不同生长期山核桃林碳氮磷生态化学计量特征
2018-05-24李晨茜闫道良郑炳松
李晨茜 闫道良 郑炳松
(浙江农林大学林业与生物技术学院 临安 311300)
生态化学计量学相关理论表明,植物中的C(碳)、N(氮)、P(磷)元素与土壤养分的供给、根系的吸收与生长、林冠光合生产等过程存在着复杂的耦合作用[1]。这些元素间的耦合过程使土壤—植物体—凋落物养分元素含量具有明显的时空变化。N∶P(氮磷比)不仅是决定群落结构和功能的关键性指标,也可以作为对群落生产力起限制性作用的指示指标[2],C∶N(碳氮比)和C∶P(碳磷比)描述了植物体结构性物质C与养分的比值关系,也即养分利用效率。研究表明,植物在年生长周期以及个体发育的不同阶段,对养分的需求存在着差异[3-4]。近年来,生态化学计量学的发展和完善为植物养分季节变化方面提供了新的思路和研究手段。因此,通过了解植物在不同发育阶段中的元素计量关系,有助于了解植物生长中主要的限制因子,尤其是对群落结构简单,植被单一脆弱的人工山核桃林生态系统演化的正向维持,及其管理措施的制定有着重要的指导意义。
植物体内的C、N、P含量及其比例关系是与环境作用的综合结果[5-6]。对植物而言,元素的失衡会影响植物的生长代谢过程,导致植物的严重病虫害,影响经济林的有效产出。鉴于此,本研究以人工山核桃林为研究对象,采用生态化学计量学的原理来研究山核桃林地土壤、细根、叶的C、N、P生态化学计量学季节变化特征,以期为山核桃农户分阶段科学管理山核桃林提供理论技术支撑。
1 材料与方法
1.1 采样地概况
采样地位于浙江省临安板桥镇(北纬30°,东经119°),山核桃栽培地山坡西南向,海拔103.8 m,属中纬度北亚热带季风气候,年均降雨量为1 613.9 mm,其中降雨多集中于4~10月,年均气温为15.8 ℃。无霜期年均237 d,采样地土壤是由石灰质岩发育而成,土壤含量少,多是石块。采样地坡度30~35°。
1.2 采样方法
选择样线上(同一海拔高度)长势一致的45 a龄丰产山核桃9株,随机取3株作为一个重复,摘取每株树体中部向阳生长的复叶10枚,采样时间为2015年每月15日(从5月开始至10月,其中10月采样是在月末,叶已经凋落)每次共采摘90枚复叶,于80 ℃下烘干至恒重,粉碎过筛备用。选取树干周围1.5 m处,刮去地表异物,采挖0~20 cm深的土壤,去除石子、枯根等杂物,于室内自然风干后,碾磨过100目筛备用。山核桃细根取样(直径<2 mm)在采土样时同时进行。
