羌塘雪兔子繁殖特征及资源分配的海拔差异
2019-07-23马文梅王一峰赵夏纬
马文梅 王一峰 赵夏纬 李 蕾
(西北师范大学生命科学学院,兰州 730070)
植物从外界环境中获取资源,并且进行资源配置是植物生长的基本特性[1]。在植物个体的生长发育及繁殖等各种生命活动过程中,高海拔区植物必须根据不同器官的不同功能把有限的资源量进行最合理最优化的分配,为了在整个生命过程中维持各种器官的生理功能,植物在不同的物候期有不同的资源分配策略[2]。对植物器官的不同资源分配方案组成了植物生活史,即植物生活史特征可以通过不同的资源分配模式体现出来[3]。在胁迫环境下,植物可使用的资源总量是有限的,加大对某一活动(特征)资源量的投资必定会削减在其他活动资源量的投资,也就是说,在植物的不同活动(生活史特征)之间存在权衡关系。植物各器官的资源分配问题,主要可以从两个方面进行探究:一是繁殖分配,二是性分配。第一种分配策略是对植物的资源分配模式做出调整,用最优的分配对策适应环境的变化,以此来提升自身适合度。研究指出影响植物器官资源分配比例的因素有:植物的生活史特征、个体大小、种群密度以及交配系统等[4]。
有许多环境因素会影响植物生长,如水分、光照、温度及海拔高度等。海拔是影响植物生长的综合因子,海拔的变化往往伴随着温度、水分等一系列环境因素的变化,迫使高山植物调整其繁殖对策来应对较高海拔环境中的繁殖压力。随海拔高度的不同,植物的资源分配模式做出怎样的调整,是当前植物生活史理论研究的热点问题之一[5]。对青藏高原菊科植物沙生风毛菊(Saussureaarenaria)[6]、毛茛科植物露蕊乌头(Aconitumgymnandrum)[7]以及瑞香科植物狼毒(Stellerachamaejasme)[8]的研究均发现,随着海拔高度上升植物的繁殖分配增大,这一现象说明在严峻的环境中植物的繁殖比营养生长较为关键[9~10]。近年来,国内外对蓼科(Polygonaceae)、龙胆科(Gentianaceae)、虎耳草科(Saxifragaceae)等植物的资源分配问题已经有了较多研究[11~13],而广泛分布于青藏高原风毛菊属植物的相关研究还不够充分。当前分析不同海拔风毛菊属植物资源分配的差异一般是用居群内各器官生物量的平均值估量的,然而植物的生物量分配往往是异速的,并且受到表型可塑性和植株个体大小的影响,关于风毛菊属植物随海拔高度变化生物量分配的异速生长特点等问题的研究还鲜见报道。
