鲜切竹笋伤害信号的初步研究
2018-06-28肖尚月金图南罗海波孙金才
何 雄,肖尚月,金图南,罗海波,孙金才
(1.浙江医药高等专科学校 食品学院,浙江 宁波 315100;2.南京师范大学 金陵女子学院,江苏 南京 210097)
鲜切竹笋伤害信号的初步研究
何 雄1,肖尚月1,金图南1,罗海波2,孙金才1
(1.浙江医药高等专科学校 食品学院,浙江 宁波 315100;2.南京师范大学 金陵女子学院,江苏 南京 210097)
摘 要:初步研究鲜切处理对距竹笋切割位点不同距离切段中伤害信号传递的影响,并通过外源信号分子茉莉酸甲酯(methyl jasmonate,MeJA)及其合成抑制剂水杨苷异羟肟酸(salicylhydroxamic acid,SHAM)处理,探索MeJA在鲜切竹笋信号分子合成和传递中的作用。结果表明:在切割位点产生了伤害信号物质,并能传导至邻近未受伤组织中,诱导苯丙氨酸解氨酶(phenylalanine ammonia lyase,PAL)、过氧化物酶(peroxidase,POD)活性的升高。MeJA处理能引起鲜切竹笋PAL、POD、超氧化物歧化酶(superoxide dismutase,T-SOD)和抗坏血酸过氧化物酶(ascorbate peroxidase,APX)活性升高,而SHAM处理对上述酶的活性升高具有一定的抑制作用,表明MeJA可能诱导并参与了鲜切竹笋在受到机械伤害后某种伤害信号的合成和传导。
关键词:植物生理;茉莉酸甲酯;竹笋;信号传导
竹笋属禾本科多年生草本植物,味鲜质脆,素有“蔬菜中第一品”之美称,作为健康天然的特色蔬菜,常以切片或切丝的方式食用,极其适合鲜切菜生产。但是,鲜切竹笋因经过去壳、切分、清洗和包装等处理,极易产生褐变和木质化等品质劣变现象,商品价值急剧下降[1-2]。因此,如何延缓鲜切竹笋品质劣变、通过合理的处理延长保鲜时间已成为亟待解决的重要问题。研究表明,切分造成的机械损伤对鲜切蔬菜生理生化代谢、风味品质,以及安全性都有较大影响,如乙烯产生量增加、呼吸速率提高、次生代谢产物和自由基大量产生积累、膜脂质过氧化作用加剧、酚类物质氧化反应加强,以及愈伤组织形成等。这一系列生理生化的异常变化都可能与伤害信号的产生和传导有关[3-7]。目前,国内关于鲜切竹笋的研究主要集中在品质劣变控制等方面[8-10],对鲜切竹笋品质劣变的机理及鲜切后伤害信号的产生与传导的研究甚少。
植物在长期的进化过程中形成了自身的生化防御系统,当受到外界机械伤害刺激时,会诱导一系列防御反应[11]。研究表明,茉莉酸甲酯(methyl jasmonate,MeJA)具有信号功能,当植物受到生物或非生物胁迫时能作为一种内源信号分子诱导相关抗逆性基因的表达,增强植物的抗逆性[12]。但是,国内对伤害信号的研究多以整棵植株对外界环境胁迫的伤害反应为主,对鲜切蔬菜伤害信号的研究报道较少。本实验初步研究了竹笋切分后伤害信号特征及切分位点邻近部位的生理生化变化规律,拟探究MeJA对鲜切竹笋信号分子合成和传递的影响,旨在为进一步探索鲜切竹笋伤害信号的弱化途径、建立鲜切竹笋贮运保鲜新方法提供依据。
