Untreated plants in the same glass jar as the PEG 8000 treated plants) *indicates a significant difference at the P< 0.05 level,Duncan test(the same as below).
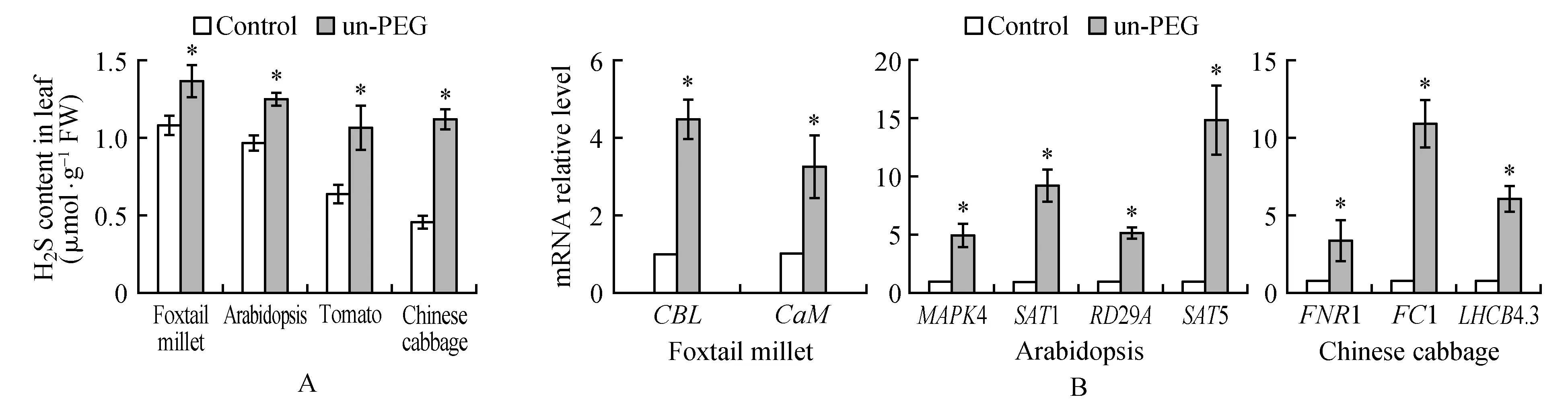
图2 PEG 8000处理谷子对邻近非胁迫植物内源H2S含量和基因表达的影响 A.纵坐标表示植物叶片H2S含量;B.纵坐标表示H2S响应基因的相对表达量;Control.与未处理谷子处于同一玻璃缸内的未处理植株;un-PEG.与PEG 8000处理谷子处于同一玻璃缸内的未处理植株Fig.2 PEG 8000 treated foxtail millet effects H2S content and gene expression of adjacent non-stressed plants A. The ordinate indicates the H2S content of the plant leaves; B. The ordinate indicates the relative expression of the H2S response gene Control. The untreated plants in the same glass jar as the untreated millet; un-PEG. The untreated plants in the same glass jar as the PEG 8000 treated millet
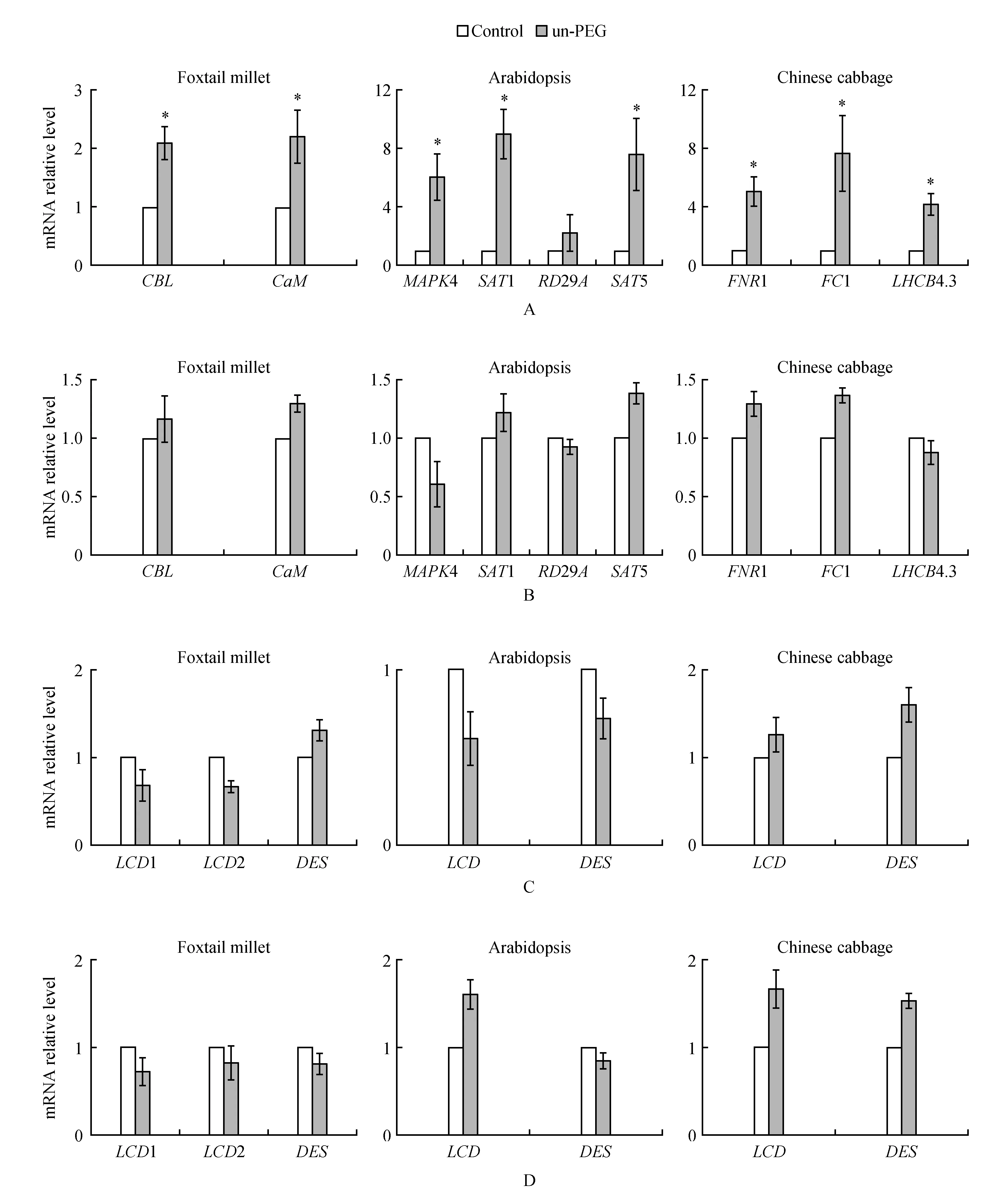
图3 PEG 8000处理拟南芥对邻近非胁迫植物基因表达的影响 A. PEG 8000处理拟南芥Col-0 12 h,同一玻璃缸内非胁迫植物H2S响应基因相对表达变化;B. PEG 8000处理拟南芥lcd/des1 12 h,同一玻璃缸内非胁迫植物H2S响应基因相对表达变化;C. PEG 8000处理拟南芥Col-0 12 h,同一玻璃缸内非胁迫植物H2S生成酶基因相对表达变化;D. PEG 8000处理拟南芥lcd/des1 12 h,同一玻璃缸内非胁迫植物H2S生成酶基因相对表达变化Fig.3 The effect of PEG 8000 treated Arabidopsis on gene expression of adjacent non-stressed plants A. PEG 8000 treatment of Arabidopsis Col-0 12 