一个植物半胱氨酸蛋白酶多克隆抗体的制备及其应用
2018-06-28彭琪琪廖乾生张恒木
彭琪琪,羊 健,廖乾生,*,张恒木,*
(1.浙江理工大学 生命科学学院,浙江 杭州 310018;2.浙江省农业科学院 病毒与生物技术研究所,浙江 杭州 310021)
一个植物半胱氨酸蛋白酶多克隆抗体的制备及其应用
彭琪琪1,羊 健2,廖乾生1,*,张恒木2,*
(1.浙江理工大学 生命科学学院,浙江 杭州 310018;2.浙江省农业科学院 病毒与生物技术研究所,浙江 杭州 310021)
摘 要:半胱氨酸蛋白酶(cysteine proteinase,CysP)是一类重要的蛋白酶家族,广泛参与植物多种生理过程。为了分析植物CysP的特性,本实验首先通过RT-PCR技术从本氏烟中扩增获得了一个编码CysP的基因(NbCysP)序列并连接至pEASYTM-T5 Zero载体,测序验证后亚克隆至原核表达载体pGEX-6P1,命名为pGEX-NbCysP;将其导入大肠埃希菌BL21plysS中诱导表达;重组表达的NbCysP融合蛋白经过亲和层析纯化,免疫兔子制备多克隆抗体,Western印迹分析显示,该抗体能和重组NbCysP蛋白发生强烈的免疫学反应且条带单一,表明所获得的CysP抗体具有良好的特异性。上述结果为进一步鉴定植物CysP的功能特性奠定了基础。
关键词:半胱氨酸蛋白酶;原核表达;多克隆抗体
半胱氨酸蛋白酶(cysteine proteinase,CysP) 是一类维持生物体内蛋白代谢平衡的重要蛋白酶家族,广泛参与生物体的多种生理过程。近年来,研究人员相继从拟南芥[1]、烟草[2]、番茄[3]、水稻[4]、月季[5]和油菜[5]等植物中克隆到多种半胱氨酸蛋白酶基因。这些半胱氨酸蛋白酶大多属于papain(木瓜蛋白酶),其催化三联体主要由半胱氨酸(Cys)、组氨酸(His)、天冬氨酸或天冬酰胺(Asp/Asn)组成,它们在肽链中的排列顺序依次为Cys-His-Asn/Asp[6]。前人研究显示,半胱氨酸蛋白酶不仅参与液泡内蛋白的水解作用[7]、转录调控等过程[8],而且在植物与病原互作过程中发挥重要作用。Shindo等[9]发现拟南芥的一个CysP突变体对真菌Botrytiscinerea更为敏感;Kaschani等[10]利用VIGS技术沉默半胱氨酸蛋白酶C14基因,结果导致植株对卵菌Phytophthorainfestans更为感病。同样,西红柿半胱氨酸蛋白酶RCR3基因的突变不仅破坏了基于Cf-2介导的真菌抗性,其突变体植株甚至对卵菌P.infestans更为敏感[11-13]。另外,CysP也可以成为真菌、细菌、病毒等多种病原物的靶标,例如,番茄黄花叶曲叶病毒(Tomatoyellowleafcurlgeminivirus,TYLCV)的沉默抑制子V2与半胱氨酸蛋白酶CYP1互作并抑制其蛋白酶活性[14];细菌R.solanacearum蛋白PopP2与半胱氨酸蛋白酶RD19互作并使其定位于细胞核,从而干扰其介导的抗病活性[15]。
由于CysP在植物生长发育过程中的多重作用,受到研究人员越来越多的重视,但在模式植物——本氏烟的研究尚少。为了鉴定本氏烟CysP的功能特性,本实验从本氏烟中克隆了一个半胱氨酸蛋白酶家族蛋白基因(NbCysP),制备该蛋白的特异性多克隆抗体。
1 材料与方法
1.1 材料
本氏烟种子由本实验室保存并种植于人工气候温室;大肠埃希菌DH5α、BL21(DE3)plysS等感受态细胞和克隆载体pEASYTM-T5 Zero购自北京全式金生物技术有限公司;pGEX-6P1载体购自Novagen公司;DNA限制性内切酶购自NEB公司;ExTaq酶系列、dNTP Mix购自TaKaRa公司;T4 DNA 连接酶购自Thermo fisher scientific公司;PCR Mix购自南京诺唯赞生物科技有限公司;QIA quick Gel Extraction 试剂盒购自QIAGEN 公司;逆转录试剂盒购自Toyobo公司;羊抗兔IgG(过氧化物酶共价结合)购自Sigma公司;Novex ECL HRP Chemiluminescent Substrate Reagent Kit购自Invitrogen公司。
1.2 本氏烟总RNA和总蛋白的提取
收集新鲜的本氏烟叶片,利用TRIzol试剂,参照说明书提取本氏烟总RNA。按照羊健等[16]方法提取叶片总蛋白。提取的本氏烟叶片总RNA和总蛋白置于-80℃冰箱保存备用。
1.3 原核表达载体的构建