1 材料与方法
1.1 材料与仪器
实验材料。供试竹笋于2017年12月购自浙江宁波当地大型超市。
试剂。茉莉酸甲酯(分析纯),购自上海瑞永生物技术有限公司。水杨苷异羟肟酸(salicylhydroxamic acid,SHAM),购自阿拉丁试剂有限公司。无水乙醇、过氧化氢、氯化钠、磷酸氢二钠、磷酸二氢钠,均为分析纯。苯丙氨酸解氨酶(phenylalanine ammonia lyase,PAL)测定试剂盒、过氧化物酶(peroxidase,POD)测定试剂盒、超氧化物歧化酶(superoxide dismutase,T-SOD)活性测定试剂盒、抗坏血酸过氧化物酶(ascorbate peroxidase,APX)、蛋白定量测试盒(考马斯亮蓝法),均购自南京建成生物工程研究所。
仪器。Thermo MR 23i型离心机,Thermo Fisher Scientific Inc.;AL104/01型电子分析天平,Mettler Toledo;SCG-4型恒温水浴锅,宁波新芝生物科技股份有限公司;GBC Cintra 20型紫外分光光度计,GBC Scientific Equipment。
1.2 方法
1.2.1 竹笋预处理
选择新鲜的、无明显机械损伤、大小均一的竹笋作为实验材料。运回实验室后,置于(4.0±0.5)℃的冰箱预冷6~12h,预冷后,去除笋壳、杂质和底端,清洗,沥干,备用。
1.2.2 伤害信号传导实验
将完整竹笋从顶部向底端5~6cm处横切,横截面切齐。采用低密度聚乙烯(LDPE)保鲜袋包装后于(4.0±0.5)℃贮藏。贮藏期间每3d取样1次,取距离切割位点0~7、7~14、14~21mm部位,分别测定PAL、POD活性。
1.2.3 外源信号分子MeJA处理
将鲜切笋随机分成2组,一组用1μmol·L-1MeJA浸泡处理5min,一组以蒸馏水浸泡5min,作为对照。沥干,贮藏条件同1.2.2节。贮藏期间每3d取样1次,取距离切割位点0~7mm部位测定酶活。
采用0.5% SHAM浸泡处理5min。沥干,贮藏条件和取样方法同上。
1.2.4 PAL、POD、T-SOD、APX酶活测定[13]
PAL活性测定。按组织质量(g)-提取液体积(mL)1∶9的比例,冰水浴匀浆,1000r·min-1离心10min,取40μL粗酶液测定D290值,并计算其活力,单位U·mg-1(以蛋白质质量计)。