1.3 土壤指标和植株全N全P含量测定
每次采集的样本带回实验室及时处理并测定分析,土壤有机质,(以C含量为衡量指标)和全N测定分别采用重铬酸钾容量法—外加热法(GB 9834-88)和半微量开氏法(GB 7173-87),酸溶法测定土壤全P。根、叶有机C采用重铬酸钾容量法—外加热法,全N、全P含量测定采用硫酸-过氧化氢消化法[7]。
1.4 数据处理及分析
C、N、P含量采用质量分数g·kg-1表示。植物N、P养分重吸收率的计算方法如下:N (P)重吸收率(%) = (植物生长挂果期活体叶片N (P) 含量-凋落物N (P)含量)/植物生长挂果期活体叶片N (P)含量× 100,其中山核桃生长挂果期采样时间是8月15日,凋落物采样时间是10月底。采用Aerts and Chapin[8]和Ellison[9]的研究结果用以诊断营养元素限制关系,即当植株的叶:N>20 mg·g-1,P>1 mg·g-1,植物生长既不受N也不受P限制;N>20 mg·g-1,P<1 mg·g-1,植物生长受P限制;N<20 mg·g-1,P>1 mg·g-1,植物生长受N限制。
数据采用SPSS 13.0软件进行统计分析,单因素方差分析判断处理间各指标的差异,显著性水平为p<0.05或p<0.01。
2 结果与分析
2.1 山核桃林地土壤C、N、P含量变化及其化学计量比
从5月到10月,土壤有机质(C)含量变化幅度为28.73~42.00 g·kg-1,最高月份(7月)有机质含量是最低月份(8月)的1.46倍,有较大的变化。土壤N含量为1.75~0.58 g·kg-1,最高位7月份,是最低月份10月的3.02倍,P含量为0.45~1.31g·kg-1,最高为6月份,是最低月份10月的2.91倍,变化显著。从表1还可以看出,土壤有机质C含量变化表现“单峰”趋势,N和P也呈现同样的变化趋势。