青藏高原多数地区气候寒冷,生态因子(生物和非生物)相对复杂。因此随着海拔的上升植物可获得的资源总量相对较低,从而导致分配给繁殖器官的资源量也就会相应的减小[14]。风毛菊属是菊科植物中的一个大属,是青藏高原植被的重要组成部分。通过野外调查发现,风毛菊属植物中的羌塘雪兔子在青藏高原的分布范围差异较大(4 200~5 200 m均有分布),是研究繁殖分配的较为理想的材料。本文主要探究以下2个问题:(1)同一种群不同个体大小间的资源量将如何分配;(2)不同海拔下其花部特征如何变化?通过以上问题的研究,初步探讨羌塘雪兔子泛化结构的维持机制及繁殖对策,以期为全面研究羌塘雪兔子的生活史策略提供一定的理论依据。
1 材料与方法
1.1 研究材料
本实验所用材料为青藏高原常见植物羌塘雪兔子(材料来源见表1),本材料属多年生草本,无茎,莲座状;根褐色,呈圆锥状,肉质,根状茎被褐色残存的叶;叶线状披针形,叶片莲座状,无叶柄;小花呈紫红色,花果期在8、9月份。生活环境常为山坡草甸、沙地以及流石滩等,集中分布于西藏、青海、新疆等西北地区。
1.2 研究区概况
实验材料采自青海省兴海县,以及玛多县、雀儿山、巴颜喀拉山、风火山、唐古拉山口。样地位置在92°54′~99°55′E、31°04′~35°32′N,样地内海拔3 500~5 400 m,平均气温-4℃,年降雨量在200~500 mm,每年12、1、2月份气温最低,7、8月份气温最高,无霜期短。样地内风毛菊属植物资源非常丰富,且在山坡草甸及流石滩等生境具有较强的生长优势。
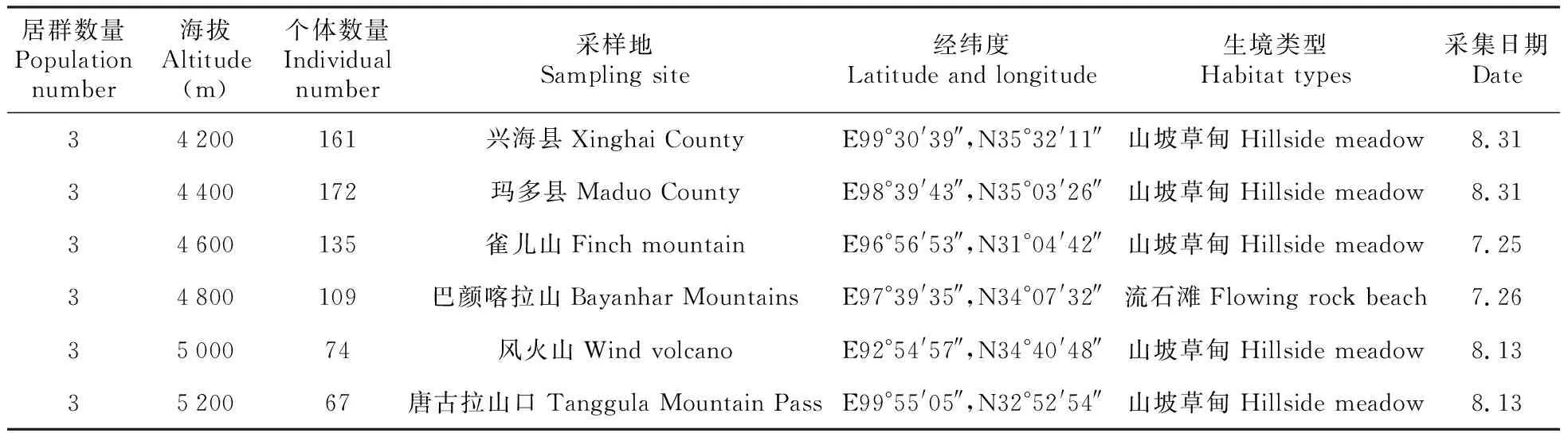
表1 羌塘雪兔子采样地概况
1.3 研究方法
2017年8月份分别在兴海县、玛多县、雀儿山、巴颜喀拉山、风火山、唐古拉山口选择6个海拔梯度(4 200、4 400、4 600、4 800、5 000、5200 m,受该区域地貌特征的限制,无法在空间相对狭小且在比较大的海拔梯度上进行取样,因而选择的样地所跨区域较大),每个海拔选取3个不同居群(居群间相隔200 m以上),在每个居群中采用随机取样的方式,采集15~25株完整的植株标本,并进行分株标号,采集结束回到实验室后,先将植株上的泥土用清水冲洗干净,对每株植物上的管状小花数量进行统计,用游标卡尺测量花柱、花丝、花药长度(以上生物量在每个居群随机抽取40~50朵完整管状小花测量后取平均值);在每个植株上选取花药未裂的花15~20朵,采用血球计数板法,取5次的平均值估算每朵花的总花粉数。