h, relative expression of H2S response genes in non-stressed plants in the same glass jar; B. PEG 8000 treatment of Arabidopsis lcd/des1 12 h, H2S response gene in non-stress plants in the same glass jar Relative expression changes; C. PEG 8000 treatment of Arabidopsis Col-0 12 h, relative expression of LCD and DES in non-stressed plants in the same glass jar; D. PEG 8000 treatment of Arabidopsis lcd/des1 12 h, relative expression of LCD and DES in non-stressed plants in the same glass jar
2.2 PEG 8000处理谷子对邻近其它非胁迫植物的H2S含量和基因表达的影响
谷子幼苗经PEG 8000处理12 h,检测同一玻璃缸内未处理的谷子、拟南芥(Col-0)、番茄和白菜叶片的H2S含量和H2S响应基因表达的影响。结果表明,与未经PEG 8000处理谷子缸内的谷子、拟南芥(Col-0)、番茄和白菜相比(图2A,Control),经40% PEG 8000处理12 h谷子缸内的非PEG 8000处理的谷子、拟南芥(Col-0)、番茄和白菜叶片的H2S含量升高(图2A,un-PEG)。H2S信号分子可以调节基因表达参与植物对胁迫的响应,选取已经报道响应H2S的谷子CBL,CaM[27],拟南芥MPK4,SAT1,RD29A,SAT5[28~29]和白菜FNR1,FC1,LHCB4.3[30]基因进行检测(图2B),与放置在未经PEG 8000处理谷子相同缸内的植物相比(图B,Control),放置在40% PEG 8000处理谷子同一玻璃缸内的非处理的谷子、拟南芥(Col-0)和白菜叶片的上述基因表达都升高(图2B,un-PEG)。
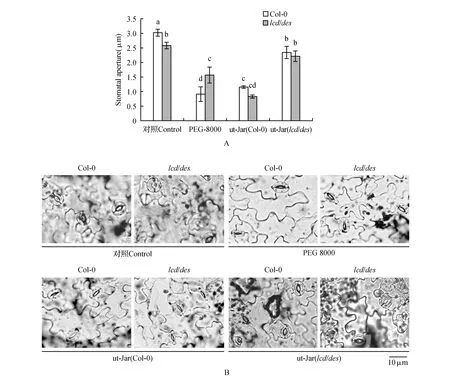