POD活性测定。采用愈创木酚法。按组织质量(g)-0.1mol·L-1磷酸盐缓冲液(pH7~9)体积(mL)1∶9的比例,冰水浴条件下制备组织匀浆液,3500r·min-1离心10min,取100μL粗酶液测定D420值,并计算其活力,单位U·mg-1(以蛋白质质量计)。
T-SOD活性测定。按组织质量(g)-0.1mol·L-1磷酸盐缓冲液(pH7.0~7.4)体积(mL)1∶4的比例,冰水浴条件下制备组织匀浆液,3500r·min-1离心10min,取40μL粗酶液测定D550值,并计算其活力,单位U·mg-1(以蛋白质质量计)。
APX活性测定。按组织质量(g)-缓冲液体积(mL)1∶9的比例,冰水浴匀浆,1000r·min-1离心10min,取100μL粗酶液测定D290值,并计算其活力,单位U·mg-1(以蛋白质质量计)。
蛋白质定量测定。采用考马斯亮蓝法。按组织质量(g)-生理盐水体积(mL)1∶9的比例,冰水浴条件下机械匀浆,2500r·min-1离心10min,取上清液,再用生理盐水同前比例稀释,取50μL样本测定D595值,并计算样本蛋白浓度,单位mg·mL-1。
1.3 数据分析
每个处理重复3次,样品测定时均设置2个平行。利用Excel 2010和Origin 8.5.1进行数据整理、方差分析与制图。
2 结果与分析
2.1 鲜切竹笋不同部位PAL和POD的活性变化
由图1可知,鲜切竹笋在4℃冷藏的前9d,不同切段PAL活性均快速升高,但距离切点越远的切段,PAL酶活性的增长速率越慢,其酶活性也越低。冷藏9d以后,各切段PAL活性均呈下降趋势。整体来看,冷藏过程中,0~7mm竹笋切段的PAL活性高于其他切段。鲜切竹笋在贮藏过程中,POD活性呈上升趋势,在贮藏前期(0~12d),各竹笋切段POD活性快速上升,0~7mm竹笋切段中的POD活性增长较快,活性整体高于其他切段。结果说明,鲜切能引起竹笋PAL和POD活性升高,推测由切割产生的信号分子会从切割位点向邻近组织细胞传递,引起植物的抗逆性反应。
2.2 MeJA处理对鲜切竹笋PAL、POD、T-SOD和APX酶活性的影响
由图2可知,鲜切竹笋在贮藏过程中,PAL和POD活性变化呈先上升后下降的趋势,MeJA处理组PAL活性在贮藏第6天达到高值,POD活性在贮藏第9天达到高值,且显著(P<0.05)高于对照组,说明MeJA处理能像机械伤害一样诱导植物组织中PAL和POD活性上升,由此推测,MeJA可能参与了竹笋防御相关的信号传导途径或伤害信号反应[14]。在贮藏后期,MeJA处理组的PAL和POD活性与对照组无显著差异。这可能是因为茉莉酸介导的信号途径主要在竹笋贮藏前期的伤害应答反应中起作用[15]。鲜切竹笋在贮藏过程中,T-SOD活性呈现先上升后下降的趋势。在贮藏前期,T-SOD活性快速增长,贮藏3~12d,MeJA处理组的T-SOD酶活性显著(P<0.05)高于对照组。