然后将样本放置于80℃的烘箱中烘干24 h,之后使用1/10 000的电子天平秤测量各株植物在地上部分生物量、茎叶干重、头状花序重、每朵管状小花重量、雄蕊重量、雌蕊重量。
1.4 统计分析
(1)繁殖特征及资源分配与海拔的关系:植株个体大小用地上部分生物量干重表示;繁殖分配是指繁殖器官生物量(头状花序干重)与地上部分生物量干重的百分比;营养分配是指营养器官生物量(茎、叶干重)与地上部分生物量干重的百分比。我们用方差分析法来分析个体大小、繁殖器官生物量、营养器官生物量、营养分配、繁殖分配、雌蕊重量、雄蕊重量、花药长度、花丝长度、花柱长度及花粉数目与海拔的相关性;同时分析管状小花数目和一朵管状小花重量之间是否存在权衡关系。
(2)资源投入及资源分配与个体大小的关系:根据植株个体大小与资源分配的异速关系,本文采用异速方程:Y=axb,探究植株个体大小(x)与资源分配(Y)的内在关系,将变量lg转换后得到以下方程:lgY=lga+blgx(其中b代表斜率,lga为截距)[15]。在数据分析时对所用数据进行了对数转换(包括营养器官生物量、繁殖器官生物量、营养分配以及繁殖分配)。所有数据均采用Excel、SPSS 17.0进行处理和分析,采用origin 8.5作图。
2 结果与分析
2.1 不同海拔间羌塘雪兔子个体大小的变化
通过图1可以得知,羌塘雪兔子的个体大小与海拔高度呈负相关关系,即羌塘雪兔子的个体大小随海拔高度的增加而减小。羌塘雪兔子在海拔4 200 m处个体大小最大,是海拔5 200 m处的1.2倍。
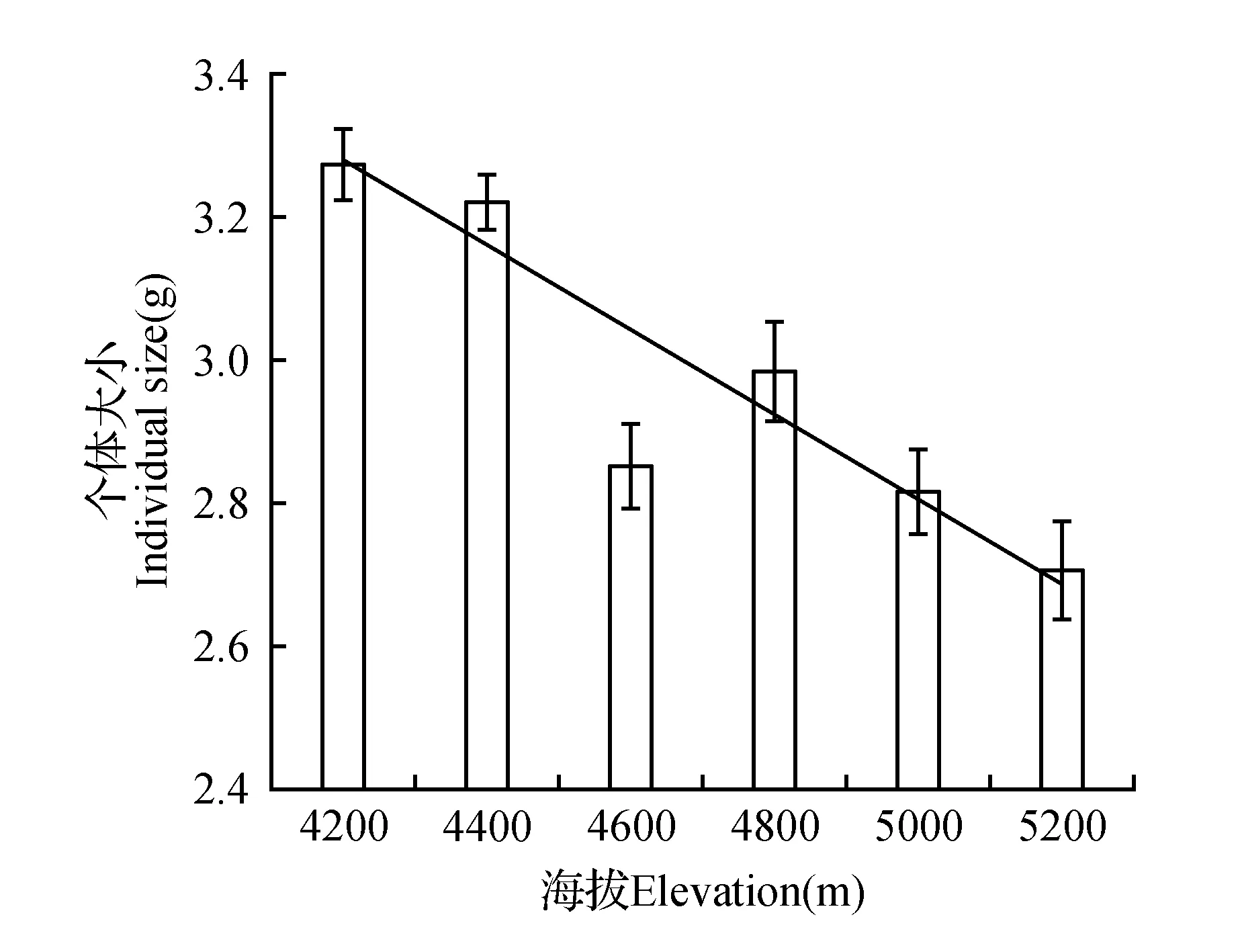
图1 不同海拔间个体大小的比较Fig.1 Comparisons of individual size among different elevation
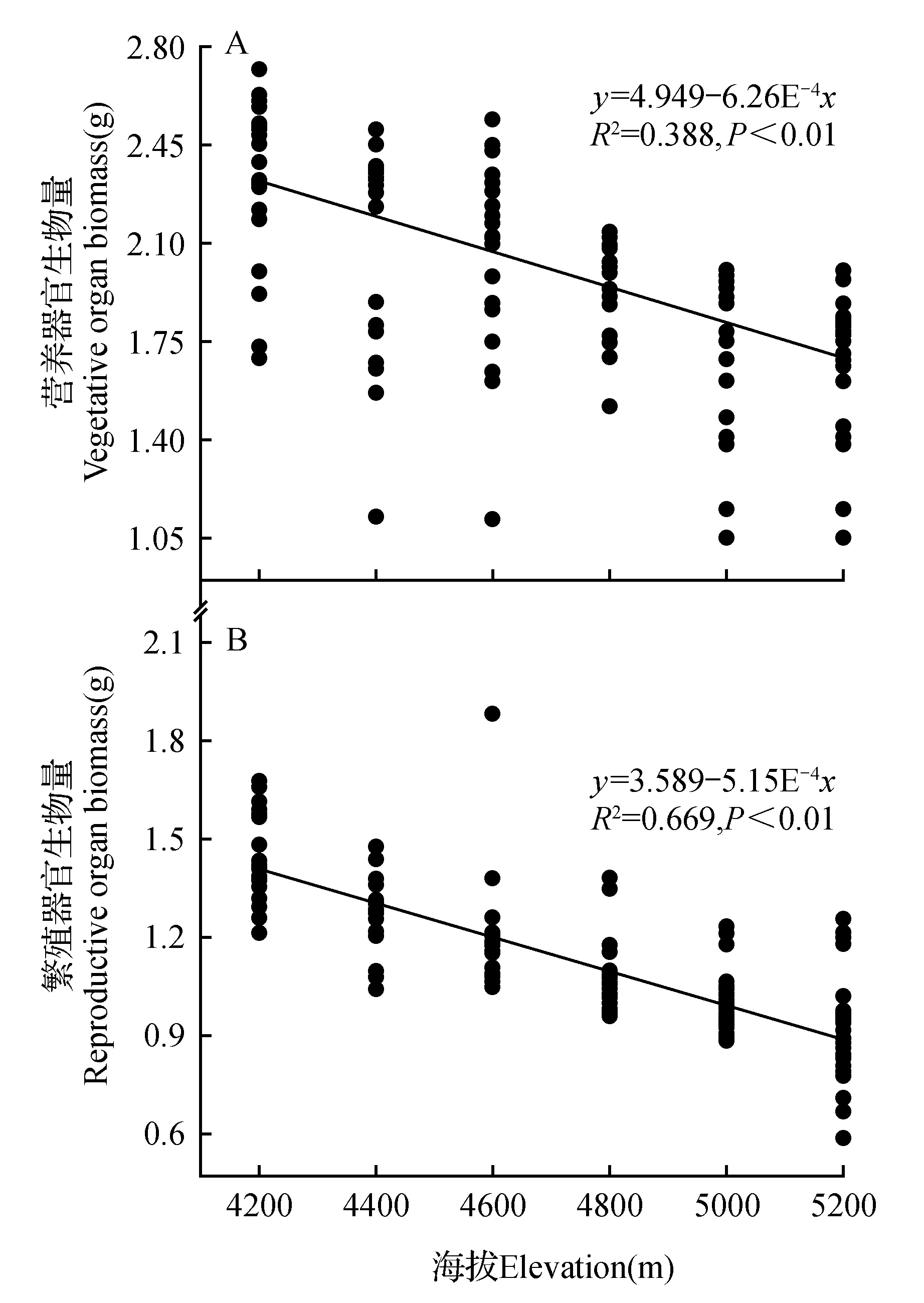