根据NCBI数据库中本氏烟NbCysP序列(登录号:XM_009771452)设计上游引物P-F(5′-CGGGATCCATGGCAACTCATAGCTCCACT-3′,下划线部分为BamHⅠ酶切位点)和下游引物P-R(5′-ACGCGTCGACGACTCGAACCTTCCCACTTAAG-3′,下划线部分为SalⅠ酶切位点)。以提取的本氏烟总RNA为模板,P-R为逆转录引物,参考Toyobo公司试剂盒说明书逆转录获得cDNA;取1~50 ng cDNA为模板,2μL上、下游引物(浓度为10μmol·L-1),再按照试剂盒说明调制PCR反应体系,94℃预变性5min;94℃变性30s,55℃退火30s,72℃延伸75s,35个循环。扩增产物经1%琼脂糖凝胶电泳并用QIA quick Gel Extraction试剂盒纯化后,连接至pEASYTM-T5 Zero载体,转化大肠埃希菌DH5α菌株,经菌落PCR和双酶切鉴定获得含目的基因的阳性克隆;目的基因再经BamHⅠ与SalⅠ双酶切和纯化后与相同双酶切的原核表达载体pGEX-6P1连接,转化大肠埃希菌DH5α,菌落PCR和测序鉴定后获得含有目的基因的表达载体pGEX-NbCysP。
1.4 重组NbCysP蛋白的诱导表达
将重组表达载体pGEX-NbCysP转化大肠埃希菌BL21(DE3)plysS菌株,挑取单菌落接种于含有50mg·mL-1氨苄的(Amp)LB培养基,于37℃培养过夜,第2天按1%的接种量转接到含Amp的新鲜LB培养基中,继续培养至D600值为0.5~0.8时加入IPTG,37℃继续振荡培养2~3h后,12000r·min-1离心1min收集,提取细菌总蛋白,按照羊健等[16]的方法分析重组蛋白的表达特点。
1.5 重组蛋白的纯化及抗体制备
参照GST.Bind Resin亲和层析试剂盒说明书,纯化重组的NbCysP蛋白。按照羊健等[16]方法用PBS缓冲液溶解纯化的蛋白,将纯化重组的NbCysP蛋白乳化后作为抗原免疫兔子,免疫3次后3~4d取血,4℃放置过夜,离心收集血清。
1.6 Western印迹分析
提取的本氏烟总蛋白经SDS-PAGE后,电转移至硝酸纤维膜上,以制备的NbCysP多克隆抗体为一抗,用HRP Chemiluminescent Substrate Reagent Kit 进行Western印迹分析,用Amersham imager 600(GE公司)成像系统获取图像信号。
2 结果与分析
2.1 原核表达载体pGEX-NbCysP的构建
我们首先以本氏烟总RNA为模板,利用引物对P-F/P-R进行PCR扩增,琼脂糖凝胶电泳显示NbCysP的特异性扩增片段大小约为1 400bp(图1),与预期大小一致。对其切胶纯化后,连接到T5克隆载体并转化大肠埃希菌DH5α,经菌落PCR和BamHⅠ/SalⅠ双酶切鉴定(图1),获得含有目的基因片段的重组克隆载体pT5-NbCysP。为了构建NbCysP蛋白的原核表达载体,从pT5-NbCysP质粒上切下目的片段,亚克隆至同样经双酶切的表达载体pGEX-6P1上,转化大肠埃希菌DH5α菌株,然后经双酶切(图1)鉴定,获得含有目的基因的重组原核表达载体质粒pGEX-NbCysP,测序结果也表明,该重组质粒中所含目的基因序列与数据库中序列完全一致,且具有完整的开放阅读框。
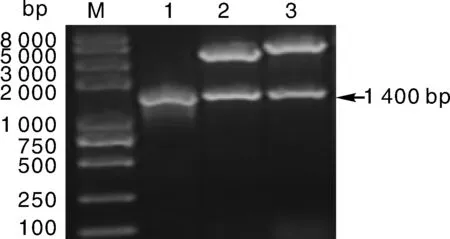
M,DL2000 DNA marker;1,NbCysP的PCR扩增产物;2,BamHⅠ和SalⅠ双酶切pT5-NbCysP;3,BamHⅠ和SalⅠ双酶切pGEX-NbCysP。M,DL2000 DNA marker;1,PCR amplification product of NbCysP;2,Double digestion of pT5-NbCysP by BamHⅠ and SalⅠ;3,Double digestion of pGEX-NbCysP by BamHⅠ and SalⅠ.图1 重组载体质粒pT5-NbCysP和pGEX-NbCysP的鉴定Fig.1 Identification of recombinant plasmids pT5-NbCysP and pGEX-NbCysP
2.2 NbCysP蛋白的表达与纯化
为了确定NbCysP重组蛋白是否表达,将诱导表达细菌样品的总蛋白用于SDS-PAGE分析。结果显示,与对照相比,含重组质粒pGEX-NbCysP的样品中存在一条高浓度的蛋白质条带,分子质量大小约77ku(图2);根据DNA man 6.0 程序计算,NbCysP相对分子质量预计约51ku,pGEX-6P1载体中GST标签大小约26ku,二者融合蛋白总相对分子质量与SDS-PAGE结果基本一致。为了表达大量的目的蛋白以利于纯化,又对诱导条件进行了优化,发现IPTG终浓度为1.0mmol·L-1,37℃诱导3h的条件下表达效果较好,之后的研究均采用同样的条件。为了获得纯化的蛋白,我们利用GST Bind Resin进行亲和层析纯化了重组的NbCysP蛋白。纯化的蛋白经SDS-PAGE检测为单一条带且大小与上述结果一致(图2-B),表明获得纯化的重组NbCysP蛋白。
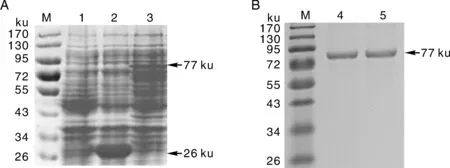