图4 PEG 8000处理拟南芥对邻近非胁迫拟南芥气孔运动的影响 A.纵坐标表示气孔孔径(PEG-8000. 40% PEG 8000处理12 h植株;ut-Jar(Col-0).与PEG 8000处理的Col-0处于同一玻璃缸内的未处理植株;ut-Jar(lcd/des1).与PEG 8000处理的lcd/des1处于同一玻璃缸内的未处理植株 不同字母表示显著性差异,Duncan test,P<0.05);B.气孔显微镜观察Fig.4 The effect of PEG 8000 treated Arabidopsis on stomatal aperture of adjacent non-stressed Arabidopsis A. Ordinate indicates stomatal aperture(PEG-8000. 40% PEG 8000 treated 12 h plant; ut-Jar(Col-0). Untreated plant in the same glass jar as PEG 8000 treated Col-0; ut-Jar(lcd/des1). Untreated plants in the same glass jar as PEG 8000 treated lcd/des1 With different letters indicating significant differences,Duncan test,P<0.05); B.Observation by a stomatal microscope
2.3 PEG 8000处理拟南芥对邻近非胁迫植物基因表达和气孔运动的影响
PEG 8000处理谷子引起邻近植物的内源H2S含量和H2S响应基因表达变化,利用拟南芥野生型Col-0和H2S产生酶基因LCD和DES1双突变体lcd/des1对该结果进行进一步验证。PEG 8000处理Col-0和突变体lcd/des1植株12 h后,检测未处理植物的基因表达变化(图3A),处于Col-0玻璃缸内的未处理白菜,拟南芥(Col-0)和谷子中受H2S调控的基因均显著升高(图3A),而处于拟南芥lcd/des1相同玻璃缸内的未处理植株基因表达变化并不明显(图3B)。为了了解邻近植物内源H2S含量升高的原因,检测其主要的H2S产生酶基因LCD、DES的表达变化。结果表明,放置在PEG 8000处理的拟南芥Col-0或lcd/des1玻璃缸内的未处理植物的LCD、DES基因表达均无显著性变化(图3C,D)。
H2S信号分子促进气孔关闭,参与植物对干旱胁迫的响应[25],对上述处理条件下的拟南芥Col-0和lcd/des1气孔孔径进行观察。结果表明,PEG 8000处理12 h的Col-0气孔孔径明显小于对照Col-0,也可导致同一玻璃缸内未处理的Col-0和lcd/des1气孔孔径减小;PEG 8000处理12 h的lcd/des1气孔孔径小于对照lcd/des1,但不能使同一玻璃缸内未处理的Col-0和lcd/des1气孔孔径减小(图4)。
3 讨论