土壤的C∶N比变化幅度为24.00~61.30,最高在10月,是最低月份(7月)的2.55倍;C∶P比的变化幅度为27.34~78.39,最高和最低月份分别为10月及6月,最高月份为最低月份的2.87倍,有较大的差异,变化趋势呈现“单峰”模式;N:P比变化幅度为0.80~1.42,最高月份(7月)是最低月份(9月)的1.78倍,变化趋势呈现“双峰”模式。
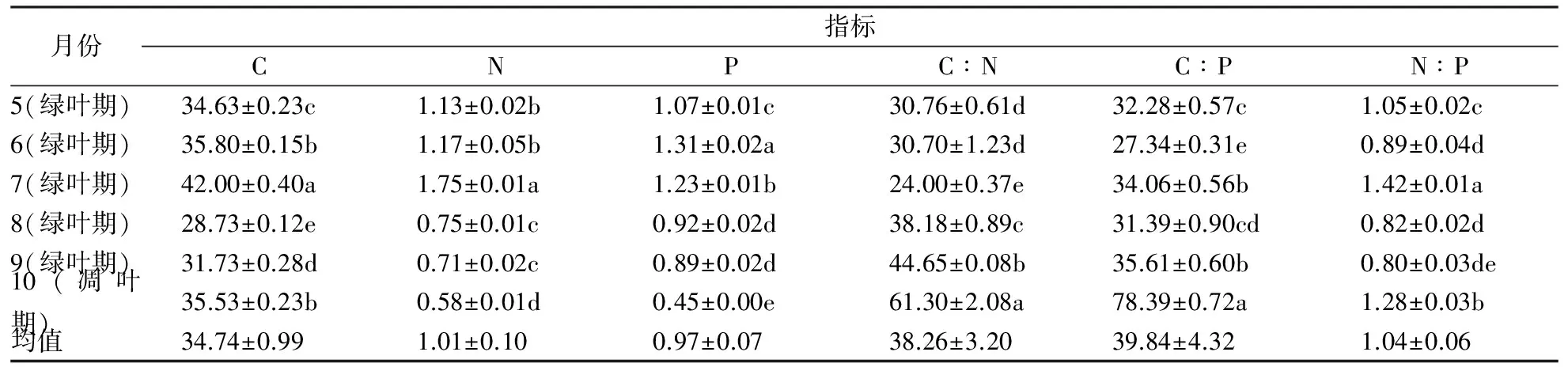
表1 土壤C、N、P含量及其化学计量随季节的变化
注:同一列数据后的不同字母表示差异显著性(p<0.05)
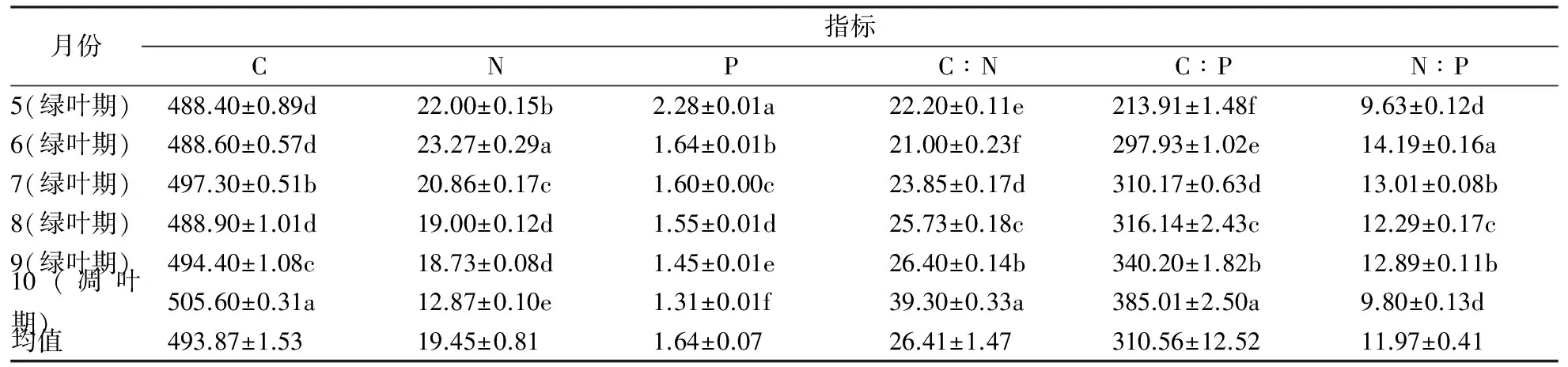
表2 叶C、N、P含量及其计量比随季节的变化
注:同一列数据后的不同字母表示差异显著性(p<0.05)
2.2 山核桃叶片C、N、P含量变化及其化学计量比
不同月份的绿叶叶片C含量在488.40~497.30 g·kg-1,其中在果实生长发育初期(7月)含量最高,为497.30 g·kg-1,以后随着果实的膨大与成熟,叶片C含量呈先下降后上升趋势,但变化不显著。绿叶的N含量最高值在叶片生长发育中期(6月)为23.27 g·kg-1,随着植物的生长,叶片N的含量逐渐下降,最低月份(9月)较最高月份下降了1.24倍。绿叶P的含量最高值出现在叶片生长初期(5月)为2.28 g·kg-1,之后叶片P含量逐月显著下降,最低月份较最高月份下降了1.57倍。对叶凋落物而言,其C含量与绿叶相比,为最高505.6 g·kg-1,显著有别于绿叶,而凋落物中N含量和P含量均降低到最低值,分别为12.87 g·kg-1和1.31 g·kg-1,与落叶前有显著差别。叶片的C、N、P含量变化均呈现“单峰”模式。
绿叶叶片C、N、P间比值变异在不同月份均表现有不同的差异。在5月,叶低C含量和高P含量导致C∶P明显低于其它月份的计量比,而N∶P也表现最低(9.63)。在6月,叶低C含量和高N含量导致C∶N最低(21.00)而N∶P表现最高 (14.19)。在9月,叶高C含量和低N、P含量导致C∶N(26.40)及C∶P(340.20)最高。叶凋落物由于高C含量和低N、P含量导致C∶N(385.01)和C∶P(39.30)最高,而N∶P在凋落物中表现最低(9.80)。
2.3 山核桃细根C、N、P含量变化及其化学计量比