图2 营养器官生物量及繁殖器官生物量与海拔的线性回归Fig.2 Linear regression relationship between vegetative organ biomass,reproductive organ biomass and elevation
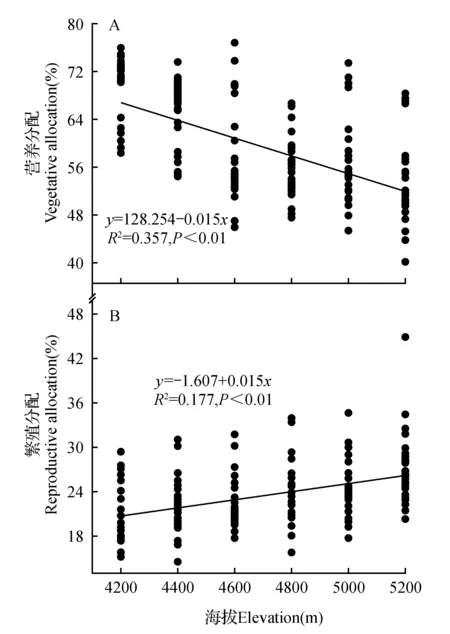
图3 营养分配及繁殖分配与海拔的线性回归Fig.3 Linear regression relationship between vegetative allocation,reproductive allocation and elevation
2.2 羌塘雪兔子资源投入及资源分配与海拔的关系
通过图2可以得知,羌塘雪兔子的营养器官生物量与海拔高度呈负相关关系(P<0.01),即羌塘雪兔子的营养器官生物量随海拔高度的增加而降低。海拔高度每增加200 m,营养器官生物量大约降低0.105 g。除此之外,羌塘雪兔子的繁殖器官生物量与海拔高度也呈现出负相关关系(P<0.01),即羌塘雪兔子的繁殖器官生物量随海拔高度的增加而降低。海拔高度每增加200 m,繁殖器官生物量大约降低0.093 g。
通过图3可以得知,羌塘雪兔子的营养分配与海拔高度呈负相关关系(P<0.01),即羌塘雪兔子的营养分配随海拔高度的增加而降低。海拔高度每增加200 m,营养分配大约降低2.572%。而羌塘雪兔子的繁殖分配与海拔高度呈正相关关系(P<0.01),即羌塘雪兔子的繁殖分配随海拔高度的增加而增加。海拔高度每增加200 m,繁殖分配大约增加1.009%。
2.3 不同海拔羌塘雪兔子资源投入及资源分配与个体大小的关系
从表2中可得知,羌塘雪兔子的营养器官生物量、繁殖器官生物量、营养分配、繁殖分配在不同海拔间与个体大小都呈显著正相关(P<0.05),即在不同海拔区域内,资源投入及资源分配随个体大小的增大而增大。
通过协方差分析可得,羌塘雪兔子营养器官生物量、繁殖器官生物量、营养分配、繁殖分配大小依赖的斜率在海拔间变化不显著(P>0.05),但截距在海拔间差异极显著(P<0.01),表明不同海拔生境对羌塘雪兔子营养器官生物量、繁殖器官生物量、营养分配、繁殖分配的大小依赖性产生了直接的影响。
2.4 羌塘雪兔子繁殖特征与海拔的关系
