短期增温对亚高山草甸生物量和土壤呼吸速率的影响
2019-03-16欧阳青代微然单贵莲袁福锦吴文荣
欧阳青,任 健,代微然,尹 俊,单贵莲,袁福锦,吴文荣
(1.云南省草地动物科学研究院,云南 昆明 650212; 2.云南农业大学 草业科学系,云南 昆明 650201;3.云南省草原监督管理站,云南 昆明 650224)
由于人类活动的影响,大气中CO2的浓度已由工业革命前的280 μmol/mol增加到20世纪90年代初期的350 μmol/mol,再加上CH4等温室气体大量排放,全球气候出现变暖[1]。据模型估计,全球气温将升高1.5~4.5℃[2],在高纬度和高海拔地区温度增高幅度会更大,青藏高原的变暖趋势尤其突出[3-4]。陆地及其生态系统对气温增加的响应已成为植物生态学的研究热点之一。气候变暖的环境下,植物生长季内的降水格局、生长季长度和土壤温度、湿度等均会发生改变。
研究表明,温度升高对植物的物候期、群落和物种组成等方面有着显著的影响[5-6],尤其是高海拔、高纬度地区生长的植物[7],增温提高了高纬度冻原地区植物的生物量[8],改变了群落结构,促进了灌木的生长[9]。增温提高了海北矮嵩草草甸群落中禾草的生物量,减少了杂草的生物量[10]。Guan等[11]的研究表明,增温对青藏高原高山草甸0~20 cm土壤有机碳没有显著影响,未来增温可能会抑制高寒草甸生态系统在干旱生长季的碳排放[12],而且高山草甸年际间土壤呼吸的变化主要受控于降水量,而生长季节的土壤呼吸与土壤温度正相关,与土壤湿度呈负相关[13]。相反,有研究报道,土壤呼吸在气候变暖(温度平均升高0.3~0.6 ℃)情况下平均增加20%[14];在温带森林土壤中,发现土壤升温的第1,2和3年土壤呼吸速率分别增加了40%,14%和20%[15],这可能是温度升高影响了微生物或根系的代谢活性[16-17]。有研究发现,随着温度的进一步升高或较高温度持续时间的延长,土壤呼吸作用对温度升高响应的敏感性将逐渐降低,产生温度适应现象(Acclimation),从而减缓土壤呼吸随温度升高而增加[18-19]。
亚高山草甸是青藏高原东缘的主要植被类型,由于长期处于低温环境,降水量少,限制了土壤呼吸速率,积累了大量的有机碳,在全球升温的背景下,高寒地区积累的碳可能会大量释放出来,从而影响气候变化。但现阶段有关增温对亚高山草甸的植物群落、土壤呼吸等的研究较少。为此,选取滇西北亚高山草甸进行短期模拟增温试验,研究短期增温作用下不同季节的土壤呼吸速率及土壤温度、土壤含水量的变化,探究温度升高对亚高山草甸土壤呼吸作用的影响,以期为管理滇西北亚高山草甸生态系统,应对未来全球气候变化提供理论依据。
1 材料和方法
1.1 研究区域概况