这可能是由于鲜切竹笋在贮藏过程中产生了大量活性氧,T-SOD作为植物体抗氧化系统的第一道防线,为清除自由基、推迟膜脂过氧化进程,而被大量诱导表达[16]。APX酶活性在鲜切竹笋贮藏过程中呈先下降后上升再下降的变化趋势。在贮藏前9d,MeJA处理组的APX活性高于对照组,在贮藏第6天达到高值,显著(P<0.05)高于对照组,但在贮藏12d后,APX活性和T-SOD活性均降低。这可能是因为随着贮藏时间延长,伤害胁迫和活性氧大量积累,抑制了T-SOD和APX的活力[17]。
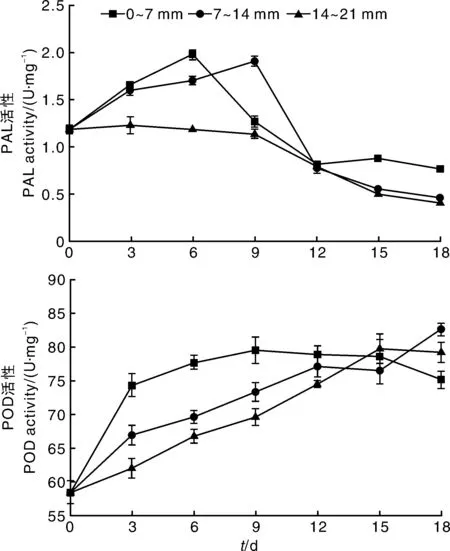
图1 竹笋鲜切后距切割位点不同距离切段中PAL和POD活性随时间的变化Fig.1 Changes of PAL and POD activities in different sections of fresh-cut bamboo shoots from cutting sites over time
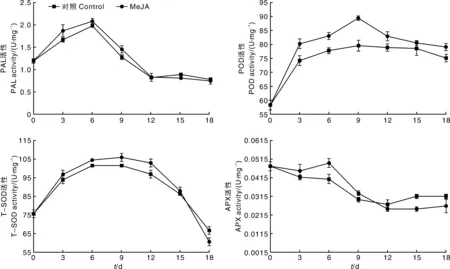
图2 MeJA处理对鲜切竹笋切段中PAL、POD、T-SOD和APX活性的影响Fig.2 Effects of MeJA on PAL,POD,APX,T-SOD activities in fresh-cut bamboo shoots
2.3 SHAM处理对鲜切竹笋PAL、POD、T-SOD和APX酶活性的影响
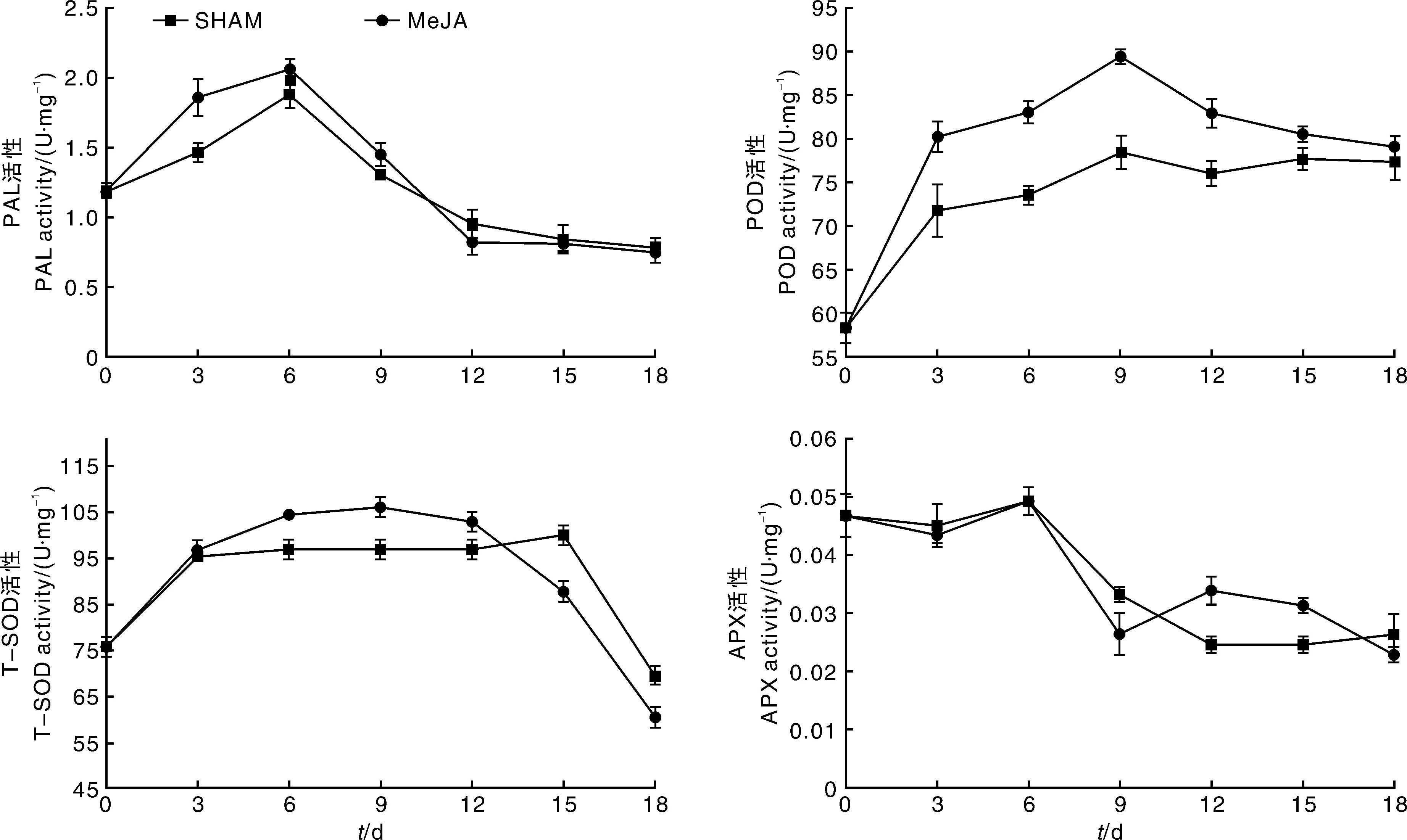
图3 SHAM处理对鲜切竹笋切段中PAL、POD、T-SOD和APX活性的影响Fig.3 Effects of SHAM on PAL,POD,APX,T-SOD activities in fresh-cut bamboo shoots
由图3可知,SHAM处理后鲜切竹笋PAL、POD和T-SOD活性呈先上升后下降的趋势。鲜切竹笋贮藏3~12d,相较于MeJA处理组,SHAM处理对PAL、POD和T-SOD酶活性起到了抑制作用,在贮藏第9天,SHAM处理组的POD和T-SOD活性显著(P<0.05)低于MeJA处理组。SHAM是茉莉酸的抑制剂,可能由于SHAM抑制了内源茉莉酸的合成[18],影响了伤害信号的传递,因而抑制了上述防御酶活性的升高。
3 讨论
为更好地适应自然环境,植物在长期的进化过程中形成了生化防御系统,能在受到机械伤害胁迫时在伤害部位产生内源物质作为伤害相关分子模式(damage-associated molecular patterns,DAMPs)[15]。竹笋鲜切后9d内,距离切割位点不同距离部位的PAL活性均快速增加,但增速存在区别,与马玉荣等[19]结果类似。这可能是因为前期伤害诱导了PAL前体物质的转化,导致PAL合成,随后伤害信号激活防御基因的表达,诱导PAL合成[20],表明机械伤害诱导了鲜切竹笋中伤害信号物质的产生。PAL和POD活性从切割位点向邻近部位逐渐降低,由此推测,竹笋鲜切后存在伤害信号物质,且能将伤害信号从切割位点传递到邻近的组织细胞中,从而引起整个组织对外界胁迫的抗性反应,这与前人的结果一致[21]。