M,蛋白marker;1,诱导后的BL21 playS;2,诱导后的仅含pGEX-6P1空载体的宿主菌;3,诱导后含pGEX-NbCysP的宿主菌;4~5,纯化的NbCysP蛋白。M,Protein marker;1,The induced BL21 playS;2,The induced host bacteria with pGEX empty vector;3,The induced host bacteria with pGEX-NbCysP;4-5,purified NbCysP protein.图2 SDS-PAGE分析重组NbCysP蛋白Fig.2 Analysis of recombinant NbCysP protein by SDS-PAGE
2.3 NbCysP蛋白多克隆抗体的特异性分析
为了分析NbCysP多克隆抗体的特异性,将纯化的抗体按1∶5000的比例稀释并用于原核表达蛋白的检测,Western印迹结果表明,在含pGEX-NbCysP的大肠埃希菌裂解物样品中检测到一条强烈的信号条带,且该条带特异性地位于相对分子质量约77ku(图3),而在不含重组载体质粒的大肠埃希菌裂解物样品中则检测不到反应信号(图3),这些结果表明,本研究所制备的多克隆抗体能够与目标蛋白发生免疫学反应且条带单一,所获得的NbCysP多克隆抗体具有良好的特异性。
2.4 NbCysP蛋白多克隆抗体的应用
为了进一步应用该抗血清,我们又提取了本氏烟植株的总蛋白,利用该多克隆抗体进行Western 印迹(图4)。结果发现该多克隆抗体可与本氏烟样品发生强烈的血清学反应,且反应条带特异性地位于相对分子质量约51ku处,表明该抗体可特异性地与本氏烟内源CysP蛋白反应,支持该抗体具有良好特异性的结论。
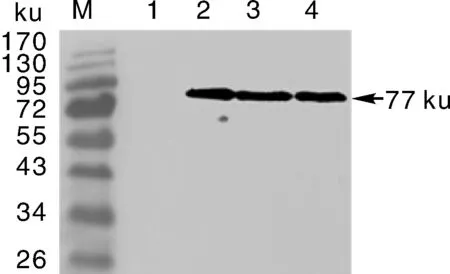
M,蛋白marker;1,不含质粒的大肠埃希菌菌株;2~4,诱导表达的含pGEX-6P1-NbCysP的菌株。M,Protein marker;1,The induced BL21 playS;2-4,The induced host bacteria with pGEX-NbCysP.图3 Western印迹分析多克隆抗体的特异性Fig.3 Specificity of polyclonal antibody by Western blotting assays
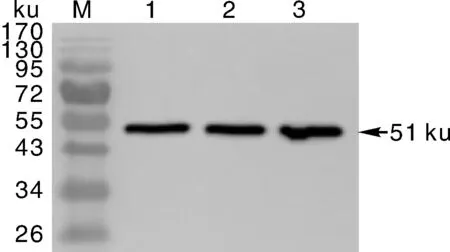
M,蛋白marker;1~3,本氏烟植物样品的生物学重复。M,Protein marker;1-3,Biological replicates of N.benthamiana samples.图4 多克隆抗体在本氏烟內源NbCysP检测中的应用Fig.4 Application of multiclonal antibodies for detection of endogenous NbCysP in N.benthamiana
3 讨论
半胱氨酸蛋白酶广泛参与植物的各种生理活动,是植物体内重要的调节因子。本研究中克隆的NbCysP与拟南芥的Cathepsin B、RD21A、本氏烟Cathepsin B,番茄C14等都属于半胱氨酸蛋白酶家族且具有一定的氨基酸序列同源性[9,17]。研究表明拟南芥和本氏烟Cathepsin B均可参与细胞的程序性死亡(PCD)过程[18-19],是植物超敏反应过程中一个关键因子;如Cathepsin B基因突变的拟南芥植株PCD过程受到抑制。拟南芥RD21A蛋白除了参与免疫反应之外,还在衰老的叶片组织中大量积累,表明拟南芥RD21A蛋白还在植物衰老过程中起作用[9,20]。番茄C14蛋白酶与P.infestansAvrBlb2互作,阻止C14分泌进入质体从而抑制其介导的抗病反应[21],表明番茄C14半胱氨酸蛋白酶在植物抗病过程中起重要作用。但本研究克隆的NbCysP功能研究尚未见报道。本研究中,我们还制备了NbCysP的多克隆抗体,Western印迹分析显示该抗体具有良好的特异性,这为深入分析NbCysP的功能奠定了基础。
:
[1] ZHANG X,LIU S,TAKANO T.Two cysteine proteinase inhibitors fromArabidopsisthaliana,AtCYSa,and AtCYSb,increasing the salt,drought,oxidation and cold tolerance[J].PlantMolecularBiology,2008,68(1/2):131-143.
[2] ZHANG X M,WANG Y,LV X M,et al.NtCP56,a new cysteine protease inNicotianatabacumL.involved in pollen grain development[J].JournalofExperimentalBotany,2009,60(6):1569-1577.