试验区域位于云南省香格里拉市小中甸镇和平村的知特小组冬季牧场,选取地势平坦,植被分布相对均一,面积为100 m×50 m的亚高山草甸作为试验样地,地理位置N 27°20′~27°43′,E 99°36′~99°59′,海拔为3 200~3 220 m,年平均气温5.8℃,年均降水量849.8 mm,无霜期120 d,土壤类型为亚高山草甸土壤,草地类型为禾草-杂类草,属高原寒温湿润性气候,主要种类有苔草(Carexsp.)、华扁穗草(Blysmussinocompressus)、藨草(Scirpustriqueter)、高原毛茛(Ranunculustanguticus)、椭圆叶花锚(Haleniaelliptica)等,生长旺季为6~9月。
1.2 样地设置
2012年3月在试验地四周建立围栏,试验地中随机设置增温和对照处理,每个处理重复4次。试验利用开顶式生长室(OTC)进行增温,开顶式生长室为六边形,由6块透明有机玻璃纤维板拼接而成,底部面积为2.25 m2,顶部开口面积为1.2 m×1.2 m,考虑到滇西北亚高山草甸植物矮小,高度设置为0.6 m,参照Molau等[20]的方法,对照(CK)为OTC底面积大小、无有机玻璃纤维板的正六边形区域。其增温原理,在有机玻璃纤维板的阻挡作用下,生长室内风速降低,空气流动减弱,使热量不易散失;另一方面,太阳辐射中的红外线对有机玻璃纤维的穿透能力较好,使得内部温度升高,该设计对植物生长环境的影响降到最小。每个处理在每个对照和增温处理的地表中放置温度采集器(Microlite,FOURTEC,Israel)采集数据,设置每30 min自动记录1次。于2013年夏季、冬季,即处理第二年,进行植物群落生物量及土壤呼吸速率的测定。
1.3 测定指标及方法
1.3.1 地上生物量和地下生物量 2013年7月在植被生长旺盛的季节,从增温处理和对照的4个重复中随机选3个样区,并在处理区内随机取25 cm×25 cm的样方,地上部分采用齐地刈割法,将样方内的植物齐地剪刈,带回实验室于60℃烘箱中烘48 h至恒重后称重;地下生物量采用钻土块法,用直径3 cm土钻取样,按照距离地表0~10、10~20、20~30、30~40 cm分层取样,每个重复由3个样点的土样混合而成,将土样带回实验室过筛清洗,将根系挑出后于烘箱中60℃烘干48 h至恒重后称质量。
1.3.2 土壤呼吸速率,土壤温度及含水量 2013年夏季(7月)、冬季(10月)选择天气晴朗的日子,采用便携式土壤通量测量系统LI-8100 (LI-COR,Lincoln,USA)测量土壤呼吸速率。在增温处理和对照的4个重复中随机选3个小区,分别埋入PVC土壤呼吸环(内径20 cm,高12 cm),每个环需埋入地下7 cm,以便在整个观测过程中起到固定位置。测量前一天在尽量不扰动PVC土壤呼吸环中土壤的情况下,小心地沿地面剪去土壤呼吸环中所有植株的地上部分,以消除测定过程中植物自养呼吸的影响,24 h平衡后开始测定土壤呼吸速率。测定时间为8∶00~20∶00,每2 h测定1次。在测定土壤呼吸速率的同时,利用仪器自带温度探针和水分含量探头同步测量记录土壤7 cm深处的温度及土壤含水量。土壤地表温度用温度采集器(Microlite,FOURTEC,Israel)收集,在对照和增温处理的土壤表面放置温度采集器,每30 min自动记录1次温度数据。温度采集器自试验开始时放置,试验中仅采集2013年7月至2014年1月(即试验开展的夏季至冬季)的数据进行分析。