通过图4可以得知,羌塘雪兔子的管状小花数目与海拔高度呈负相关关系(P<0.01),即羌塘雪兔子的管状小花数目随海拔高度的增加而降低。海拔高度每增加200 m,管状小花数目减少约1个。而管状小花重量与海拔高度呈正相关关系(P=0.004),即羌塘雪兔子的管状小花重量随海拔高度的增加而增加。海拔高度每增加200 m,管状小花重量大约增加0.000 1 g。
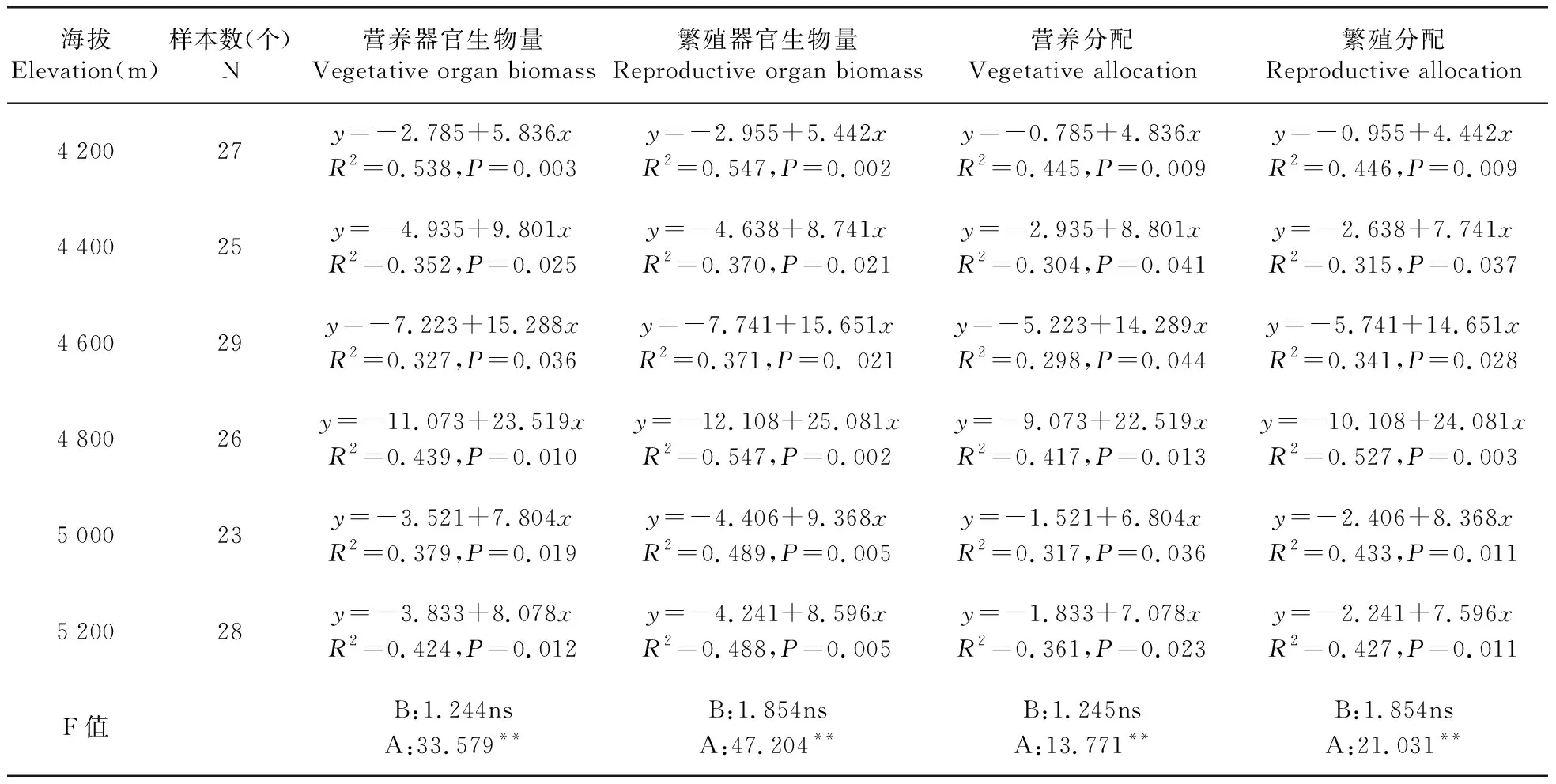
表2 不同海拔种群内个体大小与花期资源投入及资源分配的回归
注:A.截距;B.斜率;**.差异极显著(P<0.05);ns.差异不显著
Note: A.Intercept; B.Slope;**.Difference was significant atP<0.01; ns.Means no significant difference

图4 管状小花数目及重量与海拔的线性回归Fig.4 Linear regression relationship between tubular loret number,tubular loret mass and elevation
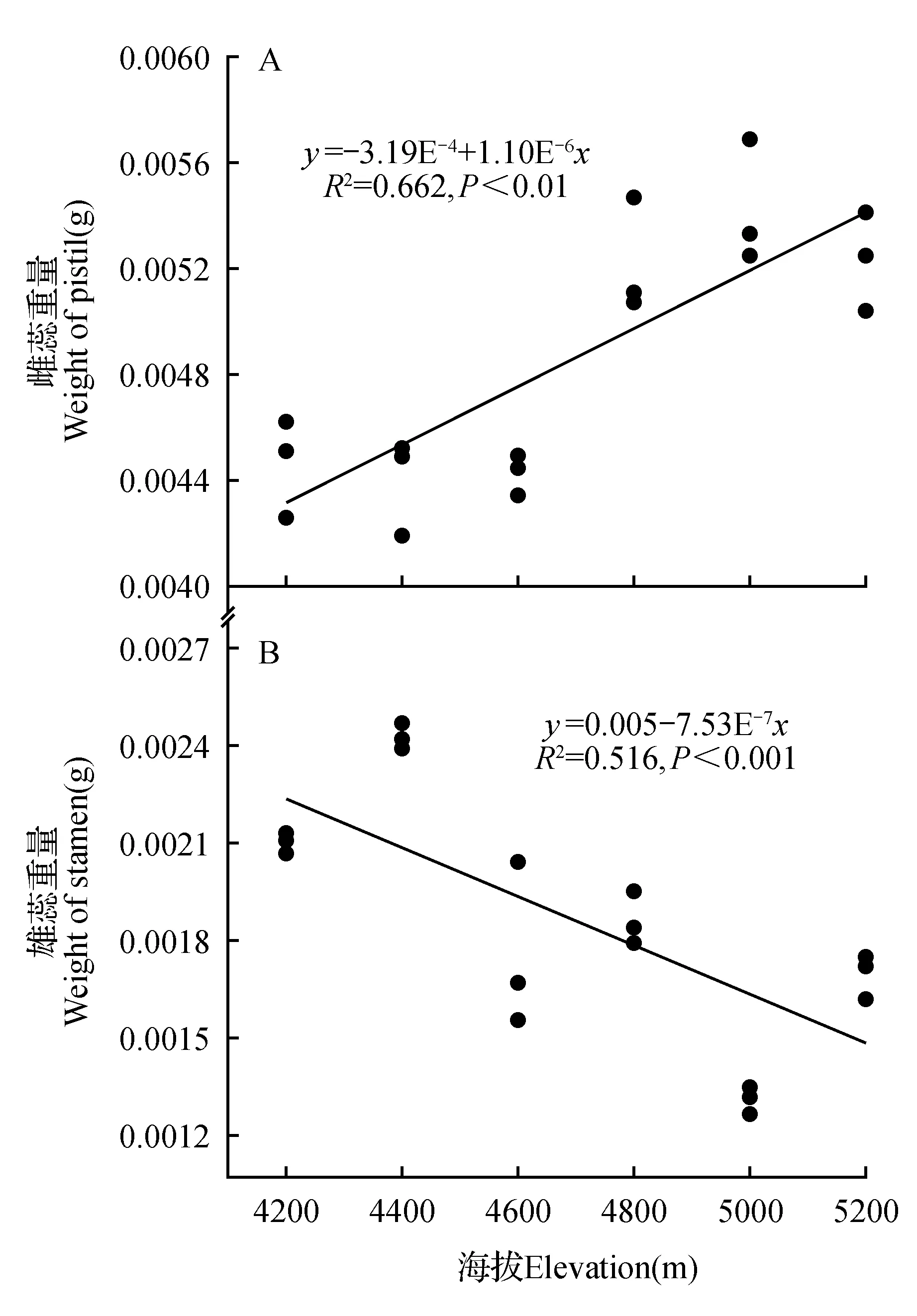
图5 雌蕊重量及雄蕊重量与海拔的线性回归Fig.5 Linear regression relationship between pistil weight,Stamen weight and elevation
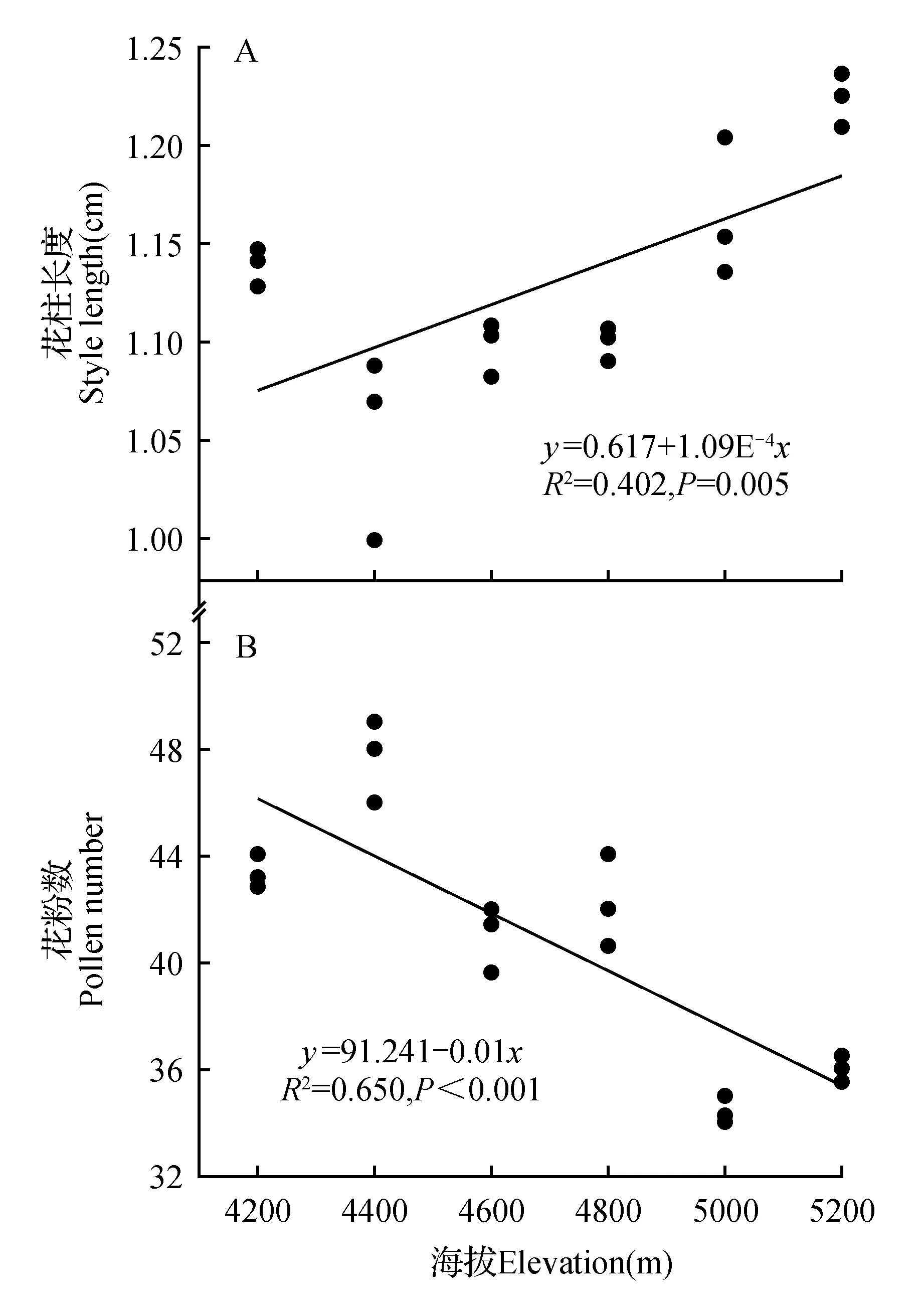
图7 花柱长度及花粉数与海拔的线性回归Fig.7 Linear regression relationship between style length,pollen number and elevation
由图5可知,羌塘雪兔子的雌蕊重量与海拔高度呈正相关关系(P<0.01),即羌塘雪兔子的雌蕊重量随海拔高度的增加而增加。海拔高度每增加200 m,雌蕊重量大约增加0.000 1 g。而雄蕊重量与海拔高度呈负相关关系(P<0.01),即羌塘雪兔子的雄蕊重量随海拔的升高而降低。海拔高度每增加200 m,雄蕊重量大约降低0.000 07 g。