[3] CHEN G H,HUANG L T,YAP M N,et al.Molecular characterization of a senescence-associated gene encoding cysteine proteinase and its gene expression during leaf senescence in sweet potato[J].Plant&CellPhysiology,2002,43(9):984-991.
[4] ABE K,EMORI Y,KONDO H,et al.Molecular cloning of a cysteine proteinase inhibitor of rice(oryzacystatin).Homology with animal cystatins and transient expression in the ripening process of rice seeds[J].JournalofBiologicalChemistry,1987,262(35):16793-16797.
[5] TRIPATHI S K,SINGH A P,SANE A P,et al.Transcriptional activation of a 37 kDa ethylene responsive cysteine protease gene,RbCP1,is associated with protein degradation during petal abscission in rose[J].JournalofExperimentalBotany,2009,60(7):2035-2044.
[6] WIEDERABDERS B.Structure-function relationships in class CA1 cysteine peptidase propeptides[J].ActaBiochimicaPolonica,2003,50(3):691-713.
[7] AVILA E L,ZOUHAR J,CARTER D G,et al.Tools to study plant organelle biogenesis.point mutation lines with disrupted vacuoles and high-speed confocal screening of green fluorescent protein-tagged organelles[J].PlantPhysiology,2003,133(4):1673-1676.
[8] MATARASSO N,SCHUSTER S,AVNI A.A novel plant cysteine protease has a dual function as a regulator of 1-aminocyclopropane-1-carboxylic acid synthase gene expression[J].PlantCell,2005,17(4):1205-1216.
[9] SHINDO T,MISASVILLAMIL J C,HORGER A C,et al.A role in immunity forArabidopsiscysteine protease RD21,the ortholog of the tomato immune protease C14[J].PLoSOne,2012,7(1):e29317.
[10] KASCHANI F,SHABAB M,BOKURT T,et al.An effector-targeted protease contributes to defense against phytophthora infestans and is under diversifying selection in natural hosts[J].PlantPhysiology,2011,154(1):1794-1804.
[11] LOZANOTORRES JL,WILBERS RH,GAWRONSKI P,et al.Dual disease resistance mediated by the immune receptor Cf-2 in tomato requires a common virulence target of a fungus and a nematode[J].ProceedingsoftheNationalAcademyofSciencesoftheUnitedStatesofAmerica,2012,109(25):10119-10135.