细根C含量在山核桃生长季变化幅度较小 (498.80~507.87 g·kg-1),随着叶逐渐凋落,细根C含量有增高的趋势 (9~10月),在10月叶完全凋落后,达到最高 (507.87 g·kg-1)。细根N含量在山核桃生长中期(7月)和末期 (10月)显著高于其它生长月份的N含量,同时,P含量在统计学上亦表现最高。在8月N含量最低,为9.34 g·kg-1,P含量最低出现在6月,为1.09 g·kg-1(表3)。
随着山核桃叶的逐渐展开到发育成熟,细根C∶N比表现明显的升高,在8月达到最高,为53.72,而后随着果实的渐熟,细根C∶N呈现显著下降趋势。由于细根P含量在7月最高,引起C∶P和N∶P比在该月最低,分别为431.62和8.11。
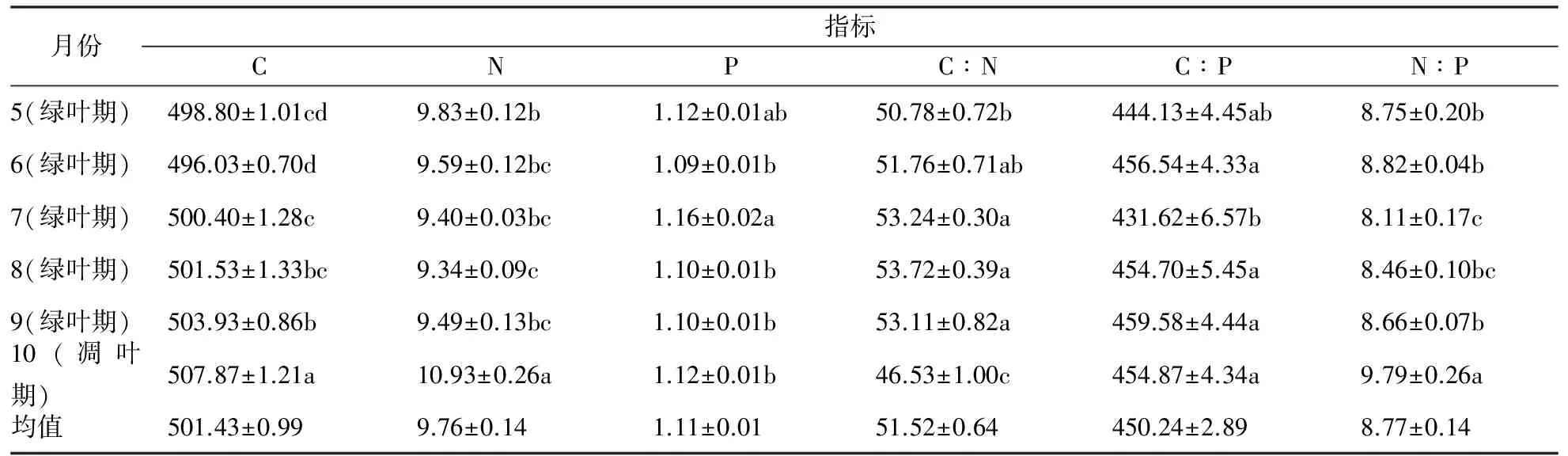
表3 细根C、N、P含量及其计量比随季节的变化
注:同一列数据后的不同字母表示差异显著性(p<0.05)
2.4 C、N、P化学计量比关联分析
叶C物质的积累量与叶N和叶P含量极显著负相关,却与叶C∶N、C∶P极显著正相关。叶N含量与叶P、N∶P,根C∶N,土壤N、P分别正相关。叶P含量与叶中的C∶N、C∶P和根中的C分别显著负相关(见表4)。
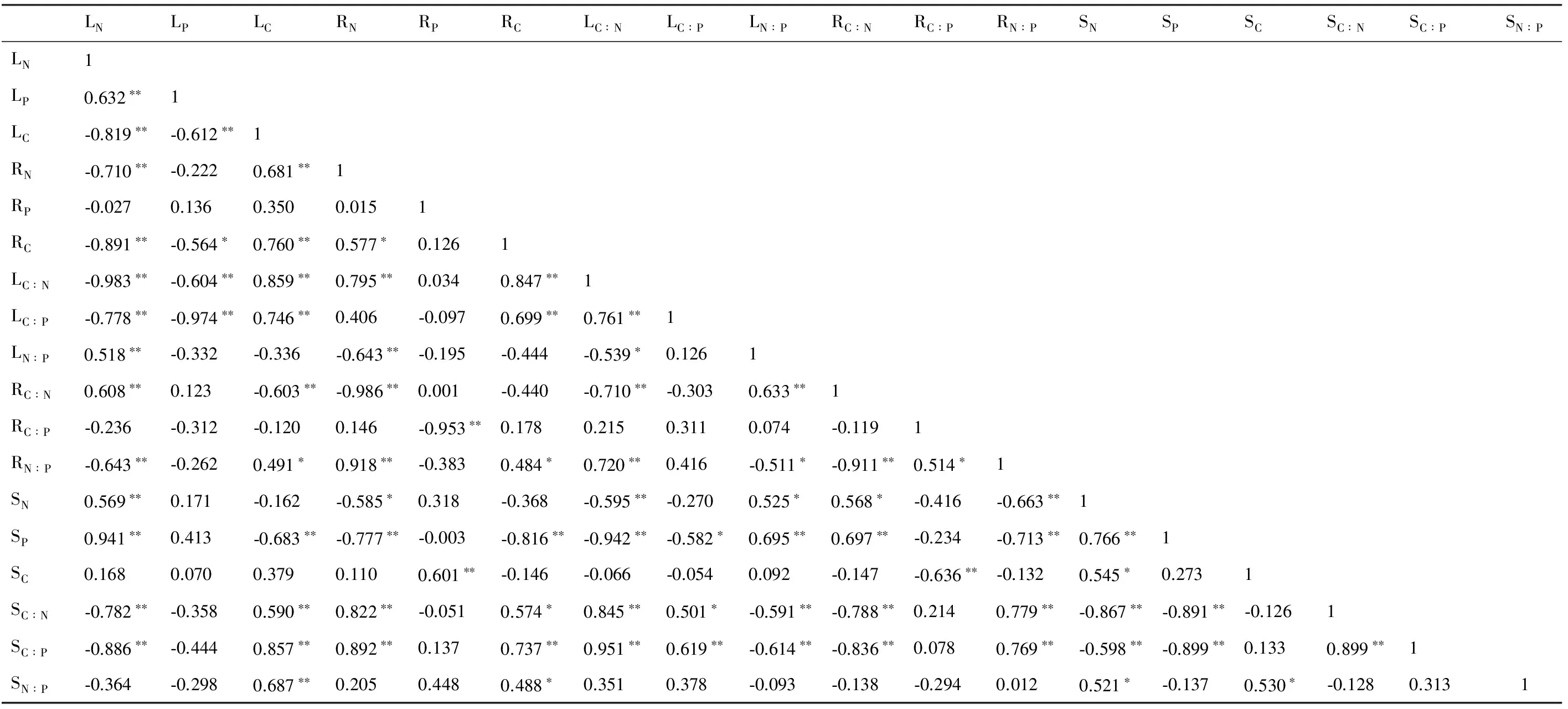
表4 C、N、P化学计量比相关分析
注:LN, leaf N; LP, leaf P; LC, leaf C; RN, root N; RP, root P; RC, root C; LC∶N, leaf C∶N; LC∶P, leaf C∶P; LN∶P, leaf N∶P; RC∶N, root C∶N; RC∶P, root C∶P; RN∶P, root N∶P; SN,soil N; SP, soil P; SC, soil C; SC∶N, soil C∶N; SC∶P, soilC∶P; SN∶P, soil N∶P
根中的C物质积累却与N含量表现正相关, 然而根中的N含量却与土壤中的N、P分别显著负相关,根中的P却与土壤C显著正相关。
2.5 山核桃叶N、P养分重吸收
山核桃叶凋落物N、P平均含量分别为12.87、1.31 g·kg-1。山核桃叶N的重吸收率为32.26%,P重吸收率为15.48%,N重吸收率显著高于P重吸收率(P<0.05)。