由图6可知,羌塘雪兔子的花药长度与海拔呈负相关(P=0.027),即羌塘雪兔子的花药长度随海拔的升高而减小。海拔高度每增加200 m,花药长度大约减小0.000 9 cm。而花丝长度与海拔呈正相关(P<0.01),即羌塘雪兔子的花丝长度随海拔的升高而增大。海拔高度每增加200 m,花丝长度大约增加0.002 6 cm。
通过图7可以得知,羌塘雪兔子的花柱长度与海拔高度呈正相关关系(P<0.01),即羌塘雪兔子的花柱长度随海拔的增加而增大。海拔高度每增加200 m,花柱长度大约增加0.017 cm。而羌塘雪兔子的花粉数与海拔呈负相关(P<0.01),即羌塘雪兔子的花粉数随海拔的升高而减少。海拔高度每增加200 m,花粉数大约减少1.222×104。
通过图6~7可发现,随着海拔的升高,羌塘雪兔子的花药长度和花粉数均降低,而花柱长度增长,花柱长度的显著变化能够提升柱头对传粉昆虫的感受性,从而确保昆虫的授粉成功率,避免因花粉数的减少而影响其繁殖力。
2.5 生物量间的权衡关系
从表3可知,羌塘雪兔子管状小花重量与数目(P<0.05)、雌蕊重量与雄蕊重量(P<0.01)、花丝长度与花药长度(P<0.01)、花丝长度与花粉数(P<0.01)之间均呈负相关关系,说明这些繁殖特征之间均存在权衡关系。
3 讨论
3.1 海拔对羌塘雪兔子繁殖特征的影响
植物生长受环境的影响较大,因此需要对不同的生长环境做出适应性调整。通过研究可以看出,随海拔高度的增加,羌塘雪兔子个体大小、营养器官及繁殖器官生物量都明显降低。高山地区具有独特的生态系统,恶劣的生长环境和多变的气候。随着海拔高度的增加,空气密度、气压、温度和饱和蒸汽压等均有所降低,紫外线辐射增加,日变化大,这些不稳定的生态因子(非生物因子)对植物的胁迫增强,使羌塘雪兔子的生长受到限制。再者,植物能够从从外界获取到的资源总量降低,可以获取最大光照和能量的能力也降低,因此在可获得有限资源总量这一条件下,植物能够分配到各个器官的资源量也会减小。王一峰等人对8个不同海拔居群川西风毛菊(S.dzeurensis)的资源分配情况进行了研究,发现川西风毛菊的个体大小、营养器官及繁殖器官生物量随海拔升高均呈现出逐渐降低的趋势[16]。本研究结果与其一致。
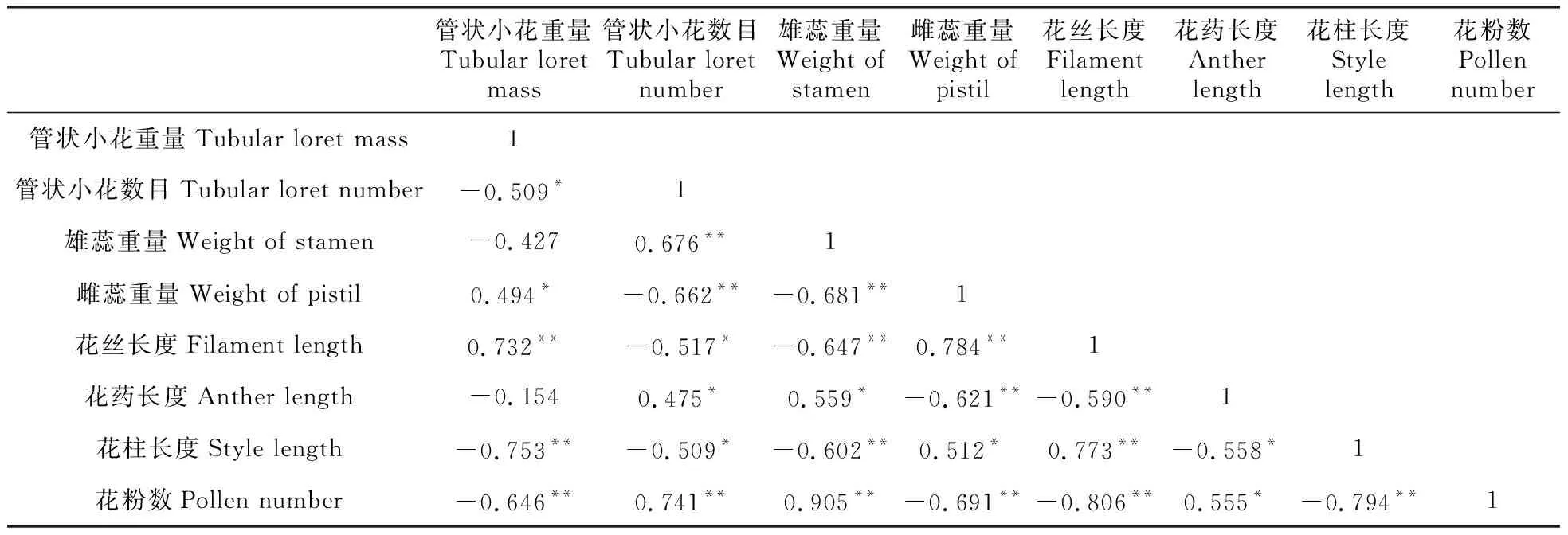
表3 繁殖特征的Pearson相关分析
*P<0.05;**P<0.01