1.4 数据处理
利用统计学软件(SPSS 19.0)进行单因素方差分析(ANOVA),采用Duncan检验进行显著性比较,结果用Mean±SE表示,Excel 2003制作相关图表。
2 结果与分析
2.1 模拟增温对土壤地表及地下温度的影响
试验期间增温处理土壤地表平均温度为8.25℃,对照为6.69℃,较对照高出1.56℃,因而开顶式生长室改变了亚高山草甸的温度环境。从温度的动态变化分析,2013年7~9月增温处理高于对照0.46~0.58℃,即夏季的增温作用较小。进入冬季以后,增温处理的月平均地表温度比对照高1.24~3.6℃,增温效果更为明显(图1),原因是亚高山草甸夏季湿度大,相比于冬季土壤较为干燥,温度上升较快,且说明了气候温度较低的时候,模拟增温作用对土壤温度增加影响更为明显。
从地下7 cm土壤温度日变化分析,夏季和冬季土壤温度日变化均呈现“单峰”。夏季12∶00~16∶00,增温后的土壤温度明显高于对照,14∶00时达到最大,为26.02℃,比对照高2.11℃,随后呈现下降趋势。而冬季增温处理的土壤温度10∶00~14∶00明显高于对照,12∶00时相差1.78℃,同时14∶00达到最大,为16.18℃,随后急剧下降(图2)。地下7 cm的土壤温度,不论是夏季还是冬季,增温处理和对照的差异并不明显,说明在试验中短期增温并没有显著提高地下7 cm的土壤温度。
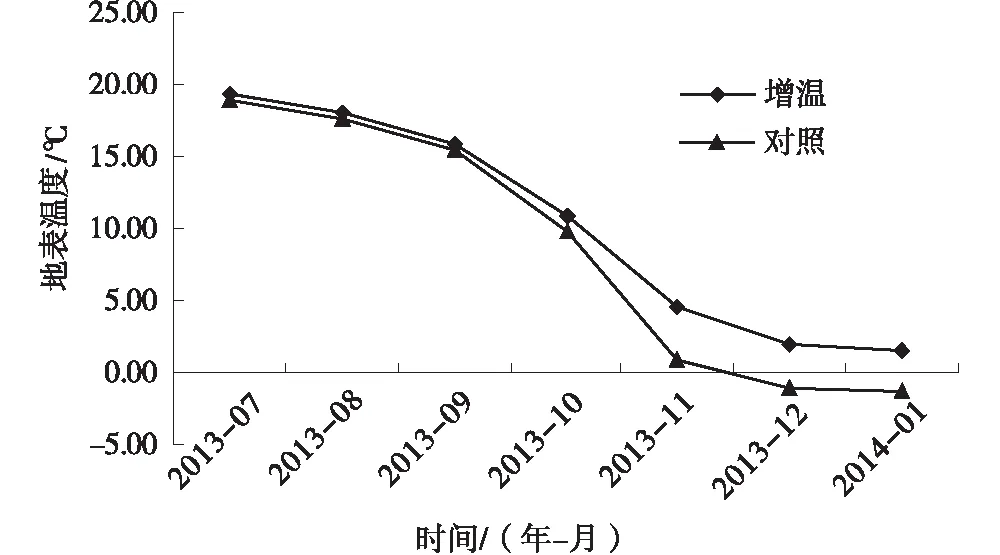
图1 增温对亚高山草甸地表月平均温度的影响(2013.7~2014.1)Fig.1 Monthly average soil surface temperature of subalpine meadows under warming treatment
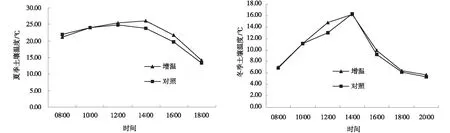
图2 增温作用下亚高山草甸夏季和冬季土壤的温度Fig.2 Warming effects on daily variation of soil temperature of subalpine meadows in summer and winter
2.2 模拟增温对土壤含水量的影响
从土壤含水量的日变化分析,夏季与冬季变化趋势不同,但是增温处理和对照区在夏季和冬季的变化曲线一致。夏季增温处理与对照之间差异不明显,在10∶00~12∶00,土壤含水量呈现最低,随后土壤含水量呈逐渐上升,而冬季到14∶00时,增温和对照的土壤含水量达到最低,与对照相比,增温使土壤水分降低了11.5%,随着时间推移,呈现先上升后降低的趋势,且对照高于增温处理(图3)。受日照影响,中午12∶00~14∶00为一天中温度最高的时间段,由于地表蒸发作用,土壤含水量也在该阶段达到最低,而夏季雨水较多,因而土壤水分相较于冬季变化较为舒缓,对比夏、冬季的土壤含水量变化,不难看出温度较低时,模拟增温作用对土壤含水量的影响更为明显,此趋势与土壤温度对模拟增温的响应一致。