3 讨论
3.1 土壤C、N、P化学计量关系变异特点及其关联性
土壤中的N和P,是植物赖以生存的主要物质基础,也是限制植物生长的重要养分,而N∶P是可用作确定养分限制阈值的一个指标[10]。人工山核桃林土壤N∶P均值为1.14,低于伊犁河谷野果林土壤N∶P(6.05)[11],远低于中国土壤平均值(9.3)和全球平均值(13.1)[12-13]。研究表明,温带地区植物生长主要受土壤N限制,如黄土丘陵土壤的N∶P为0.86,表现为显著N缺乏[14]。人工山核桃林土壤有如此低的N∶P表明,山核桃林土壤缺P可能性很小,土壤N可能相对缺乏。土壤养分元素之间是密切相关的。本研究中,土壤N含量与土壤P含量极显著正相关,与土壤C显著正相关。土壤P含量与土壤C含量也表现一定的正相关,这与已有研究土壤P含量的变化趋势随有机质含量的增加而增加相一致[15]。总体上,山核桃林地土壤C、N、P三者之间均为相互促进、协同增长的关系。土壤C∶P低有利于促进微生物分解有机质释放养分,促进土壤中有效P含量的增加,高的C∶P不利于植物的生长[16]。山核桃林土壤C∶P值为38.87,低于中国土壤均值(136),远低于全球均值(186)[12-13],表明山核桃林土壤P含量较高。同时,在一定的全氮含量范围内,土壤P增加的速率高于N含量增加的速率(拟合方程为:y=-0.18+1.42x,y为P含量,x为N含量)。原因可能在于N能促进植物生长,尤其促进根系生长,而植物在生长过程中根系会分泌一些低分子有机酸等物质,对土壤中的P具有一定的活化作用,而且有机N肥本身的生理酸性也促进了磷酸盐的溶解和释放,增加了土壤中的P含量。因此,增加山核桃林地土壤有机质的含量,对于山地P含量的提升有促进作用。土壤中的有机C、N和P在不同月份一般都表现显著变化,这可能与植物不同的生长发育阶段对元素的需量与吸收有关。
3.2 山核桃C、N、P化学计量关系变异特征及N、P重吸收