繁殖分配指的是植物在整个生长发育过程中将同化产物分配到繁殖部分(头状花序)所占的比例,分配的越多则繁殖分配越高,反之则越低。繁殖分配的大小不仅受到自身遗传因素的影响,更重要的是对环境改变的响应[17],研究发现,随着海拔的上升,羌塘雪兔子面临更加恶劣的生存环境,植物能够通过光合作用获取到的资源不断减少,但其繁殖分配随海拔高度的增加而增大。由于植物资源总量有限,相应的羌塘雪兔子营养分配随海拔高度的增加而减小。在严峻的高山环境条件下,植物体为了使生命得以延续,需要将有限的资源和能量在生殖与生长之间进行最优化的分配,这是植物体对环境的一种适应性趋势。因此,羌塘雪兔子通过提高繁殖分配以更好地完成有性繁殖。
研究还发现,随着海拔高度增加,羌塘雪兔子花柱长度、花丝长度增加,而花粉数目显著减少。一方面,是由于在高海拔地区羌塘雪兔子将更多的资源用于增长柱头的支撑结构——花柱,有利于增加传粉昆虫在访花过程中花粉的携带量和柱头的接触面积,进而弥补了高海拔地区传粉昆虫数量少且活动频率低等对传粉过程中带来的不利影响。这种性器官的变异机制既合理的分配了高海拔区有限的可利用资源,又可以达到有效的传粉效果和有性繁殖的目的。另一方面,在外界资源受限的情况下,羌塘雪兔子通过减少花粉数,将有限的资源用于提高花粉质量以最大程度地保证花粉的生殖活力,从而确保其生活史的顺利完成。
3.2 不同海拔羌塘雪兔子个体大小与资源分配的关系
羌塘雪兔子的营养器官生物量、繁殖器官生物量、营养分配、繁殖分配在不同海拔间与个体大小都呈显著正相关,且截距在海拔间差异显著,说明海拔生境对羌塘雪兔子资源投入及资源分配的大小依赖性具有直接的影响。这是因为在自然生境中,植物个体大小与资源投入及资源分配之间的关系是由于某些环境因素造成的,而不是自然选择导致的结果,不具有遗传基础。
研究区在海拔高度为4 200~5 200 m范围内的青藏高原,羌塘雪兔子大多生长于山坡草甸、高山流石滩等不同生态环境中,致使海拔高度增加后温度、湿度及土壤肥力等的生境差异。这些微生境、微气候的变化不仅造成不同样地繁殖特征的变化,更造成不同海拔不同个体大小的资源分配的显著差异,这可能是造成羌塘雪兔子分布范围差异较大的主要因素。因此,在高寒区研究植物的资源分配问题时,我们更应该重视植物个体大小对资源分配的影响。
3.3 繁殖性状间的权衡关系