[12] SONG J,WIN J,TIAN M Y,et al.Apoplastic effectors secreted by two unrelated eukaryotic plant pathogens target the tomato defense protease Rcr3[J].ProceedingsoftheNationalAcademyofSciencesoftheUnitedStatesofAmerica,2009,106(5):1654-1659.
[13] ILYAS M,HORGER A,BOZKURT T,et al.Functional divergence of two secreted immune proteases of tomato[J].CurrentBiology,2015,25(17):2300-2306.
[14] BARZIV A,LEVY Y,CITOVSKY V,et al.TheTomatoyellowleafcurlvirus(TYLCV) V2 protein inhibits enzymatic activity of the host papain-like cysteine protease CYP1[J].Biochemical&BiophysicalResearchCommunications,2015,460(3):525-529.
[15] BERNOUX M,TIMMERS T A,BRIERE C,et al.RD19,anArabidopsiscysteine protease required for RRS1-R-Mediated resistance,is relocalized to the nucleus by theRalstoniasolanacearumPopP2 effector[J].PlantCell,2008,20(8):2252-2264.
[16] 羊健,张恒木,陈剑平,等.水稻黑条矮缩病毒p8蛋白的原核表达、抗血清制备及其特性[J].植物保护学报,2007,34(3):252-256.
YANG J,ZHANG,H M,CHEN J P,et al.Prekaryotic expression antiserum preparation and some properties of p8 protein ofRiceblack-streakeddwarfFijivirus[J].ActaPhytophylacicaSinica,2007,34(3):252-256.(in Chinese with English abstract)
[17] RICHAU K H,KASCHANI F,VERDOES M,et al.Subclassification and biochemical analysis of plant papain-like cysteine proteases displays subfamily-specific characteristics[J].PlantPhysiology,2012,158(4):1583-1599.
[18] GE Y,CAI Y M,BONNEAU L,et al.Inhibition of cathepsin B by caspase-3 inhibitors blocks programmed cell death inArabidopsis[J].CellDeath&Differentiation,2016,23(9):1493-1501.
[19] GILORY E M,HEIN I,HOOM R V D,et al.Involvement of cathepsin B in the plant disease resistance hypersensitive response[J].PlantJournal,2007,52(1):1-13.
[20] KOIZUMI M,YAMAGUCHIi-SHINOZAKI K,TSUJI H,et al.Structure and expression of two genes that encode distinct drought-inducible cysteine proteinases inArabidopsisthaliana[J].Gene,1993,129(2):175-182.
[21] BOZKURT TO,SCHORNACK S,WIN J,et al.Phytophthora infestans effector AVRblb2 prevents secretion of a plant immune protease at the haustorial interface[J].ProceedingsoftheNationalAcademyofSciencesoftheUnitedStatesofAmerica,2011,108(51):20832-20837.
Preparationandapplicationofmulticlonalantibodyagainstplantcysteineproteinase
PENG Qiqi1,YANG Jian2,LIAO Qiansheng1,*,ZHANG Hengmu2,*
(1.CollegeofLifeSciences,ZhejiangSci-TechUniversity,Hangzhou310018,China;2.InstituteofVirologyandBiotechnology,ZhejiangAcademyofAgriculturalSciences,Hangzhou310021,China)
Abstract:Cysteine proteases(CysPs) are a family of important proteases that are involved in a wide range of plant physiological processes.To characterize such proteinase,a gene encoding CysP was amplified by RT-PCR fromNicotianabenthamianaplants and ligated into pEASYTM-T5 Zero vector.The insertion was sequenced and then sub-cloned into the prokaryotic expression plasmid pGEX-6P1,named as pGEX-NbCysP.The plasmid was transformed intoEscherichiacoliBL21plysS for inducible expression.The recombinant NbCysP fusion protein was purified with affinity chromatography and used for producing polyclonal antibody by immunizing rabbits.In immunoblotting assays,the polyclonal antibody could react strongly with the recombinant NbCysP protein as the presence of a single band,indicating that the antibody was specific against the proteinase.These results laid a foundation for further characterization of its function.
Key words:cysteine proteinase;prokaryotic expression;multiclonal antibodies
中图分类号:Q78
A
文章编号:1004-1524(2018)06-0881-05
收稿日期:2018-03-28
基金项目:国家自然科学基金(3150160)
作者简介:彭琪琪(1992—),男,河南驻马店人,硕士研究生,主要从事植物病理学研究。E-mail:1399767318@qq.com
,廖乾生,E-mail:qshliao@aliyun.com;张恒木,E-mail:zhhengmu@tsinghua.org.cn
10.3969/j.issn.1004-1524.2018.06.01
(责任编辑张 韵)