2.3 模拟增温对地上和地下生物量影响
在增温作用下,夏季亚高山草甸植物群落的地上生物量比对照高148.76 g/m2,两者之间的差异显著(P<0.05)(图4)。0~40 cm地下生物量,增温作用使其在土层中的分布发生了改变,增温处理根系集中分布在0~10和10~20 cm土层,而对照则主要集中在0~10 cm。因而,0~20 cm的土层增温提高了地下生物量。增温作用促进了根系的发育,使得根系蔓延到10~20 cm土层。根系的垂直分布总体上都呈现从上到下的地下生物量分布减少的规律。这与土壤养分状况及高海拔地区的土壤特性有关,土壤表层储存有大部分的土壤养分和有机质,能够给予根系发育更多的养分。
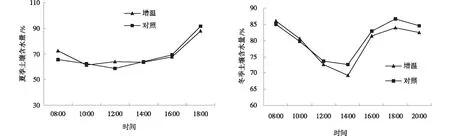
图3 增温作用下亚高山草甸夏季和冬季土壤的含水量Fig.3 Daily soil water content of subalpine meadows in summer and winter under warming treatment

图4 增温作用下亚高山草甸的地上和地下生物量Fig.4 Warming effect on aboveground and underground biomass of subalpine meadow注:不同小写字母表示处理与对照间差异显著(P<0.05)
2.4 模拟增温对土壤呼吸速率的影响
增温处理和对照夏季和冬季日动态(8∶00~20∶00)的土壤呼吸速率均呈现“单峰”曲线,先升高后降低,冬季的土壤呼吸速率明显低于夏季,且波动更大(图5)。从增温处理分析,土壤呼吸速率在夏季的最高值为5.66 μmol/(m2·s),而在冬季仅为2.79 μmol/(m2·s),相差较大。说明增温处理提高了土壤呼吸速率。
在夏季,从早晨8∶00开始,增温处理的土壤呼吸速率明显大于对照,随着时间的推移,二者的土壤呼吸速率均呈上升趋势,14∶00时土壤呼吸速率达到最高,且差距最小,随后,土壤呼吸速率呈现下降趋势,对照下降的更快,18∶00时差值达到1.19 μmol/(m2·s)。到冬季,增温和对照的土壤呼吸速率最高值也出现在14∶00,分别为2.79和3.25 μmol/(m2·s);14∶00后,对照组的土壤呼吸速率急剧下降。与对照0.53~3.52 μmol/(m2·s)的变幅相比,增温处理下的土壤呼吸速率日变化波动较小。
2.5 土壤呼吸速率日变化与温度、土壤水分的关系
相关性分析表明,增温处理和对照组的土壤呼吸速率和土壤温度呈正相关(表1),但相关性不显著;而土壤呼吸速率和土壤含水量呈现负相关,但相关性同样不显著。土壤温度和土壤含水量,在短期增温和对照组中,两者均呈极显著负相关(P<0.01)。
在冬季,相关性分析表明(表2),不管是增温处理还是对照区,土壤呼吸速率和土壤温度均呈极显著的正相关(P<0.01),土壤呼吸速率和土壤含水量呈现显著的负相关(P<0.05),土壤温度和土壤含水量呈极显著负相关(P<0.01)。
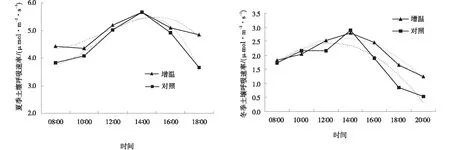