虫媒植物花的大小会对传粉者的吸引能力与植物的繁殖成功率产生一定影响,如果同一个体同时产生较多的花,将会导致同株异花授粉以及花粉不必要的耗费,而较大的个体花朵会增加自我维持的资源损失(如呼吸作用)[18]。性分配理论中有一个重要的假设:认为花的大小与数目之间存在着权衡关系[19~21],到目前为止,这种权衡关系仅在少数的研究中得到了证实。本研究结果指出,羌塘雪兔子的管状小花重量与数量之间显示出负相关关系。随着海拔的上升,羌塘雪兔子采取生产“少而优”的管状小花的对策。羌塘雪兔子做出这一选择,即存在这种权衡关系的原因可能是,如果把有限的资源投入于增加管状小花数目,那么不但要生成繁殖器官,还要生成总苞片等其他辅助构件,因此羌塘雪兔子把更多的有效资源投入到每个管状小花的大小上,这对于完善花部特征的构建及种群繁衍具有一定意义,且能够最大限度的促使繁殖成功。此外,研究还发现,羌塘雪兔子雌蕊重量与雄蕊重量、花丝长度与花药长度、花丝长度与花粉数之间也存在着这种权衡关系。对川西风毛菊(S.dzeurensis)、波缘风毛菊(S.undulata)、禾叶风毛菊(S.graminea)等不同植物的研究发现,植物会在许多生物量之间进行权衡[15,22~23]。植物在特定的时间内所获取的资源总量有限,在其整个生命历程中,包括生长、生殖、储存能量、修复等过程都需要投入资源和能量,所以植物体为了生长发育必须对有限的资源进行权衡分配[24]。
4 结论
本文以广泛分布于青藏高原高山草甸的羌塘雪兔子为研究材料,通过样地调查和实验分析获取数据,从花部特征、资源投入及资源分配等角度初步分析和探讨其繁殖特征随海拔的生态适应性对策。青藏高原气候环境复杂,许多以海拔为主的环境因子迫使高山植物调整其繁殖对策来应对较高海拔环境中的繁殖压力。羌塘雪兔子选择减小个体大小来应对青藏高原的高寒胁迫;花期繁殖分配随海拔增大,将有限的可利用资源投入到繁殖结构上,从而提高繁殖成功率;不同海拔生境对羌塘雪兔子资源投入及资源分配的大小依赖性产生了直接的影响;随着海拔的升高,羌塘雪兔子采取将雄蕊的花丝增长,雌蕊的花柱增长等花部特征变异的策略来提高授粉成功机率;管状小花数目随海拔升高而减小,而管状小花重量随海拔升高而增加,根据Pearson相关分析发现,这两者之间存在“此消彼长”的权衡关系,在其他繁殖特征间也发现了这种权衡关系。总的来说,羌塘雪兔子为了更好的适应青藏高原高寒环境,其繁殖特征随着复杂的气候变化表现出了很强的适应性。