茉莉酸与植物抗性密切相关。研究表明,内外源茉莉酸/MeJA均可以将伤害信息传递到植物体的其他组织部位,进而诱导一系列防御反应的发生[22]。MeJA作为细胞信号传导系统的重要物质,参与植物防御信号的转导与放大过程[23]。本研究表明,MeJA处理后能明显提高PAL、POD和T-SOD活性,与前人研究结果一致[24-25]。苯丙烷代谢作为植物遭受伤害胁迫而诱导的次生代谢途径,是鲜切竹笋的重要防御途径[26],POD是植物在逆境条件下酶促防御系统的关键酶之一,和SOD相互协调配合,清除过剩的自由基,以提高植物的抗逆性[24]。结合本实验结果,推测MeJA可能与鲜切竹笋防御相关信号的传导途径或伤害信号反应相关。APX酶在植物活性氧清除系统中居于核心地位,是胞内氧化还原态水平调控的关键抗氧化物酶组分[27]。MeJA处理的鲜切竹笋在贮藏前期APX活性高,而贮藏后期活性降低,与马杰等[28]研究结果一致。
SHAM是茉莉酸类物质的抑制剂[18]。当植物受到伤害时,植物体内茉莉酸及其衍生物的含量显著增加,能诱导一系列与抗逆有关的基因表达[22]。本研究表明,SHAM处理对PAL、POD和T-SOD活性起抑制作用,显著降低POD和T-SOD活性高值,可能是因为SHAM处理降低了鲜切竹笋内源茉莉酸的含量,进而抑制了相关防御酶活性的升高,进一步说明茉莉酸甲酯可能参与了鲜切竹笋受到机械伤害后某种伤害信号的合成与传导。
综上,本研究表明,在竹笋切割后,在切割点会产生伤害信号物质,并能传递到邻近未受伤组织中,发起植物组织内部的防御反应。外源信号分子MeJA能诱导与鲜切竹笋防御系统相关的PAL、POD、T-SOD、APX活性升高,说明MeJA可能与鲜切竹笋防御相关的信号传导途径和伤害信号反应相关。SHAM作为MeJA的合成抑制剂,能抑制鲜切竹笋PAL、POD、T-SOD活性增强,由此进一步说明MeJA可能诱导并参与了鲜切竹笋在受到机械伤害后某种伤害信号的合成与传导。但是对于该信号物质具体为何种物质、通过何种途径产生等,还有待进一步的研究和论证。
:
[1] LU S M,XU Y G.Physiological and biochemical changes of fresh-cut bamboo shoot(Phyllostachysheterocyclevar.pubescens) during cold storage[J].JournaloftheScienceofFood&Agriculture,2004,84(8):772-776.