植物个体间通过挥发性物质传递信息。1983年,Baldwin等报道叶片的机械损伤可导致附近的未受损树木的酚类和单宁等升高[31]。Dudareva等将已发现的植物挥发性物质分为四类:萜类化合物、脂肪酸衍生物、苯环及苯丙烷衍生物和氨基酸衍生物[32],这些物质多数属于次级代谢物,可作为信号分子、抗氧化剂等发挥多种生理或生态功能。除了植物,这些挥发性物质在动物,细菌及真菌中同样存在。近十几年来,NO、CO和H2S等气体信号分子的相关研究大量报道,集中于这些分子在细胞内产生的调节,外源供体处理提高细胞或个体对胁迫的抵抗等机制研究。植物体细胞内的H2S主要由酶催化产生,可被低温、干旱等胁迫诱导升高,参与后续的胁迫抵抗和耐受反应。本研究发现,PEG 8000胁迫可引起植物向周围环境中释放H2S,使邻近非胁迫植株的内源H2S含量升高(图1),通过对邻近植株中H2S合成酶基因表达变化的检测,发现这些植物中的H2S合成酶基因表达没有显著变化,所以邻近植株内源H2S含量升高是由于吸收空气中的H2S导致,并不是H2S合成酶基因被诱导表达。受胁迫植物释放的H2S还调节邻近植株的H2S响应基因表达(图2~3)和气孔运动(图4),即H2S不仅可以作为细胞内的信号分子发挥生理调节作用,还可以作为植物不同个体间的信息传递分子,参与整个植物群体对逆境胁迫的响应,为植物“语言”的组成添加了新的“成员”。在本研究的实验体系中,所使用的玻璃缸容积为7L,检测到受胁迫植物所在的环境H2S浓度升高,并且对邻近植株产生影响,但是在自然条件下,受胁迫植株释放到环境中的H2S的量,及H2S的有效作用范围和作用持续时间难以检测,这也是其它种类的绿叶挥发物需要进一步研究的内容。大部分创伤诱导挥发物或绿叶挥发物需要通过气孔进入邻近植物,而H2S气体信号分子不只通过气孔进入植物细胞。此外,茉莉酸甲脂[33],水杨酸[34]等需要经特定的受体激活信号途径,但H2S无特定的受体分子,目前研究发现H2S可以对多种蛋白质进行翻译后修饰(S-sulfhydration),对蛋白质的活性进行调节[35~36]。
动物个体间通过声音、行为、气味等方式传递信息,动物个体可以通过不同的音节组成不同的信息。那植物是否会通过释放不同的挥发物组合来达到类似于动物个体信息传递中不同音节的效果,从而实现对不同胁迫的预警机制?或者通过不同挥发物组合达到迅速或缓慢传递信息的目的?总之,这种植株间的信号交流是植物在长期的进化过程中形成的一种保护机制,对植物个体或群体的生存和繁衍都具有重要的意义。
4 结论
从本文的研究结果看来,H2S作为一个气体信号分子,它不仅可以增强植物自身对于逆境胁迫的抵抗力,还可以作为植株间信息交流的信号分子,向邻近的健康植株传递胁迫预警信号,使它们提前调整自身的新陈代谢,以应对可能即将来临的危险,这种植物进化出来的保护机制对植物种群的生存起到至关重要的作用。