按照Aerts and Chapin 和Ellison的标准,山核桃在57月三个月的生长期间,N>20 g·kg-1,P>1 g·kg-1,植株生长既不受N限制,也不受P限制。但在山核桃果实膨大关键期(不饱和脂肪酸合成期),即8、9月两个月中,N<20 g·kg-1,P>1 g·kg-1,表明植株生长受到了N限制,这与笔者观察测定的该期间山核桃叶片叶绿素含量明显小于其它生长期间有一致之处(因为叶绿素含量与叶N含量成显著正相关,数据没有给出)。以上说明,在山核桃果实不饱和脂肪酸形成关键期增施适量的N肥是提高山核桃品质的重要因素之一(数据没有给出)。叶C与叶片的C∶N和C∶P呈现极显著相关,由此可以推断N、P的协调供应对山核桃植株碳水化合物合成的重要性。植物叶片的C∶N和C∶P反映植物吸收营养所能同化C的能力,在一定程度上体现植物的营养利用效率。山核桃叶片C∶N和C∶P分别为26.41和310.56,高于全球植物平均水平22.5和232,说明山核桃对营养利用效率较高。
养分重吸收是植物一种重要的养分保护策略。研究表明,莎草科植物无脉薹草(Carexenervis) N的重吸收率达68%,该科植物中P的重吸收利用率高达80%[17],长芒草(Stipabungeana)的N和P重吸收率分别为60.35%和71.75%[18]。新银合欢(Leucaenaleucocephala) 的N、P重吸收率分别在42.76%~55.90%和26.35%~40.60%之间,N重吸收率均大于P重吸收率,而全球水平的N和P重吸收率约为50%[3]。比较而言,山核桃的N、P重吸收率明显较低,这除了与不同植物的遗传因素决定养分的重吸收有差异之外,还可能与植物生长的环境因子有关。已有研究表明,在养分贫瘠的环境,植物通过加强养分重吸收适应生存环境[19]。但另有研究认为,较高的N、P养分转移率并不是适应贫瘠环境的机制,可能是物种固有的特征[20]。研究表明,山核桃没有通过加强自身的N重吸收提高对缺N的限制,关于山核桃养分重吸收的机制是我们下一步需要迫切弄清的问题之一,相信问题的进一步阐明,对于山核桃林N循环规律的了解,以及科学经营山核桃林提供指导。
参 考 文 献
[1]Ladanai S,Agren G I, Olsson B A. Relationships between tree and soil properties inPiceaabiesandPinussylvestrisforests in Sweden[J]. Ecosystems, 2010, 13(2): 302-316.