[2] 周琦,陈季旺,高俊,等.鲜切雷竹笋冷藏过程中木质化机理的研究[J].食品科学,2012,33(14):307-311.
ZHOU Q,CHEN J W,GAO J,et al.Lignification mechanism of fresh-cut bamboo shoots during cold storage[J].FoodScience,2012,33(14):307-311.(in Chinese with English abstract)
A B,BARAT J M,et al.Extending and measuring the quality of fresh-cut fruit and vegetables:a review[J].TrendsinFoodScience&Technology,2007,18(7):373-386.
[4] FRANCIS G A,GALLONE A,NYCHAS G J,et al.Factors affecting quality and safety of fresh-cut produce[J].CriticalReviewsinFoodScience&Nutrition,2012,52(7):595-610.
[5] WATADA A E,KO N P,MINOTT D A.Factors affecting the quality of fresh-cut produce[J].PostharvestBiology&Technology,1996,9(2):115-125.
[6] TOIVONEN P M A,BRUMMELL D A.Biochemical bases of appearance and texture changes in fresh-cut fruit and vegetables[J].PostharvestBiology&Technology,2008,48(1):1-14.
[7] 王健,宋亚,方佳宁,等.植酸处理对鲜切紫甘蓝品质的影响[J].浙江农业学报,2015,27(11):2017-2023.
WANG J,SONG Y,FANG J N,et al.Effect of phytic acid on the quality of fresh-cut red cabbage[J].ActaAgriculturaeZhejiangensis,2015,27(11):2017-2023.(in Chinese with English abstract)
[8] 罗海波,何雄,包永华,等.鲜切果蔬品质劣变影响因素及其可能机理[J].食品科学,2012,33(15):324-330.
LUO H B,HE X,BAO Y H,et al.Affecting factors and possible mechanisms of quality deterioration in fresh-cut fruits and vegetables[J].FoodScience,2012,33(15):324-330.(in Chinese with English abstract)
[9] 陈海光,冯卫华,白卫东,等.鲜切竹笋褐变控制的研究[J].中国食品学报,2010,10(4):233-239.
CHEN H G,FENG W H,BAI W D,et al.Studies on control of browning in fresh-cut bamboo shoots[J].JournalofChineseInstituteofFoodScience&Technology,2010,10(4):233-239.(in Chinese with English abstract)
[10] 罗自生,张莉.壳聚糖/纳米SiOx复合物涂膜对鲜切竹笋品质和生理的影响[J].中国农业科学,2010,43(22):4694-4700.
LUO Z S,ZHANG L.Effect of chitosan/nano-SiOx complex on quality and physiology of fresh-cut bamboo shoot[J].ScientiaAgriculturaSinica,2010,43(22):4694-4700.(in Chinese with English abstract)
[11] RYAN C A.Protease inhibitors in plants:genes for improving defenses against insects and pathogens[J].AnnualReviewofPhytopathology,1990,28(1):425-449.
[12] 李清清,李大鹏,李德全.茉莉酸和茉莉酸甲酯生物合成及其调控机制[J].生物技术通报,2010(1):53-57.
LI Q Q,LI D P,LI D Q.The research progress in biosynthesis and regulation of jasmonates[J].BiotechnologyBulletin,2010(1):53-57.(in Chinese with English abstract)
[13] 曹建康,姜微波.果蔬采后生理生化实验指导[M].北京:中国轻工业出版社,2007.