图5 增温作用下亚高山草甸夏季和冬季土壤的呼吸速率Fig.5 Warming effect on soil respiration rate of subalpine meadows in summer and winter
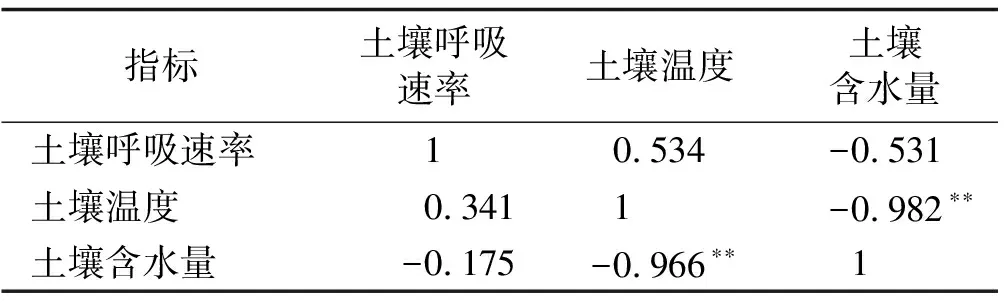
指标土壤呼吸速率土壤温度土壤含水量土壤呼吸速率10.534-0.531土壤温度0.3411-0.982∗∗土壤含水量-0.175-0.966∗∗1
注:**表示在0.01水平上显著相关,下同
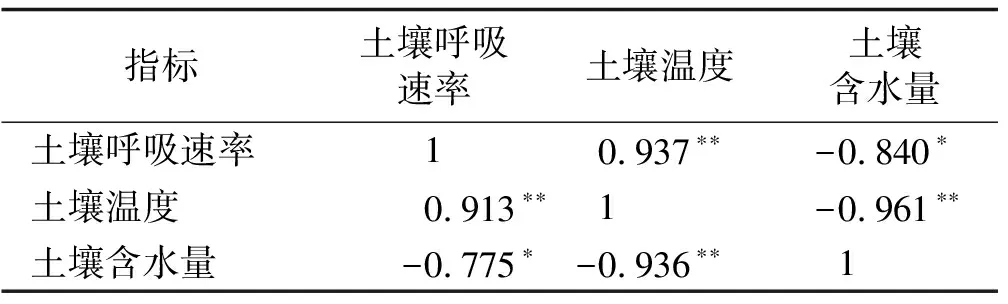
表2 土壤呼吸速率和土壤温度、土壤水分的相关性(冬季)
3 讨论
试验中使用的增温设施——开顶式生长室(OTC)与其他增温方法相比,不仅可以有效提高土壤的温度,而且具有方便易行、对生长环境的影响较小的特点,因而广泛地用于模拟增温的控制试验之中[21]。在OTC的作用下,试验中增温处理的平均地表温度为8.25℃,对照为6.69℃,处理较对照高出1.56℃,增温效果符合IPCC(2007)报告中预测的全球温度增加范围[2]。说明开顶式生长室能有效地提高土壤表面温度,其增温原理是在有机玻璃纤维板的阻挡作用下,生长室内风速降低,空气流动减弱,使热量不易散失;另一方面,太阳辐射中的红外线对有机玻璃纤维的穿透能力较好,使得内部温度升高[22]。
从地表温度的动态变化分析,OTC在冬季的增温效果高于夏季,原因是冬季增温处理与对照之间土壤湿度差别大有一定的关系,试验中测定7 cm土壤水分含量在冬季14∶00时,增温和对照的土壤含水量相差11.5%,而夏季相差不大。土壤温度的日变化呈现单峰曲线,尤其在冬季比较明显,表明气候温度较低的时候,模拟增温作用对冬季土壤温度的影响更明显,类似的结果在北美大平原草原(Northern Great Plains grassland)增温试验中上也有发现,Flanagan等[23]分析认为这与降雨的季节性有很大的关系。土壤含水量的日变化,夏季与冬季变化趋势不同,夏季变化较为舒缓,夏季增温处理与对照之间差别不明显,其原因是因为亚高山草甸夏季降雨较多,土壤水分处于饱和状态,使得增温对土壤水分的影响不大。而冬季不论是增温处理还是对照,土壤含水量均在14∶00时呈现一个低谷值,增温使土壤水分降低了11.5%。这与土壤温度对增温的响应一致,在气候温度较低时,模拟增温作用对土壤含水量的影响更大。冬季模拟增温引起的土壤含水量的降低与增温试验增强了土壤蒸腾作用有关[24],OTC使得土壤温度升高,进而增加了土壤水分的蒸发,这与Carey等[25]和Li等[26]的研究结果相一致。
研究报道,增温作用使土壤含水量明显降低[25-26],相反地上生物量显著增加[26]。这与试验结果一致,在试验中,模拟增温作用下,亚高山草甸在夏季的地上和地下生物量均高于对照,类似的增温效应在我国的海北矮嵩草草甸、北美大草原也有发现[10,23],且增温作用下的地上生物量显著高于对照组。尽管增温对草地植物有促进生长作用,但分析时不能一概而论,因为草地存在很大的异质性,降水等因素同样影响植物生长,例如,在北美大草原的试验中,第年降水变化导致增温的效应不明显。另外,在草地上根系的分解大概需要5~7年的时间[27],因而有必要开展更长的增温试验来确认地下生物量的变化。