[2]Güsewell S. N: P ratios in terrestrial plants: Variation and functional significance[J]. New Phytologist, 2004, 164: 243-266.
[3]邓浩俊, 陈爱民, 严思维, 等. 不同林龄新银合欢重吸收率及其C:N:P化学计量特征[J]. 应用与环境生物学报,2015,21(3):522-527.
[4] 闫道良, 黄有军, 金水虎, 等. 山核桃功能器官细根、叶和林地土壤C、N、P化学计量时间变异特征[J]. 水土保持学报, 2013, 27(5):255-259.
[5] Elser J J, Sterner R W, Gorokhova E, et al. Biological stoichiometry from genes to ecosystems[J]. Ecology Letters, 2000, 3(6): 540-550.
[6] Zeng D H, Chen G S. Ecological stoichiometry: a science to explore the complexity of living systems[J]. Chinese Journal of Plant Ecology, 2005, 29(6): 1007-1019.
[7]张韫. 土壤·水·植物理化分析教程[M]. 北京: 中国林业出版社, 2011.
[8]Aerts R, Chapin F S. The mineral nutrition of wild plants revisited: a re-evaluation of processes and patterns[J]. Advances in Ecology Research, 2000, 30:1-67.
[9]Ellison A M. Nutrient limitation and stoichiometry of carnivorous plants[J]. Plant Biology, 2006, 8,:740-747.
[10]洪江涛, 吴建波, 王小丹. 全球气候变化对陆地植物碳氮磷生态化学计量学特征的影响[J]. 应用生态学报, 2013(24): 2658-2660.
[11]陶冶, 张元明, 周晓兵. 伊犁野果林浅层土壤养分生态化学计量特征及其影响因素[J]. 应用生态学报, 2016, 27(7): 2239-2248.
[12] Zhao F, Sun J, Ren C, et al. Land use change influences soil C, N, and P stoichiometry under Grain-to-Green Program in China[J]. Scientific Reports, 2015, 5: 10195.
[13] Cleveland C C, Liptzin D. C:N:P stoichiometry in soil: Is there a “Redfield ratio” for the microbial biomass? [J]. Biogeochemistry, 2007, 85: 235-252.
[14]朱秋莲, 邢肖毅, 张宏, 等. 黄土丘陵沟壑区不同植被区土壤生态化学计量特征[J]. 生态学报, 2013,33(15): 4674-4682.
[15]Liang G Q, Lin B, Lin J X, et al. Effect of long term fertilization on the forms of inorganic phosphorus in calcareous fluvo-aquic soil[J]. Plant Nutrition and Fertilizer Science, 2001, 7(3): 241-248.
[16] 王建林, 钟志明, 王忠红, 等. 青藏高原高寒草原生态系统土壤碳磷比的分布特征[J]. 草业学报, 2014, 23(2): 9-19.
[17] Niklas K J. Plant allometry, leaf nitrogen and phosphorus stoichiometry, and interspecific trends in annual growth rates[J]. Annuals of Botany, 2006, 97: 155-163.
[18] 安卓, 牛得草, 文海燕, 等. 氮素添加对黄土高原典型草原长芒草氮磷重吸收率及C:N:P化学计量特征的影响[J]. 植物生态学报, 2011, 35(8): 801-807.
[19] 齐泽民, 王开运. 密度对缺苞箭竹凋落物养分归还及养分利用效率的影响[J]. 应用生态学报, 2007, 18:2025-2029.
[20]阎恩荣, 王希华, 郭明, 等. 浙江天童常绿阔叶林、常绿针叶林与落叶阔叶林的C:N:P化学计量特征[J].植物生态学报, 2010, 34 (1): 48-57.