[14] CAMPOS-VARGAS R,SALTVEIT M E.Involvement of putative chemical wound signals in the induction of phenolic metabolism in wounded lettuce[J].PhysiologiaPlantarum,2002,114(1):73-84.
[15] 何付新,远方,田宇,等.白菜伤害反应中内源信号分子及其基因表达变化[J].农业生物技术学报,2016,24(6):847-858.
HE F X,YUAN F,TIAN Y,et al.Expression of endogenous signal molecules and gene in wound response ofBrassicarapasubsp.pekinensis[J].JournalofAgriculturalBiotechnology,2016,24(6):847-858.(in Chinese with English abstract)
[16] 张亚宏,孙万仓,魏文慧,等.自交对甘蓝型油菜叶片SOD,CAT,APX活性的影响[J].华北农学报,2008,23(1):105-108.
ZHANG Y H,SUN W C,WEI W H,et al.Changes of SOD,CAT and APX ofBrassicanapusinfected by selfing in total growth period[J].ActaAgriculturaeBoreali-Sinica,2008,23(1):105-108.(in Chinese with English abstract)
[17] OMS-OLIU G,ODRIOZOLA-SERRANO I,SOLIVA-FORTUNY R,et al.The role of peroxidase on the antioxidant potential of fresh-cut ‘Piel de Sapo’ melon packaged under different modified atmospheres[J].FoodChemistry,2008,106(3):1085-1092.
[18] 何秀丽,张智乾,石立委,等.水杨苷异羟肟酸和茉莉酸甲酯对唐菖蒲试管结球的影响[J].园艺学报,2008,35(5):707-714.
HE X L,ZHANG Z Q,SHI L W,et al.Effects of salicylhydroxamic acid(SHAM) and methyl jasmonate(MJ) on invitrocorm formation ofGladiolushybridus[J].ActaHorticulturaeSinica,2008,35(5):707-714.(in Chinese with English abstract)
[19] 马玉荣,MIKAL SALTVEIT,王庆国.鲜切生菜伤害信号的初步研究[J].中国农学通报,2010,26(22):165-169.
MA Y R,SALTVEIT M,WANG Q G.Preliminary research on wound signal in fresh-cut lettuce[J].ChineseAgriculturalScienceBulletin,2010,26(22):165-169.(in Chinese with English abstract)
[20] ASEMOTA H N,WELLINGTON M A,ODUTUGA A A,et al.Effect of short-term storage on phenolic content,o-diphenolase and peroxidase activities of cut yam tubers(Dioscoreasp.)[J].JournaloftheScienceofFood&Agriculture,1992,60(3):309-312.
[21] 彭金英,黄勇平.植物防御反应的两种信号转导途径及其相互作用[J].植物生理与分子生物学学报,2005,31(4):347-353.
PENG J Y,HUANG Y P.The signaling pathways of plant defense response and their interaction[J].ActaPhotophysiologicaSinica,2005,31(4):347-353.(in Chinese with English abstract)
[22] 刘娜,马海军,刘静,等.水杨苷异羟肟酸对香水百合切花保鲜效应的研究[J].安徽农业科学,2012,40(22):11418.
LIU N,MA H J,LIU J,et al.Effects of salicylhydroxamic acid(SHAM) treatment on fresh-keeping effect of cutLiliumcasaBlanca flowers[J].JournalofAnhuiAgriculturalSciences,2012,40(22):11418.(in Chinese with English abstract)
[23] BI X F,WU J H,ZHANG Y,et al.High pressure carbon dioxide treatment for fresh-cut carrot slices[J].InnovativeFoodScience&EmergingTechnologies,2011,12(3):298-304.
[24] 王艳颖,胡文忠,姜波,等.茉莉酸甲酯处理抑制鲜切梨褐变机理的研究[J].食品工业科技,2012,33(23):339-342.