研究发现,模拟增温下,土壤呼吸速率均高于对照,温度增加亚高山草甸将释放出更多碳,这进一步证明了Liu等[28]研究结果。从土壤呼吸速率日变化的分析,夏、冬季的趋势基本相同,均呈现出了单峰曲线,在14∶00出现峰值。且冬季的土壤呼吸速率的单峰曲线更加明显。这与土壤温度的日变化曲线有着相似之处,且土壤呼吸速率随着土壤温度的下降而下降。这说明土壤呼吸和土壤温度相互影响,且变化趋势相同。一昼夜内的温度变化相对较大,水分变化相对较小,所以土壤呼吸作用的日动态主要受温度控制[29]。不过,土壤的呼吸作用同时受降雨的影响。在典型草原土壤呼吸特征的研究中,王庚辰等[30]发现,丰水年土壤呼吸与土壤温度有较强的关系,而在干旱年份,土壤呼吸与土壤温度的关系明显减弱,可见土壤含水量在土壤呼吸与温度之间的相关性方面有着重要的制约作用。Wu等[31]认为增温将使得土壤呼吸速率平均提高12%、生态系统的呼吸作用提高27%。有两方面的原因导致增温促进土壤呼吸作用,其一是温度增加直接提高了呼吸速率;其二温度增加使得更多的碳能分配给根系、土壤中的菌类以及分泌物,从而间接地提高了呼吸作用[32]。
影响土壤呼吸的因子众多,不同研究条件下得出的土壤呼吸速率与土壤温度(不同深度)、土壤含水量、气温等的相关性不同,且影响土壤呼吸速率的相关因子之间并不是相互孤立,它们之间也是相互影响。除了温度、土壤水分含量外,土壤呼吸速率还与气候变化、植被状况、凋落物分解等多种因素密切相关,特别是在水分成为限制因子的干旱半干旱地区,水分与土壤温度共同影响土壤的呼吸作用[33]。从土壤呼吸速率、温度、水分的相关性分析,试验中,无论增温处理还是对照,冬季亚高山草甸的土壤呼吸速率、土壤温度和土壤含水量两两之间均呈现显著相关性,尤其是土壤呼吸速率与温度之间呈现出极显著的正相关性,土壤温度和土壤含水量两者之间则呈现出了极显著的负相关性,而夏季增温处理和对照的土壤温度和土壤含水量也呈现出了极显著的负相关性。这与Lin等[34]的研究发现一致。据Joo等[35]的研究结果,土壤含水量低于20%时,土壤呼吸速率与水分呈正相关,高于20%则呈现负相关。试验中土壤呼吸速率的最高值出现温度较高的12∶00~14∶00,表明了土壤呼吸受到水、热等因子的共同作用,出现日变化单峰曲线,最高值出现在水热单配较好的时期。
4 结论
短期增温显著提高了滇西北亚高山草甸的地上生物量,并促进了根系向10~20 cm土层扩展。短期增温对滇西北草地含水量的影响与季节有关,在气候寒冷时,增温对土壤温度和土壤含水量的影响更为明显。短期增温提高了土壤呼吸速率,且夏季的土壤呼吸速率显著高于冬季。相关性分析发现,土壤呼吸速率与土壤温度呈现出显著的正相关性,而土壤温度和土壤含水量两者之间则呈现出了显著的负相关性。