WANG Y Y,HU W Z,JIANG B,et al.Study on the mechanism of browning inhibited by methyl jasmonate treatment on fresh-cut pear[J].Science&TechnologyofFoodIndustry,2012,33(23):339-342.(in Chinese with English abstract)
[25] 张馨跃,王艳颖,李天,等.茉莉酸甲酯处理对鲜切芹菜贮藏品质的影响[J].保鲜与加工,2013,13(5):24-28.
ZHANG X Y,WANG Y Y,LI T,et al.Effects of methyl jasmonate treatment on storage quality of fresh-cut celery[J].Storage&Process,2013,13(5):24-28.(in Chinese with English abstract)
[26] 马杰,胡文忠,毕阳,等.外源乙烯处理对鲜切莴苣和甘蓝苯丙烷代谢的影响[J].食品工业科技,2013,34(21):327-329.
MA J,HU W Z,BI Y,et al.Effect of exogenous ethylene treatments onphenyloropanoid metabolism in tissues of fresh-cut lettuce and cabbage[J].Science&TechnologyofFoodIndustry,2013,34(21):327-329.(in Chinese with English abstract)
[27] 张梦如,龚明,杨玉梅,等.植物细胞质APX1的研究进展[J].中国农业科技导报,2015,17(3):8-18.
ZHANG M R,GONG M,YANG Y M,et al.Research progress on plant cytosolic APX1[J].JournalofAgriculturalScience&Technology,2015,17(3):8-18.(in Chinese with English abstract)
[28] 马杰,胡文忠,毕阳,等.外源乙烯和茉莉酸甲酯处理对鲜切莴苣活性氧代谢的影响[J].食品工业科技,2013,34(20):338-341.
MA J,HU W Z,BI Y,et al.Effect of exogenous ethylene and methyl jasmonate(MeJA) on reactive oxygen metabolism in tissue of fresh-cut cabbage[J].Science&TechnologyofFoodIndustry,2013,34(20):338-341.(in Chinese with English abstract)
Preliminaryresearchonwoundsignalinfresh-cutbambooshoots
HE Xiong1,XIAO Shangyue1,JIN Tunan1,LUO Haibo2,SUN Jincai1
(1.FacultyofFoodScience,ZhejiangPharmaceuticalCollege,Ningbo315100,China;2.GinlingCollege,NanjingNormalUniversity,Nanjing210097,China)
Abstract:In order to explore the wound signal induced by mechanical damage in fresh-cut bamboo shoots,the effect of fresh-cut treatment on the damage signal transmission of bamboo shoots at different distances from the cutting site and the influence of methyl jasmonate(MeJA) on the synthesis MeJA and transmission of signal molecule in fresh-cut bamboo shoots were investigated with the exogenous signal molecule and its synthetic inhibitor salicylhydroxamic acid(SHAM).The results showed that the wound signal substance was generated at the cleavage site,and it could be transmitted to the adjacent uninjured tissue to increase the activities of phenylalanine ammonia lyase(PAL) and peroxidase(POD).MeJA treatment could increase the activities of PAL,POD,superoxide dismutase(T-SOD) and ascorbate peroxidase(APX) in fresh-cut bamboo shoots,while SHAM treatment had a certain inhibitory effect.Thus,it was hypothesized that MeJA could induce and participate in the synthesis and transmission of some damage signal after mechanical damage in fresh-cut bamboo shoots.
Key words:plant physiology;methyl jasmonate;bamboo;signal transmission
中图分类号:Q945;S644.2
A
文章编号:1004-1524(2018)06-0992-07
收稿日期:2018-03-23
基金项目:浙江省教育厅科研项目(Y201432415);浙江省重点研发项目(2017C02021)
作者简介:何雄(1979—),女,浙江宁波人,硕士,副教授,研究方向为食品加工。E-mail:hex@mail.zjpc.net.cn
10.3969/j.issn.1004-1524.2018.06.15
(责任编辑高 峻)
